
A. M. Ahmed
Department of Zoology, Faculty of Sciences, Minia University, El-Minia, Egypt







ABSTRACT
Great efforts are currently being done to utilize the immune system of mosquito vector in the battle against malaria. However, this strategy still facing some limiting factors mainly the reproductive cost, the price of immune induction. This study has been introduced to show that inducing non-costly immune responses in the African malaria vector, Anopheles gambiae, is possible via the oral administration of Black Seed Oil (BSO), a natural botanical extract from Nigella sativa, to mosquitoes. The humoral anti-Micrococcus luteus activity, induced by LPS injection and melanization, against inoculated Sephadex® beads, were studied here (at 18 h post-treatment). The humoral anti-bacterial activity was measured using inhibition zone assay. The degrees of melanization response against negatively charged CM C-25 and neutral CM G-25 Sephadex® beads were monitored under the microscope. Reproductive cost was monitored in terms of percentages of follicular apoptosis and resorption at 18 and 24 h post-treatment, respectively. Data of this study demonstrate that, mosquitoes maintained on 0.3% BSO-glucose mixture (in 10% glucose) showed, on one hand, significant increases in the humoral anti-bacterial activity or melanization response when injected with LPS or inoculated with beads respectively. Moreover, these tow responses were more pronounced in blood fed mosquitoes. On the other hand, significant declines in the % of follicular apoptosis and resorption were reported in the ovaries of the same immunized mosquitoes. In addition, BSO showed no toxic effect on ookinetes development, both in vivo and in vitro, or on the viability of mosquito cells in vitro. Therefore, these data may indicate that the non-toxic BSO may have a dual effect as it enhanced immunity and reduced the concomitant impact on the reproductive fitness. Thus, this study suggests BSO as a candidate in support of the immuno-control strategy in the battle against malaria.
PDF Abstract XML References Citation
How to cite this article
A. M. Ahmed, 2007. A Dual Effect for the Black Seed Oil on the Malaria Vector Anopheles gambiae: Enhances Immunity and Reduces the Concomitant Reproductive Cost. Journal of Entomology, 4: 1-19.
DOI: 10.3923/je.2007.1.19
URL: https://scialert.net/abstract/?doi=je.2007.1.19
DOI: 10.3923/je.2007.1.19
URL: https://scialert.net/abstract/?doi=je.2007.1.19
INTRODUCTION
Malaria is one of the most serious and deadly diseases as it causes up to 300 million clinical cases and kills over 2 millions a year, most of them are from children under 5 (WHO, 1999). Yet, the situation can be expected to get worse, not only because of the increasing resistance of the parasites and their vectors against drugs and insecticides, respectively (WHO, 1999), but also because of the deteriorating socioeconomical conditions in many malaria-endemic areas in the world (Manfredi, 1999). And hence, new alternative control methods are urgently needed. One recently proposed objective is to block malaria transmission by genetically modifying the mosquito vector to become incompetent to malaria transmission (Curtis, 1994; James et al., 1999; Kohoza et al., 2000). This objective is mainly relying upon utilising the vector immune system to kill malaria parasite in its gut. This malaria immuno-control strategy was originally based on the realization that the immune system of mosquitoes has potential responses to kill malaria parasite at several stages of its developmental cycle, specially the ookinetes (Collins et al., 1986; Paskewitz et al., 1988). These immune responses include the production of nitric oxide and antibacterial peptides that lyses ookinetes in the midgut wall (Luckhart et al., 1998; Shahabuddin et al., 1998a) and/or the melanization of both ookinetes and young oocysts (Paskewitz et al., 1988). These, in fact, are the mechanisms by which refractory mosquitoes block malaria development in the midgut in nature (Dimopoulos et al., 2001).
In malaria susceptible mosquitoes, it has been proven that midgut invasion by ookinetes triggers some immune responses in both midgut cells and systemically (Dimopoulos et al., 2001; Levashina, 2004), including up-regulation of the transcription of immune genes encoding defensin and nitric oxide (Luckhart et al., 1998; Dimopoulos et al., 2002). However, these induced responses are not effectively enough as they did not kill the ookinetes in the midgut. Moreover, the blood meal has been shown to significantly up-regulate the immune response (Koella and Sørensen, 2002; Ahmed, 2005a, b). However, also this up-regulation is inadequate to kill malaria parasite in the vector midgut. Based on these findings, the current study relies upon the suggestion that if such immune responses would sufficiently enhanced, they might efficiently block the malaria life cycle in mosquito vector.
On the other hand however, mounting immune responses were shown to be very costly in terms of reproductive fitness (Moret and Schmid-Hempel, 2000; Ahmed et al., 2002; Schwartz and Koella, 2004) to the extend that refractory mosquitoes have completely lost their ability to reproduce after 25 generation of successful blocking of malaria parasite (Somboon et al., 1999). Thus, in a previous study (Ahmed, 2004), a natural botanical extract, the Black Seed Oil (BSO), was tested as natural immune enhancing agent that showed significant reduction in the total number of fully developed oocysts in infected mosquitoes. However, the effect of this immune-enhancive natural extract on the concomitant reproductive cost of mounting immune responses still to be investigated.
Black seed, N. sativa, is a Dicotyledon belongs to the botanical family of Ranunculaceae, has been employed as protective and curative remedy for numerous disorders and is known to have many medicinal properties in traditional medicine (Chopra et al., 1956; Nadkarni, 1976). Most of studies on the biological effect of N. sativa have dealt with its crude extracts or its active principles, the quinones, thymoquinone and dithymoquinone (Daba and Abdel-Rahman, 1998; Nagi et al., 1999). N. sativa has also been reported to have various biological and antimicrobial activities (Worthen et al., 1998; Hanafy and Hatem, 1991). Recently, several studies have shown the enhancive effect of the black seed on the immune system of both vertebrates (Haq et al., 1999; El-Dakhakhny et al., 2002; Abdel-Ghaffar et al., 2003) and insects (Abu El-Magd, 1995; Ahmed, 2004).
Thus, the main goal of this study is not to show that induction of immunity in An. gambiae can be costly, since the concomitant reproductive cost and the mechanism behind it have already been established (Ahmed et al., 2002; Ahmed and Hurd, 2006). But rather, to investigate whether or not inducing non-costly immune responses is possible, which may be in support of the malaria immuno-control strategy. Hence, the BSO, was tested in this study through three conditions. The first, experimental mosquitoes were maintained on oral administration of 0.3% BSO all time. The second, since blood meal is an essential resource for mosquito reproduction (Clements, 1992) and enhances immunity (Chun et al., 1995; Schwartz and Koella, 2002; Ahmed, 2005a ,b), mosquitoes therefore were allowed to imbibe enough amounts of blood (until being folly engorged). The third, the induced immune responses, against lipopolysaccharide (LPS) and Sephadex® beads, as well as the concomitant follicular apoptosis and resorption were used as evaluation parameters for the dual effect of BSO.
MATERIALS AND METHODS
Mosquito Maintenance
The African malaria vector, An. gambiae (KIL strain), was maintained in standardized conditions in Vector Biology Laboratory at Keele University, UK in order to produce adults of similar sizes, as outlined in Ahmed et al. (1999). As mosquito size affects vitellogenesis (Briegel, 1990), experimental groups were therefore checked to verify similarity in body size (and hence, blood meal size) as detailed in Ahmed et al. (1999). No significant differences were detected in wing sizes of mosquito groups within treatments in this study (Table 1).
Oral Administration of Black Seed Oil (BSO)
Crude extract from N. sativa seeds was used as a source of BSO. Based on a previous study (Ahmed, 2004), 0.3% BSO (v/v) [mixed with 10% glucose solution (w/v)] was used for mosquito feeding. This mixture was being shacked vigorously several times a day to insure a homogenous mixture most of time. Experimental mosquitoes were allowed to feed on BSO-glucose mixture immediately after adult emergence and until the end of experiments. Mosquitoes were then allowed to blood feed prior to any further treatment, or left without blood feeding based on the experimental protocols (Fig. 1 and 2).
Test of BSO Effect on a Mosquito Cell-line
The toxicity of BSO was initially tested on Suakoko strain cells (Sua 4.0), a mosquito cell line derived from An. gambiae (Catteruccia et al., 2000), which was kindly provided by Hans-Michael Müller (The European Molecular Biology Laboratory, Heidelberg, Germany). Cells were maintained at 27°C in Schneider’s insect medium supplemented with 10% foetal bovine serum, 50 U mL-1 penicillin and 50 μg mL-1 streptomycin (Sigma-Aldrich). Cells were counted before use and cultured at 1x105 cells/100 μL medium under the microscope filter sterilized. BSO-glucose mixture (0.3% v/v) was added to each of 5 cellular samples (5 replicates, n = 5) of cellular medium (at 3.0 μL oil/1000 μL cellular medium each) to mimic the approximate concentration fed to experimental mosquitoes (0.3%). An amount of 3.0 μL from filter sterilized 10% glucose solution alone (0.3%) was added to each of another 5 samples as sham control. A third group of 5 samples were left un-treated as control. Cells were then incubated at 27°C for 24 h, after which, dead cells were visualized by adding 100 μL of Trypan blue (0.4%, Sigma-Aldrich) to each sample. Numbers of dead cells (stained blue) (Fig. 3B) were immediately counted and mean % of cellular death was calculated in each treated culture.
Test of BSO Effect on Ookinetes Development
In vivo Test (in mosquito midgut)
Effect of BSO on the development of ookinetes of Plasmodium yoelii nigeriensis in mosquito guts was tested. Mosquitoes were maintained on 0.3% BSO-glucose mixture from adult emergence and until the end of experiment. Six days old mosquitoes were starved for 12 h prior to feeding on gametocytemic mouse [infected with P. y. nigeriensis [Killick-Kendrick (N67)] as detailed in Ahmed et al. (1999). Fully engorged females were randomly selected and allowed access to 0.3% BSO-glucose mixture again until used for dissecting their midguts at 20 h post-feeding/infection. Each of 5 blood meals was individually expelled gently from the dissected gut onto a clean 1 mL eppendorf tube containing 50 μL Aedes Physiological Saline (APS) and gently dissolved. An amount of 10 μL of the resulting suspension was dropped on a clean slide, left for few minutes until air dried, fixed for 1 min in 90% methanol, rinsed with tap water and then air-dried before Giemsa's staining. Slides were then observed under phase contrast oil immersion (x1000 magnification). Ookinetes were counted in all the viewing fields across the slide (Fig. 4A). Means of 5 replicates (5 different counts from 5 different mosquitoes) of total number of ookinetes were counted and used for comparison.
In vitro Test (in ookinete culture)
A malaria infected mouse, having exflagellating microgametocytes (at least 5 exflagellation/microscopic thick blood-field) was anaesthetized prior to blood collection by cardiac puncture. Gametocytemic blood was used for in vitro ookinetes culture according to Al-Olayan et al. (2002). Briefly, infected blood was diluted 1 to 10 with sterile supplemented RPMI-1640 medium [25 mM Hepes, 2 mM glutamine supplemented with 50 mg of hypoxanthine, 2 g sodium bicarbonate, 10 mL antibiotic mixture, penicillin/streptomycin (penicillin, 5000 U/mL/streptomycin 5 mg mL-1)] (Sigma-Aldrech, UK) and 10% foetal bovine serum. The diluted blood culture was passed through a Whatman® CF11 column (Sigma-Aldrech, UK) to remove white blood cells in order to prevent attacking the parasite in the medium (Sinden et al., 1985). The purified blood medium was then divided into two subcultures (5 mL each). The first group was treated with filer-sterilized 0.3% BSO-glucose mixture and mixed very well by gentle pipetting up and down several times. The second group was treated with filer-sterilized 0.3% glucose solution (10% v/v) as control. The two cultures were then incubated for 20 h at 20°C (Sinden et al., 1985). The resulting mature banana-shaped ookinetes (Fig. 4B) were freshly counted in each culture using haemocytometer. Mean number of 5 replicates (from 5 different samples; n = 5) of ookinetes in each culture was counted and used for comparison.
Monitoring of Follicular Apoptosis (FA) and Follicular Resorption (FR)
Ten ovary pairs, from each treated mosquito groups, were dissected in Phosphate Buffered Saline (PBS) or APS for monitoring FA or FR at 18 or 24 h post-treatments, respectively. For monitoring FA, dissected ovaries were used for detecting a caspase-like activity using CaspaTagTM Fluorescein Caspase (VAD) (Intergen Company) according to manufacturer’s instructions and as detailed in Ahmed and Hurd (2006). Follicles that exhibiting caspase-like activity (undergoing apoptosis) were appear fluorescing and were easily counted under the fluorescent microscope. The % of apoptotic follicles was calculated from the total number of developing ovarian follicle in each ovary pair. For monitoring FR, dissected ovaries were immersed in Neutral Red [0.5% (w/v)] solution in citrate-phosphate buffer (0.1 moll-1 citric acid/0.1 moll-1sodium citrate) at pH 6 for 1 min to visualize resorped follicles (Bell and Bohm, 1975; Clements and Boocock, 1984) as detailed in Ahmed (2005a). The % of FA or FR was calculated/ovary pair (in 10 different mosquitoes from each treated group in each case; n = 10).
Experimental Protocols
Effect of BSO and Blood Meal on Humoral Antibacterial Activity, FA and FR
As shown in Fig. 1, six days-old mosquitoes (maintained on 0.3% BSO-glucose mixture) were starved for 12 h prior to blood feeding on an anaesthetized CD mouse, or left without blood feeding. After blood feeding, fully engorged females were randomly selected and assigned to sub-groups for the experimental procedures. Non-blood fed (glucose fed) or blood fed mosquitoes were injected with 0.25 μL of APS or LPS (10 ng/0.25 μL APS) into the thoracic haemocoel using a calibrated hand-made heat-pulled microcapillary needle as detailed in Ahmed et al. (2002). Injected mosquitoes were then maintained in small cages and allowed access to the 0.3% BSO-glucose mixture again until the end of experiment (Fig. 1). At 18 h post-injections, haemolymph were collected from 5 active mosquitoes (have the ability to fly) and used freshly for inhibition zone assay against M. luteus (NCTC 2665, Sigma, UK) as outlined in Ahmed et al. (2002).
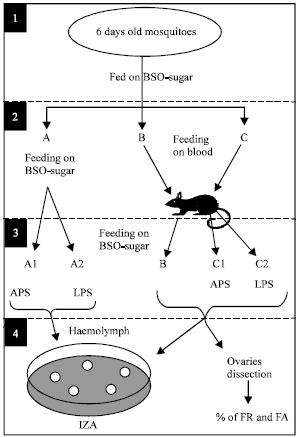 | |
| Fig. 1: | Experimental design for studying the effect of BSO and blood meal on the humoral response, FR and FA in An. gambiae |
| 1): | Six days old mosquitoes were used. |
| 2): | Mosquitoes were divided into groups A, B and C (≈ 100 mosquitoes each). Group A was kept feeding on BSO-glucose, while B and C were allowed to feed on mouse blood. |
| 3): | After feeding, groups A and C were divided into A1 and A2 and C1 and C2, respectively and immediately injected with APS or LPS as shown. Group B was kept without injection as control. All groups were then allowed access to BSO-glucose mixture again until 18 h later. |
| 4): | Haemolymph was collected from each group and used freshly for Inhibition Zone Assay (IZA). Ten mosquitoes from B, C1 or C2 were dissected for monitoring Follicular Apoptosis (FA) or Follicular Resorption (FR) at 18 or 24 h post-treatment, respectively |
Ten mosquitoes (n = 10) from each treated group were dissected for monitoring FA or FR at 18 or 24 h post-injection, respectively in parallel with control groups (Fig. 1) as detailed in Ahmed and Hurd (2006) and Ahmed (2005b), respectively.
Effect of BSO and Blood Meal on Melanization Response, FA and FR
As shown in Fig. 2, six days old mosquitoes (maintained on 0.3% BSO-glucose mixture) were starved for 12 h prior to blood feeding on an anaesthetized CD mouse, or left without blood feeding. Non-blood fed or blood fed mosquitoes were inoculated with negatively charged CM C-25 or neutral CM G-25 Sephadex® beads (Sigma-Aldrich, UK) suspended in APS as detailed in Ahmed (2005a). Only one bead (suspended in 0.25 μL APS) was inoculated into the thoracic haemocoel (one bead/mosquito) using a suitable calibrated hand-made microcapillary needle. Un-inoculated control group and APS-injected (0.25 μL APS/mosquito) (groups C1 and C2 in Fig. 2) were used as control and trauma control, respectively for FA and FR.
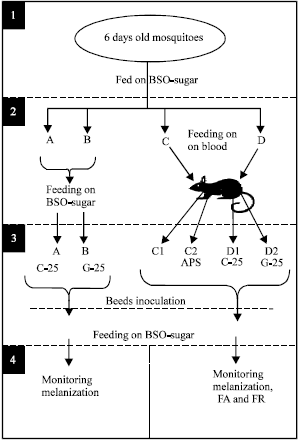 | |
| Fig. 2: | Experimental design of studying the effect of BSO and blood meal on the melanization response, FA and FR in An. gambiae |
| 1): | Six days old mosquitoes were used. |
| 2): | Experimental mosquitoes were divided into groups A, B, C and D (≈ 50 mosquitoes each). Group A and B were kept feeding on glucose-oil. C and D were allowed to feed on mouse blood |
| 3): | Fully engorged mosquitoes from group C were then subdivided into C1 and C2 (un-injected and APS-injected respectively, as controls for FA and FR). Group D was divided into D1and D2 and inoculated with C-25 and G-25 beads, respectively. All groups were then allowed to keep feeding on BSO-glucose mixture again for 18 h. |
| 4): | At 18 h later, mosquitoes were then dissected for monitoring melanization and Follicular Apoptosis (FA), or for monitoring Follicular Resorption (FR) at 24 h post-inoculation |
Beads inoculated mosquitoes were then maintained in small cages and allowed access to the 0.3% BSO-glucose mixture again (Fig. 2). Ten active mosquitoes (n = 10) were dissected at 18 h post-treatment for monitoring beads melanization. According to Koella and Sørensen (2002) and Ahmed (2005a), bead melanization was scored in three degrees: no visible melanization (white bead), patchy (dotted or partly melanized bead) and complete melanization (dark-black bead). Ten mosquitoes (n = 10) from each treated group were dissected for monitoring FA or FR, at 18 or 24 h post-injection, respectively, in parallel with control groups (Fig. 2) as detailed in Ahmed (2005a), Ahmed and Hurd (2006) and Ahmed (2005b), respectively.
Statistical Analysis
All statistical analyses were undertaken using MINITAB software (MINITAB, State College, PA, v. 13.1, 2001). Data of each experiment were first tested for normality and variances homogeneity prior to further analysis with the suitable test (mentioned when appropriate in the results). One-way ANOVA was used to determine the overall effects of the treatments within the normally distributed data that have homogeneous variances and individual comparisons within treatments were made using Tukey’s Pairwise comparison. Two-Sample t-test was used to analyse normally distributed data that had non-homogeneous variances. Kruskal-Wallis test was used to determine the overall effects of treatments prior to the individual comparisons using Mann-Whitney U for the non-parametric data (not normally distributed).
RESULTS
In this study, six days old mosquitoes (maintained on 0.3% BSO-glucose mixture) were used for inducing humoral activity against injected LPS or melanization response against inoculated Sephadex beads® in non-blood fed or blood fed mosquitoes. Experimental mosquitoes were verified to have similar body sizes (based on wing lengths measurements) within each experiment (Table 1).
| Table 1: | Wing length measurements of mosquitoes used in the study of humoral activity and melanization response respectively (Fig. 1 and 2). Wing lengths of mosquitoes were first measured to ensure similar body sizes before any further treatments. Data were first tested for normality using Anderson-Darling test prior to Mann-Whitney U test, which showed no significant differences in wing lengths between mosquito groups of each experiment |
 | |
 | |
| Fig. 3: | Mosquito cell line cultures [An. gambiae Suakoko strain (Sua 4.0)] 24 h post-incubation at 27°C in Schneider’s insect medium. A: represents an untreated and unstained culture. B: represents a Trypan Blue stained culture 24 h post-incubation/treatment with 0.3% BSO-glucose mixture (v/v), 0.3% glucose solution (10% w/v) (sham control) or untreated (control). Dead cells (stained blue, arrows) were counted in each treated culture and % of cellular death was calculated under the microscope |
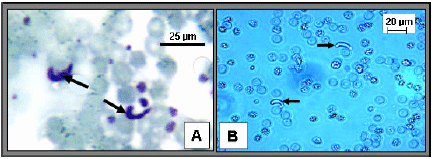 | |
| Fig. 4: | Mature ookinetes as they appear under the light microscope. A: represents Giemsa's stained mature ookinetes (arrows) prepared from blood meals of BSO-glucose fed mosquitoes 20 h post-infection. B: represents freshly prepared alive ookinetes (arrows) from BSO-glucose treated in vitro ookinetes culture 20 h post-treatment. Ookinetes were counted in control [treated with 0.3% glucose (10% w/v)] or treated cultures in each case |
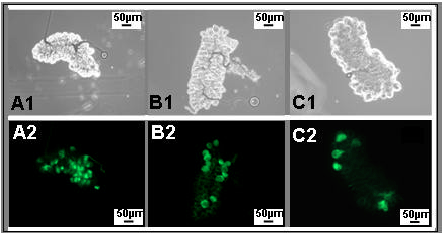 | |
| Fig. 5: | Follicular Apoptosis (FA) in ovaries of experimental mosquitoes as shown under a florescent microscope at 18 h post-LPS injection/blood feeding. A2 and B2 represent FA as a result of LPS injection in 10% glucose fed mosquitoes and 0.3% BSO-glucose fed mosquitoes, respectively. C2 represents FA in 0.3% BSO-glucose-fed mosquitoes after APS-injection. A1, B1 and C1 represent the same ovaries under normal light microscope |
Effect of BSO on Mosquito Cell-line
The mixture of 0.3% BSO-glucose was tested against the mosquito cell line An. gambiae Suakoko strain (Sua 4.0) (Fig. 3A). The 0.3% glucose solution alone (10%, v/v) was added to control cells. Five different counts (n = 5) from each treated culture were done in this experiment. Cells mortalities were 6.32±0.78, 6.44±0.87 and 5.6±0.89% within un-treated, glucose treated and BSO-glucose treated cultures, respectively (Plate 3B and Fig. 6). One-way ANOVA showed no significant difference in cells mortality within the three treatments (P>0.05, n = 5) (Fig. 6). This may indicate that the BSO has no toxic effect on mosquito cells.
Effect of BSO on Ookinetes Development in vivo and in vitro
Toxicity of BSO was tested against ookinetes development in vivo (in midgut) and in vitro (in ookinetes culture). Blood meals from malaria infected mosquitoes fed on 0.3% BSO-glucose mixture or 10% glucose solution showed a mean number of ookinetes of 105.80±7.47 or 120±7.49, respectively (Fig. 4A and Fig. 7).
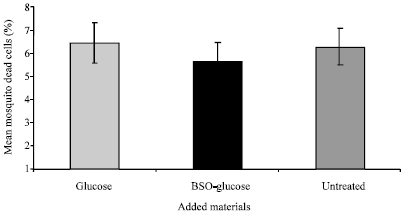 | |
| Fig. 6: | Effect of 0.3% BSO-glucose mixture on mosquito cell line [An. gambiae Suakoko strain (Sua 4.0)]. This histogram represents cells mortality after treatment with 0.3% of glucose solution (10% w/v) (sham control), 0.3% of BSO-glucose mixture (v/v) or untreated (control). Data were first tested for normality using Anderson-Darling test prior to One-way ANOVA which showed no significant difference in cell mortality between the three groups (P>0.5, n = 5). Error bars represent standard errors of means of 5 replicates |
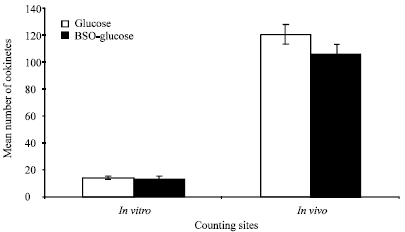 | |
| Fig. 7: | Effect of 0.3% BSO-glucose mixture on ookinetes development both in vivo and in vitro. For in vivo test, midguts of glucose- or BSO-glucose fed mosquitoes were dissected for counting ookinetes in their blood meals 20 h post-infection. For in vitro test, the purified medium of gametocytemic mouse-blood was treated with 0.3% of BSO-glucose mixture or glucose solution (10% v/v) before incubation for 20 h at 20°C. Ookinetes were then counted under the microscope in each culture. Data were first tested for normality using Anderson-Darling test prior to Tow-sample t-test which showed no significant difference in ookinetes number between treatments in both in vivo and in vitro tests (P>0.05, n = 5). Error bars represent the standard errors of means of 5 replicates |
Mann-Whitney U test showed no significant difference in the number of ookinetes counted in midguts of glucose fed and BSO-glucose fed mosquitoes (P>0.05, n = 5) (Fig. 7). Moreover, ookinetes in glucose-treated or BSO-glucose treated blood-ookinetes cultures showed a mean number of 14.40±0.92 or 13.80±1.83 ookinetes, respectively (Fig. 4B and Fig. 7). Two sample t-test showed no significant difference in ookinetes numbers between glucose-treated and BSO-glucose treated blood cultures (P>0.05, n = 5) (Fig. 7).
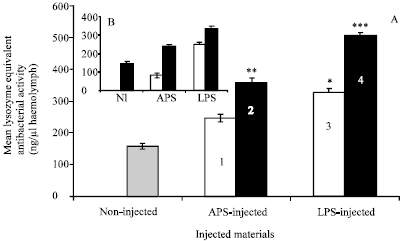 | |
| Fig. 8: | (A):Humoral anti-M. luteus activity equivalent to a standard lysozyme activity, induced by LPS injection in the malaria vector An. gambiae (maintained on 0.3% BSO-glucose mixture) at 18 h post-injection. BSO enhanced this activity in non-blood fed mosquitoes (white bars), in blood fed mosquitoes (black bars) and in blood fed non-injected control mosquitoes (dotted bar). Data were first tested for normality using Anderson-Darling test prior to One-way ANOVA. Error bars represent the standard errors of means of 5 replicates. *represents significant higher activity comparing to (1) and control. **represent significant higher activity comparing to (1) and control. ***represent significant higher activity comparing to 1, 2, 3 and control (Tukey’s Pairwise comparison, P<0.05). Group (1) showed significant higher activity comparing to control. (B): For comparison, the same activity in non-injected (NI) or APS and LPS-injected blood fed mosquitoes that maintained on 10% glucose solution only (Ahmed, 2005b) |
This may indicate that BSO has no effect on the development of ookinetes neither in vivo nor in vitro.
Effect of BSO and Blood Meal on Mosquito Response to LPS Injection
The Humoral Anti-M. luteus Activity
Mosquitoes used in this study were similar in body sizes as shown by Mann-Whitney U test on wing lengths (Table 1). The effect of BSO on the humoral anti-M. luteus activity induced by LPS injected into non-blood fed and blood fed mosquitoes was explored in this study as shown in (Fig. 1). Humoral activity against M. luteus was detected in haemolymph from non-injected mosquitoes (control) 18 h post-blood meal which was equivalent to the activity of 156.0±9.80 ng lysozyme (Fig. 8A). ANOVA showed that sham-injected non-blood fed mosquitoes have a significant increase in the humoral anti-M. luteus activity compared to blood fed un-injected control ones (244.60±10.90 v 156.0±9.80 ng) (Tukeys Pairwise comparison, P<0.5, n = 5) (Fig. 8A). Furthermore, LPS-injected non-blood fed mosquitoes showed a significant increase in this activity compared to sham-injected or control mosquitoes (324.00±14.40 v 244.60±10.90 or 156.0±9.80 ng, respectively) (Tukeys Pairwise comparison, P<0.5, n = 5) (Fig. 8A). In addition, blood meal enhanced this response as blood fed mosquitoes showed significant increase in this activity when injected with APS and LPS compared to those non-blood fed ones (356.70±14.90 v 244.60±10.90 and 505.78±7.80 v 324.00±14.40 ng, respectively) (Tukeys Pairwise comparison, P<0.5, n = 5) (Fig. 8A). This may indicate an enhancive effect for the BSO on the humoral activity which improved by the blood meal. As a comparison, non-blood fed or blood fed mosquitoes of this study (maintained on 0.3% BSO-glucose) showed 23 or 34% increase of humoral activity respectively compared to those with the same treatments of a previous study (maintained on 10% glucose solution only) (Fig. 8A compared to Fig. 8B (Ahmed, 2005b).
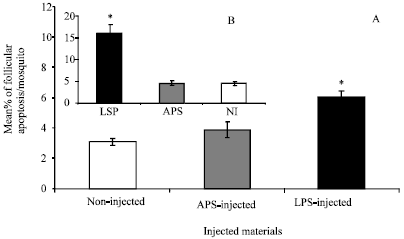 | |
| Fig. 9: | Follicular Apoptosis (FA) induced as a result of LPS-triggered immunity in BSO-fed mosquitoes compared to glucose fed ones 18 h post-injection. Histograms A and B represent the % of FA in BSO-glucose fed mosquitoes of this study and in glucose fed mosquitoes of a previous study (Ahmed and Hurd, 2006), respectively for comparison. In this study, FA was monitored in the ovaries of BSO-glucose-fed mosquitoes at 18 h post-APS injection, LPS injection or non-injected mosquitoes (A). Data were first tested for normality using Anderson-Darling test prior to tow-sample t-test. Error bars represent the standard errors of means of 10 replicates. *Significant higher compared to both sham-injected and control mosquitoes (P<0.05, n = 10) |
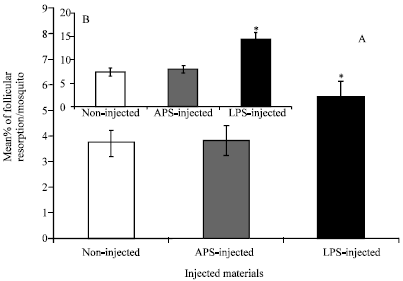 | |
| Fig. 10: | Follicular Resorption (FR) induced as a result of LPS-triggered immunity in BSO-fed mosquitoes 24 h post-injection. Histograms A and B represent the % of FR in BSO-glucose fed mosquitoes of this study and in glucose fed mosquitoes of a previous study (Ahmed 2005b), respectively for comparison. In this study, FR was monitored in the ovaries of BSO-glucose-fed mosquitoes at 24 h post-APS injection, LPS injection or non-injected mosquitoes (A). Data were first tested for normality using Anderson-Darling test prior to tow-sample t-test. Error bars represent the standard errors of means of 10 replicates. *Significant higher compared to both sham-injected and control mosquitoes (P<0.05, n = 10) |
This indicates that BSO may have enhanced the humoral activity against injected LPS.
The Concomitant FA and FR
In this experiment, ovaries from injected blood fed mosquitoes were dissected 18 h post-injections for monitoring FA (10 ovary pairs in each case) (Fig. 5). Non-injected mosquitoes (control) showed a % of 3.1±0.2 apoptotic follicles in their ovaries (Fig. 9A). Two-sample t-test showed no significant difference in the % of apoptotic follicles between sham-injected and control ones (03.9±0.50 v 03.1±0.2) (P>0.05, n = 10) (Fig. 9A). However, LPS-injected mosquitoes showed significant higher % of FA compared to control or sham-injected ones (6.05±0.43 v 3.1±0.2 or 3.9±0.5, respectively) (t-test, P<0.05, n = 10) (Fig. 5 and Fig. 9A). However, percentages of FA in LPS-injected mosquitoes here was 63% lower comparing to that of a previous study when mosquitoes were reared on 10% glucose solution only [Fig. 5 and Fig. 9A compared to Fig. 9B (Ahmed and Hurd, 2006)]. This indicates that BSO may have inhibited follicular apoptosis that induced as a result of humoral response.
Follicular resorption was also monitored in another mosquito samples within the same treated groups at 24 h post-injections (10 ovary pairs in each case). Control ovaries (from non-injected mosquitoes) showed 3.7±0.5% of the developing follicles undergoing resorption (Fig. 10A). Two-sample t-test showed no significant difference in the % of FR between sham-injected and control ones (3.82±0.57 v 3.7±0.5) (P>0.05, n = 10) (Fig. 10A). However, LPS-injection induced a significant higher % of FR compared to control or sham-injection (5.51±0.58 v 3.7±0.5 or 3.82±0.57, respectively) (P<0.05, n = 10) (Fig. 10A). As a comparison, these percentages of FR were relatively lower than that in mosquitoes reared up on 10% glucose solution only (50, 52.5 and 61.8% reduction in control, LPS-injected and LPS-injected mosquitoes respectively) [Fig. 10A compared to Fig. 10B (Ahmed, 2005b)]. These data indicate that BSO may have reduced follicular resorption induced as a result of humoral response.
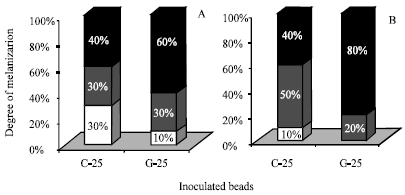 | |
| Fig. 11: | Melanization response in BSO-glucose fed mosquitoes shown against thoracic inoculated Sephadex® beads 18 h post-inoculation. A: represents melanization responses against beads in non-blood fed mosquitoes. B: represents melanization responses against beads inoculated immediately after a blood meal. Thoraces from each mosquito groups were dissected at 18 h post-inoculation for scoring the degrees of beads melanization. White bars represent the % of mosquitoes that showed un-melanized (white) beads. Gray bars represent the % of mosquitoes that showed patchy (half melanized) beads. Black bars represent the % of mosquitoes that showed completely melanized (black) beads |
Effect of BSO and Blood Meal on Mosquito Response to Inoculated Beads
Melanization Response
Percentages of mosquitoes showed the three melanization degrees were calculated (n = 10 mosquitoes for each) within non-blood fed (glucose fed) or blood fed groups. In non-blood fed groups, 30% of mosquitoes showed un-melanized negatively charged CM C-25 beads, while only 10% showed un-melanized neutral CM G-25 ones (Fig. 11A). Half-melanization was scored in 30% of mosquitoes against both kinds of beads. Complete melanization was shown in 40 or 60% of mosquitoes against CM C-25 or CM G-25, respectively (Fig. 11A). In blood fed mosquitoes however, only 10% of mosquitoes did not melanize CM C-25 beads, while no un-melanized CM G-25 bead was scored (Fig. 11B). Half melanization was shown in 50 or 20% of mosquitoes against CM C-25 or CM G-25 respectively (Fig. 11B). Whereas, complete melanization was shown in 40 or 80% of mosquitoes against CM C-25 or CM G-25, respectively (Fig. 11B).
The Concomitant FA and FR
In this experiment, 10 ovary pairs from treated or un-treated blood fed mosquito groups were dissected at 18 h post-treatment for monitoring FA. Control ovaries (from un-treated mosquitoes) showed 3.49% of their developing follicles undergoing apoptosis. ANOVA showed no significant difference in the percentage of follicles undergoing apoptosis between control and sham injected mosquitoes (3.49% v 3.33±0.82) (Tukeys Pairwise comparison, P<0.5, n = 10) (Fig. 12A). However, ovaries from C-25 or G-25 inoculated mosquitoes showed significant higher percentages of their developing follicles undergoing apoptosis than those of APS injected (7.3±0.82 v 3.33±0.82) or (6.2±0.7 v 3.33±0.82), respectively (Tukeys Pairwise comparison, P<0.5, n = 10) (Fig. 12A). No significant difference in the % of apoptotic follicles were shown between the ovaries of C-25 and G-25 inoculated mosquitoes. As a comparison, these percentages of FA were relatively lower, specially against the neutral CM G-25 beads as it reduced by 22% comparing to that of a previous study when mosquitoes were reared on 10% glucose solution only (Fig. 12A compared to Fig. 12B (Ahmed and Hurd, 2006).
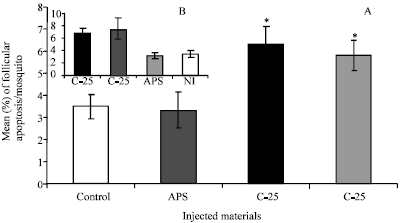 | |
| Fig. 12: | Follicular Apoptosis (FA) induced as a result of Sephadex® beads inoculation in BSO-glucose fed mosquitoes 18 h post-inoculation. Histograms A and B represent a comparison between the % of FA in BSO-fed mosquitoes of this study and that in 10% glucose fed mosquitoes of a previous study (Ahmed and Hurd, 2006) for comparison. In this study, FA was monitored in the ovaries of beads-inoculated mosquitoes 18 h post-C-25 and G-25 inoculation, APS injection (sham control) or non-injected mosquitoes (control). Data were first tested for normality using Anderson-Darling test prior to ANOVA. Error bars represent the standard errors of means of 10 replicates. * Significant higher compared to both sham-injected or control mosquitoes (Tukey’s Pairwise comparison, P<0.05, n = 10) |
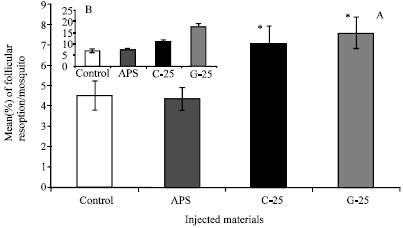 | |
| Fig. 13: | Follicular Resorption (FR) induced as a result of Sephadex® beads inoculation in mosquitoes 24 h post-inoculation/blood feeding. Histograms A and B represent a comparison between the % of FR in BSO fed mosquitoes in this study and that in glucose fed mosquitoes in a previous study (Ahmed, 2005a) respectively for comparison. In this study, FR was monitored in the ovaries of beads-inoculated mosquitoes 24 h post- C-25 and G-25 inoculation, APS injection (sham control) or non-injected mosquitoes (control). Data were first tested for normality using Anderson-Darling test prior to ANOVA. Error bars represent the standard errors of means of 10 replicates. * Significant higher compared to both sham-injected and control mosquitoes (Tukey’s Pairwise comparison, P<0.05, n = 10) |
This indicates that BSO may have reduced follicular apoptosis that induced as a result of melanization response.
Follicular resorption was monitored in another mosquito sample within the same group at 24 h post-injections (10 ovary pairs in each case; n = 10). Control or APS-injected mosquitoes showed 4.5±0.7 or 4.35±0.58% of FR per ovary pair, respectively (Fig. 13A). ANOVA showed significant higher FR in C-25 and G-25 inoculated mosquitoes compared to that in APS-injected ones (7.08±0.84% v 4.35±0.58% and 7.58±0.77% v 4.35±0.58%, respectively) (Tukeys Pairwise comparison, P<0.5, n = 10) (Fig. 13A). However, these percentages of FR were relatively lower compared to those in mosquitoes reared on 10% glucose solution only (i.e., 36.4 and 57% lower in C-25- and G-25-inoculated mosquitoes, respectively [Fig. 13A compared to Fig. 13B (Ahmed, 2005a).
DISCUSSION
In this study, it is important to clarify three points: the first; FA or FR were monitored 18 or 24 h post-blood feeding/treatment, respectively based on previous studies (Hopwood et al., 2001; Ahmed, 2005a, b). The second; ookinetes were counted 20 h post-blood meal as the number of ookinetes invading mosquito’s midgut has been shown to peak around 20 h post-infective blood meal (Ahmed, 2004). Thus, these time points were considered in the experimental designs of the study (mentioned when appropriate). The third; the size of the blood meal taken by mosquito may participate as a direct/indirect factor in most of the differences detected in treated mosquitoes of this study (i.e., differences in melanization degrees, humoral activity, FA and FR). Therefore, blood meal size was accounted for to be certain that these differences were not because of different sized blood meals. Thus, wing size (the distance from the wing tip to the distal end of the allula) was used as an estimate of body size (Table 1) and hence, blood meal size (Briegel, 1990).
Melanization and humoral activity were proved to be the most effective immune responses against malaria parasite in refractory mosquitoes (Collins et al., 1986; Shahabuddin et al., 1998b) and thus, they were studied here against the Sephadex® beads model and LPS, respectively. Blood fed mosquitoes, (maintained on 0.3% BSO-glucose mixture) showed significant increases in both melanization and humoral activity when compared to those blood fed ones (maintained on glucose solution only) (Ahmed, 2005a, b). This may be in support of Ahmed (2004) who recorded significant reduction in the number of mature oocysts in BSO-fed mosquitoes. This indicates that the reduction in oocysts number may have happened via BSO-induced immune response(s) that were sufficient to kill a significant number of developing ookinetes/young oocysts in midgut, not because of a toxic effect of BSO on these two stages. Evidence for this is the proved non-toxic effect of BSO on developing ookinetes both in vivo and in vitro shown in this study. Furthermore, BSO induced immune stimulation may refer to one or all of its active constituents (Daba and Abdel-Rahman, 1998; Nagi et al., 1999) and/or to its protein content (Haq et al., 1999). These data suggest that if a malaria-infective blood meal is synchronized with feeding on 0.3% BSO (the concentration used in this study), it may sufficiently induce immune responses to block malaria life cycle in mosquito midgut via enhancing the vector immunity.
On the other hand, the proved reproductive cost of mounting immune responses (Moret and Schmid-Hempel, 2000; Ahmed et al., 2002; Schwartz and Koella, 2004) is, in fact, a major limiting factor to the malaria immuno-control strategy. This kind of cost occurs via triggering the FA which leads to FR in the ovaries of immunized mosquitoes (Ahmed 2005a, b; Ahmed and Hurd, 2006) and hence, reduce fecundity (Ahmed et al., 2002). In the evolutionary point of view, this cost may indicate a life-history strategy exists whereby a maximum fitness is achieved by balancing competing needs via re-partitioning of nutritional resources between reproduction and immunity (Moret and Schmid-Hempel, 2000; Schwartz and Koella, 2004). Moreover, the current study and others (Ahmed, 2005a, b; Ahmed and Hurd, 2006), showed that stimulation of both melanization and humoral arms of the mosquito immune system resulted in the induction of apoptotic pathways in the developing ovarian follicles leading to FR. This is likely to be the major mechanism behind fecundity reduction that may be due to the allocation of some resources towards melanizing the inoculated beads which, on the other hand, more likely to occur on the cost of egg formation (since fewer resources might have been directed to egg production). This kind of resource re-allocation is more likely to be the direct result of FA and hence, FR not because of a toxic effect of the injected materials on follicular development. Evidences for this are: (a) no significant difference in mortality between LPS/beads-injected and sham-injected mosquitoes occurred (data not shown) and (b) a toxicity test has showed no toxic effect for LPS on the mosquito cell line when applied in the same dose used in this study (Ahmed and Hurd, 2006). Moreover, differences in melanization degrees monitored against beads of different charges, did not result in concomitant differences in FA and FR, which may indicate a non-dose-related response. Evidence for this has been provided by Ahmed et al. (2002) when they demonstrated that humoral activity induced by LPS injection was dose-dependent while fecundity reduction was not.
It is well-known that apoptosis, a process leading to cell death, plays an important role in the control of various biological systems, such as immune responses, hematopoiesis, embryonic development and carcinogenesis in vertebrates (Raff, 1992). In insects, oogenesis is one of the most important processes that controlled by apoptosis (Cavaliere et al., 1998; Buszczak and Cooley, 2000). In mosquitoes, regulation of egg batch size occurs naturally according to the availability of nutrients. Thus FR was found to occur when inadequate blood meal is imbibed by mosquito (Clements and Boocock, 1984). Several studies have shown that FR has a link to the occurrence of apoptosis in the follicular epithelial cells and provided evidences that FA initiates FR (Hopwood et al., 2001; Uchida et al., 2004). Jeong and Kim-Ha (2003) demonstrated that the RNA binding protein (RBP9), expressed in the ovarian germline cells of Drosophila, is involved in the regulation of particular genes that function in the differentiation of ovarian cells. They also proved that over-expression of this protein during oogenesis did cause apoptosis in the developing follicles of this fly. Based on these findings, it would be suggested that immune induction could have directly/indirectly triggered over-expression of this protein by somehow and subsequently, induced increase in FA that monitored here in this study. Besides, a direct link between immune induction and apoptosis has been suggested by Georgel et al. (2001) who provided the immune deficiency gene (imd) [of the antibacterial activity in Drosophila (Lemaitre et al., 1995) as an evidence for this link. They demonstrated that imd gene encodes a 30 kDa protein with a death domain and that, over-expression of this gene triggers apoptosis as it is involved in the apoptotic response to the DNA damage (Brachmann et al., 2000; Zhou et al., 1999). They also reported that this apoptotic effect is controlled by another kind of protein, the anti-apoptotic baculovirus protein (P35) (Hay et al., 1994). If this is the case in mosquitoes, it would be suggested that BSO-reduced FA may have occurred via a tight control of RBP9 and P35 transcriptions and thus, proper follicular development has maintained. Evidences for this are, on one hand, the black seeds have been shown to prevent the progressive apoptosis in Hep-2, laryngeal carcinoma cells, that were pre-treated with apoptosis-inducing factors (Corder et al., 2003). On the other hand, it has been proven that BSO is a protective agent against the chromosomal fragmentation induced in mouse cells (Aboul-Ela, 2002). Since chromosomal fragmentation has been reported as a sign of FA in mosquitoes (Hopwood et al., 2001), it would be assumed that BSO may have the same effects in mosquitoes and thus, normal follicular development in immunised mosquitoes was maintained.
In addition, Koella and Sørensen (2002) proved that the adequate nutritional resources resulted in complete melanization of C-25 by An. stephensi compared to inadequately-fed ones. Taking this into consideration and based on the current data, it would be suggested that BSO-glucose mixture may significantly increased immune responses in blood fed mosquitoes via providing adequate nutrition for immunity as well as for compensating the concomitant shrinkage of nutrition allocated for vitellogenesis. And hence, both immunity and vitellogenesis (in terms of normal FA an FR processes) occurred efficiently in parallel. Thus, beside being immuno-enhancive, the BSO also reduced the concomitant reproductive costs of immune stimulation and thus, maintained normal FA and FR comparing to those in immunized glucose fed mosquitoes (Ahmed, 2005a, b). Hence, this dual effect of BSO, as well as not being toxic to mosquito, may suggest it as a suitable candidate in support of the immuno-control strategy. Finally, since crude BSO has several active ingredients, the quinones, thymoquinone, nigellone and dithymoquinone (Daba and Abdel-Rahman 1998; Nagi et al., 1999), which activate the vertebrate immune system, it is still to be investigated whether or not all or one of these ingredients is responsible for immune induction in mosquitoes.
ACKNOWLEDGMENTS
Great thanks is due to Prof. Hilary Hurd (Parasitology and Vector Biology Laboratory, Biological Sciences Department, School of Life Sciences, Keele University, Keele, UK) for allowing me to use her laboratory facilities to achieve this study during one of my visits to her research group. Many thanks to her lab technicians for their supports and help in rearing mosquitoes. Thanks to Prof. Hans-Michael Müller (The European Molecular Biology Laboratory, Heidelberg, Germany) for providing the An. gambiae cell line used in this study.
REFERENCES
- Aboul-Ela, I.E., 2002. Cytogenetic studies on Nigella sativa seeds extract and thymoquinone on mouse cells infected with schistosomiasis using karyotyping. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 516: 11-17.
CrossRefDirect Link - Ahmed, A.M., 2004. Activation of the immune system of Anopheles gambiae against malaria parasite: A comparison between bacterial injection and a botanical extract. Proceedings of the International Conference of Biological Sciences, April 28-29, 2004, Faculty of Sciences, Tanta University, Tanta, Egypt, pp: 122-141.
- Ahmed, A.M., 2005. Melanization of sephadex beads by the malaria vector, Anopheles gambiae: Effect of blood meal and mechanisms of reproductive costs. J. Egypt. Ger. Soc. Zool., 47E: 69-85.
Direct Link - Ahmed, A.M., 2005. The humoral anti-bacterial response of Anopheles gambiae and the immunity-reproduction trade-off: Between the hope and limitation of the malaria immuno-control strategy. Proceedings of the 3rd International Conference of Applied Entomology, March 23-24, 2005, Cairo University, pp: 351-374.
- Ahmed, A.M., P. Taylor, R. Maingon and H. Hurd, 1999. The effect of Plasmodium yoelii nigeriensis on the reproductive fitness of Anopheles gambiae. Invertebr. Reprod. Dev., 36: 217-222.
Direct Link - Ahmed, A.M., S. Baggott, R.L.Maingon and H. Hurd, 2002. The costs of mounting an immune response are reflected in the reproductive fitness of the mosquito Anopheles gambiae. Oikos, 97: 371-377.
Direct Link - Ahmed, A.M. and H. Hurd, 2006. Immune stimulation and malaria infection impose reproductive costs in Anopheles gambiae via follicular apoptosis. Microbes Infec., 8: 308-315.
PubMedDirect Link - Al-Olayan, E.M., A.L. Beetsma, G.A. Butcher, R.E. Sinden and H. Hurd, 2002. Complete development of mosquito phases of the malaria parasite in vitro. Science, 295: 677-679.
Direct Link - Brachmann, C.B., O.W. Jassim, B.D. Wachsmuth and R.L. Cagan, 2000. The Drosophila bcl-2 family member dBorg-1 functions in the apoptotic response to UV-irradiation. Curr. Biol., 10: 547-550.
Direct Link - Briegel, H., 1990. Fecundity, metabolism and body size in Anopheles (Diptera: Culicidae), vectors of malaria. J. Med. Entomol., 27: 839-850.
CrossRefPubMedDirect Link - Buszczak, M. and L. Cooley, 2000. Eggs to die for: Cell death during Drosophila oogenesis. Cell Death Differ., 7: 1071-1074.
PubMedDirect Link - Catteruccia, F., T. Nolan, C. Blass, H.M. Muller, A. Crisanti, F.C. Kafatos and T.G. Loukeris, 2000. Toward Anopheles transformation: Minos element activity in anopheline cells and embryos. Proc. Natl. Acad. Sci. USA., 97: 2157-2162.
PubMedDirect Link - Cavaliere, V., C. Taddei and G. Gargiulo, 1998. Apoptosis of nurse cells at the late stages of oogenesis of Drosophila melanogaster. Dev. Genes Evol., 208: 106-112.
CrossRefDirect Link - Chun, J., M. Riehle and S.M. Paskewitz, 1995. Effect of mosquito age and reproductive status on melanization of Sephadex beads in Plasmodium-refractory and -usceptible strains of Anopheles gambiae. J. Invertebr. Pathol., 66: 11-17.
PubMedDirect Link - Clements, A.N. and M.R. Boocock, 1984. Ovarian development in mosquitoes: Stages of growth and arrest and follicular resorption. Physiol. Entomol., 9: 1-8.
CrossRefDirect Link - Collins, F.H., R.K. Sakai, K.D. Vernick, S. Paskewitz and D.C. Seeley et al., 1986. Genetic selection of a Plasmodium-refractory strain of the malaria vector Anopheles gambiae. Science, 234: 607-610.
CrossRefDirect Link - Corder, C., H. Benghuzzi, M. Tucci and Z. Cason, 2003. Delayed apoptosis upon the treatment of Hep-2 cells with black seed. Biomed. Sci. Instrum., 39: 365-370.
PubMedDirect Link - Daba, M.H. and M.S. Abdel-Rahman, 1998. Hepatoprotective activity of thymoquinone in isolated rat hepatocytes. Toxicol. Lett., 95: 23-29.
CrossRefDirect Link - Dimopoulos, G., H.M. Muller E.A. Levashina and F.C. Kafatos, 2001. Innate immune defense against malaria infection in the mosquito. Curr. Opin. Immunol., 13: 79-88.
PubMedDirect Link - Dimopoulos, G., G.K. Christophides, S.J. Schultz, K.P. White, C. Barillas-Mury and F.C. Kafatos, 2002. Genome expression analysis of Anopheles gambiae: Responses to injury, bacterial challenge and malaria infection. Proc. Natl. Acad. Sci. USA., 99: 8814-8819.
PubMedDirect Link - El-Dakhakhny, M., N.J. Madi, N. Lembert and H.P. Ammon, 2002. Nigella sativa oil, nigellone and derived thymoquinone inhibit synthesis of 5-lipoxygenase products in polymorphonuclear leukocytes from rats. J. Ethnopharmacol., 81: 161-164.
Direct Link - Georgel, P., S. Naitza, C. Kappler, D. Ferrandon, D. Zachary, C. Swimmer, C. Kopczynski, G. Duyk, J.M. Reichhart and J.A. Hoffmann, 2001. Drosophila Immune Deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev. Cell, 1: 503-514.
PubMedDirect Link - Hanafy, M.S.M. and M.E. Hatem, 1991. Studies on the antimicrobial activity of Nigella sativa seed (black cumin). J. Ethnopharmacol., 34: 275-278.
CrossRefDirect Link - Haq, A., P.I. Lobo, M. Al-Tufail, N.R. Rama and S. Al-Sedairy, 1999. Immunomodulatory effect of Nigella sativa proteins fractionated by ion exchange chromatography. Int. J. Immunopharmacol., 21: 283-295.
CrossRefPubMedDirect Link - Hay, B.A., T. Wolff and G.M. Rubin, 1994. Expression of baculovirus P35 prevents cell death in Drosophila. Development, 120: 2121-2129.
Direct Link - Hopwood, J.A., A.M. Ahmed, A. Polwart, G.T. Williams and H. Hurd, 2001. Malaria-induced apoptosis in mosquito ovaries: A mechanism to control vector egg production. J. Exp. Biol., 204: 2773-2780.
Direct Link - James, A.A., B.T. Beerntsen, M. de L. Copurro, C. J. Caotes, J. Coleman, N. Jasinskiene and A.U. Krettli, 1999. Controlling malaria transmission with genetically-engineered, Plasmodium-resistant mosquitoes: Milestones in a model system. Parasitology, 41: 461-471.
PubMedDirect Link - Jeong, K. and J. Kim-Ha, 2003. Expression of Rbp9 during mid-oogenesis induces apoptosis in egg chambers. Mol. Cells, 16: 392-396.
Direct Link - Kohoza, V., A. Ahmed, W-L. Cho, N. Jasinskiene, A.A. James and A. Raikhel, 2000. Engineering blood meal-activated systemic immunity in the yellow fever mosquito, Aedes aegypti. Proc. Nat. Acad. Sci. USA., 97: 9144-9149.
Direct Link - Koella, J.C. and F.L. Sørensen, 2002. Effect of adult nutrition on the melanization immune response of the malaria Vector Anopheles gambiae. Med. Veter. Entomol., 16: 316-320.
Direct Link - Lemaitre, B., E. Kromer-Metzger, L. Michaut, E. Nicolas, M. Meister, P. Georgel, J.M. Reichhart and J.A. Hoffmann, 1995. A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defence. Proc. Natl. Acad. Sci. USA., 92: 9465-9469.
Direct Link - Levashina, E.A., 2004. Immune responses in Anopheles gambiae. Insect Biochem. Mol. Biol., 34: 673-678.
CrossRefDirect Link - Luckhart, S., C.Y. Vodovotz and L.R. Rosenberg, 1998. The mosquito Anopheles stephensi limits malaria parasite development with inducible synthesis of nitric oxide. Proc. Natl. Acad. Sci. USA., 95: 5700-5705.
Direct Link - Manfredi, C., 1999. Can the resurgence of malaria be partially attributed to structural adjustment programmes?. Parassitology, 41: 389-390.
PubMedDirect Link - Moret, Y. and P. Schmid-Hempel, 2000. Survival for immunity: The price of immune system activation for bumblebee workers. Science, 290: 1166-1168.
CrossRefDirect Link - Nagi, M.N., K. Alam, O.A. Badary, O.A. Al-Shabanah, H.A. Al-Sawaf and A.M. Al-Bekairi, 1999. Thymoquinone protects against carbon tetrachloride hepatotoxicity in mice via an antioxidant mechanism. IUBMB Life, 47: 153-159.
CrossRefDirect Link - Paskewitz, S.M., M.R. Brown, A.O. Lea and F.A. Collins, 1988. Ultrastructure of the encapsulation of Plasmodium cynomolgi (B strain) on the midgut of a refractory strain of Anopheles gambiae. Parasitology, 74: 432-439.
CrossRefDirect Link - Sinden, R.E., R.H. Hartley and L. Winger, 1985. The development of Plasmodium ookinetes in vitro: An ultrastructural study including a description of meiotic division. Parasitology, 91: 227-244.
CrossRefDirect Link - Schwartz, A. and J.C. Koella, 2002. Melanization of Plasmodium falciparum and C-25 Sephadex beads by field caught Anopheles gambiae (Diptera: Culicidae) from Southern Tanzania. J. Med. Entomol., 39: 84-88.
PubMedDirect Link - Schwartz, A. and J.C. Koella, 2004. The cost of immunity in the yellow fever mosquito, Aedes aegypti depends on immune activation. J. Evol. Biol., 17: 834-840.
PubMedDirect Link - Shahabuddin, M., S. Cociancick and H. Zieler, 1998. The search for novel malaria transmission-blocking targets in the mosquito midgut. Parasitol. Today, 14: 493-496.
PubMedDirect Link - Shahabuddin, M.I. P. Fields, J.A. Bulet, Hoffman and L.H. Miller, 1998. Plasmodium gallinaceum differential killing of some mosquito stages of the parasite by insect defensin. Exp. Parasitol., 89: 103-112.
CrossRefDirect Link - Somboon, P., L. Prapanthadara and W. Suwonkerd, 1999. Selection of Anopheles dirus for refractoriness and susceptibility to Plasmodium yoelii nigeriensis. Med. Vet. Entomol., 13: 355-361.
PubMedDirect Link - Uchida, K., M. Nishizuka, D. Ohmori, T. Ueno, Y. Eshita and A. Fukunaga, 2004. Follicular epithelial cell apoptosis of atretic follicles within developing ovaries of the mosquito Culex pipiens pallens. Insect Physiol., 50: 903-912.
Direct Link - Worthen, D.R., O.A. Ghosheh and P.A. Crooks, 1998. The in vitro anti-tumor activity of some crude and purified components of blackseed, Nigella sativa L. Anticancer Res., 18: 1527-1532.
PubMed - Zhou, L., Z. Song, J. Tittel and H. Steller, 1999. HAC-1, a Drosophoila homolog of APAF-1 and CED-4 functions in developmental and radiation-induced apoptosis. Mol. Cell, 4: 745-755.
PubMedDirect Link