
Azza A. Shehata
Department of Botany, Faculty of Science, Alexandria University, Egypt
International Journal of Botany
Year: 2008 | Volume: 4 | Issue: 1 | Page No.: 67-76







ABSTRACT
Pollen morphology of sixteen species including two subspecies and representing four genera: Geranium L., Erodium L., Monsonia L. and Pelargonium L´ Hér. of Geraniaceae in Egypt were investigated by the aid of light and scanning electron microscopy. The study showed that pollen grains were radially symmetrical, isopolar, prolate spheroidal to oblate spheroidal, tricolp (ate) orate. Tectum was generally striate/reticulate, reticulate/gemmate or reticulate. On the basis of pollen morphological characters, principally aperture types and exine sculpture, pollen grains were separated into three types and three subtypes representing different taxonomic categories. A key to the different pollen types and subtypes was provided. The results demonstrated that pollen morphological characters, as pollen shape, size, aperture characters as well as exine sculpture and structure, were taxonomically significant at the generic level and to some extent at the specific level. Moreover, the results confirmed the eurypalynous nature of the family.
PDF Abstract XML References Citation
How to cite this article
Azza A. Shehata, 2008. Pollen Morphology of Egyptian Geraniaceae: An Assessment of Taxonomic Value. International Journal of Botany, 4: 67-76.
DOI: 10.3923/ijb.2008.67.76
URL: https://scialert.net/abstract/?doi=ijb.2008.67.76
DOI: 10.3923/ijb.2008.67.76
URL: https://scialert.net/abstract/?doi=ijb.2008.67.76
INTRODUCTION
Geraniaceae is a cosmopolitan family of mostly temperate and subtropical annual or perennial herbs and a few small shrubs, comprising about 750 species belonging to five genera: Erodium L` Her., Geranium L., Monsonia L., Sarcocaulon (DC.)Sweet and Pelargonium L`Her. (Hutchinson, 1969; Mabberley, 1997). The family is subdivided by Hutchinson (1969) into two tribes: Geranieae with primarily actinomorphic flowers (Erodium, Geranium, Monsonia and Sarcocaulon) and Pelargonieae with zygomorphic flowers (Pelargonium). Geraniaceae has been studied by several authors utilizing different criteria, as floral anatomy (Al-Nowaihi and Khalifa, 1973); nectaries (Link, 1991); chloroplast DNA variations (Price et al., 1990); rbcL sequence comparisons (Price and Palmer, 1993); comparative karyological studies (Albers, 1990); Floral morphological characters (Aldasoro et al., 2002) etc.
The application of pollen morphology in the recognition, identification and interpretation of relationships of plants at various hierarchical levels in plant taxonomy and production of pollen keys has been reported in many angiosperm families (Erdtman, 1966; Mulder, 2003; Huysmans et al., 2003; Al-Quran, 2004). Moreover, the advent of scanning techniques has greatly enabled the study of pollen morphology with great precision, thereby making their application much more efficient and dependable.
The pollen morphology of Geraniaceae or some of its representatives has been studied by several researchers Bortenschlager (1967), El-Oqlah (1983, 1989), Verhoeven and Venter (1987), Stafford and Blackmore (1991) and Stafford and Gibby (1992). Recently, Aedo et al. (2007) have used the palynological data in combination with morphological data in their taxonomic revision of genus Geranium.
In Egypt, Geraniaceae is represented by twenty five species belonging to three genera namely: Erodium, Geranium and Monsonia (Tackholm, 1974; Boulos, 2000). As far as the author is aware, the systematic investigation of the pollen morphology of Geraniaceae in Egypt had not been studied comprehensively, although some of its pollen was occasionally identified from neighboring countries (El-Ghazaly, 1991). The main objective of the present investigation was to investigate and describe the pollen morphology of some of the taxa of wild and cultivated Geraniaceae growing in Egypt and to show how far the pollen morphological variations could be used to distinct between the taxa studied.
MATERIALS AND METHODS
The study is based on the pollen morphology of 16 species including two subspecies of wild and cultivated Egyptian representatives of Geraniaceae.
Pollen samples of each studied species were collected from living collections as well as from herbarium specimens kept in the herbaria of Alexandria University and the Agricultural Museum (Table 1). Identification and plant name followed Boulos (2000). The pollen grains were prepared for light (LM) and Scanning Electron Microscopy (SEM) by the standard method described by Erdtman (1960). For LM studies, pollen grains were mounted on glycerin jelly onto glass slides. For examination by SEM, dried pollen grains were mounted onto stubs with double sided tape. The specimens were coated with gold using a Fine Coat Sputter JFC 1100 and scanned using the JEOL JSM 5300 SEM, at 15 and 25 KV accelerating voltage, at Electron Microscopy Unit, Alexandria University.
| Table 1: | Collection data of the investigated specimens |
 | |
Pollen diameter, polar axis (P) and equatorial diameter (E) were measured on at least 20 mature pollen grains with LM using oil objective lens and P/E was calculated. Lumina and muri were measured in the middle of the mesocolpia, chiefly from digital SEM images; by using a minimum sample of two sets of 10 measurements from each SEM graphs. The terminology employed in this investigation follows that of Erdtman (1969) and Punt et al. (1994).
RESULTS
The palynological data of the investigated taxa are shown in Table 2.
General pollen morphological characters: The present results revealed that pollen grains were usually monad, isopolar, radially symmetrical. The mean value of polar axis (P) ranged from 44 μm in Pelargonium grandiflorum to 111 μm in Erodium gruinum, while the mean value of equatorial diameter (E) ranged from 32 μm in Pelargonium grandiflorum to 106 μm in Erodium gruinum. The grains were circular to slightly triangular with convex sides in polar view and circular to elliptic in equatorial view. Apertures in the studied taxa were tricolp(ate)orate. Tectum showed variable sculptures (reticulate, striate/reticulate and reticulate/gemmate).
The studied pollen grains were found to be categorized in terms of exine sculpture into three pollen types and three subtypes summarized as follows:
Type I: Striate/Striate-Reticulate type:
Species: Erodium cicutarium, E. ciconium, E. crassifollium, E. glaucophyllum, Erodium gruinum, E. laciniatum subspecies laciniatum and subspecies pulverulentum, E. malacoides and E. moschatum.
Shape: Oblate spheroidal to prolate spheroidal.
Pollen class: Tricolporate.
Exine ornamentation: Complex and consists of three layers. The first layer corresponding to the sexine (1) is formed by the apices of the columellae, above which there is a semitectate reticulum (sexine 2). The uppermost layer (sexine 3) consists of long or short winding striae which form a distinct separate layer. In E. moschatum, the striate elements appear to be more interwoven, curved and fused with the underlying muri of the reticulum beneath. In E. malacoides, the striae were unbranched and closely spaced, while they were branched, sparsely spaced and running at various directions in E. cicutarium, E. ciconium, E. laciniatum subspecies laciniatum and subspecies pulverulentum, E. gruinum, E. crassifollium and E. glaucophyllum.
Within this pollen type, it was not possible to sharply distinguish between the species (Fig. 1-16).
| Table 2: | Pollen morphological data of the studied taxa of Geraniaceae |
 | |
| *: Mean values followed by range in parenthesis | |
Type II. Reticulate/Gemmate type:
Species: Geranium molle, G. trilophum and G. dissectum
Shape: Oblate spheroidal to prolate spheroidal
Pollen class: Tricolporate
Exine ornamentation: Tectum reticulates. However it is heavily ornamented with bacula and gemmae. Supratectal processes were of variable sizes. Two main forms of gemmae were distinguished: Bifid large gemmate projections with distinct shallow striations and smaller rod like bacula with slightly swollen head. Intermediate-sized processes occur between these two forms but were relatively few in number. G. dissectum is distinguished from the rest of species by its small size range (Table 2) (Fig. 17-22).
Type III: Reticulate type: In this type, tectum was simple and consisting of a closely defined reticulum which was hetero-or homobrochate. Lumina were regular or irregular in shape with sparsely or closely spaced baculae. Muri that were linking tops of columellae were either smooth or beset with supratectal elements of various shapes. According to the nature of muri the reticulate pollen type is subdivided into three subtypes (Fig. 23-30).
Subtype A: Monsonia heliotropioides subtype:
Species: Monsonia heliotropioides and M. nivea
Shape: Oblate spheroidal
Pollen class: Tricolpate
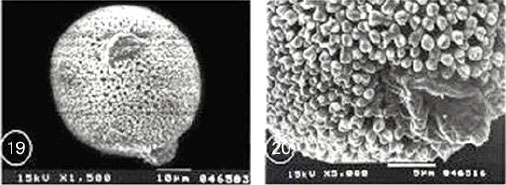
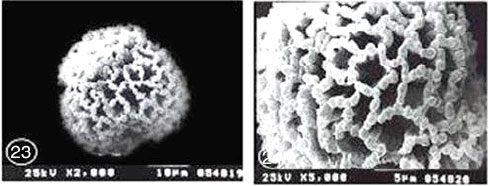
Exine ornamentation: Coarsely reticulate. The reticulum was heterobrochate. The average area of lumina was 5.22 μm2 for Monsonia heliotropioides and 2.47 μm2 for M. nivea. The surface of nexine was perforated. Muri was articulate, fragmentimurate, simplibaculate. Baculae were closely spaced (sometimes fused), relatively stout and interconnected by muri which were provided with knob like elements of varying shape and size. The mean value of muri width was 1.01 μm for M. heliotropioides and 0.97 μm for M. nivea (Fig. 23-26).
Subtype B: Monsonia senegalensis subtype:
Species: Monsonia senegalensis.
Shape: Oblate spheroid.
Pollen class: Tricolpate.
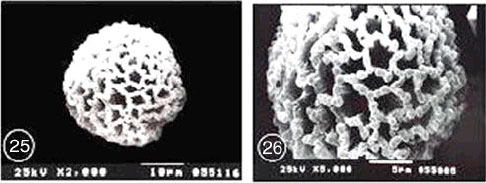
Exine ornamentation: Coarsely reticulate. The reticulum was generally homobrochate, characterized by five to six angled wide lumina. The mean of lumina area was 4.42 μm2, each muri was supported by one to four columellae. The surface of nexine was slightly granular at high magnification in SEM. Baculae were interconnected by straight sided and smooth muri. The mean of muri width was 0.63 μm. In some parts, the reticulum was generally level, but some muri may dip in the centre forming peaks at some of the supporting columellae (Fig. 27 and 28).
Subtype C: Pelargonium grandiflorum subtype:
Species: Pelargonium grandiflorum
Shape: Prolate
Pollen class: Tricolpate
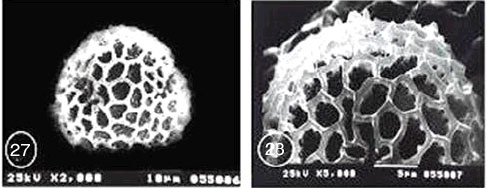
Exine ornamentation: Reticulate, lumina generally angular in shape. The measure of lumina area was 1.5 μm2. The surface of nexine was smooth. The mean of muri width was 0.60 μm. Muri were laterally compressed and somewhat undulating giving the surface a slightly striate appearance (Fig. 29 and 30).
 |
 |
 |
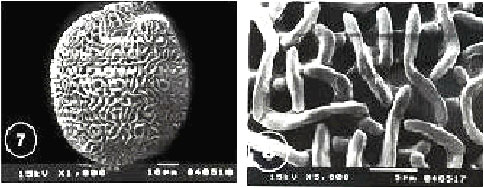 |
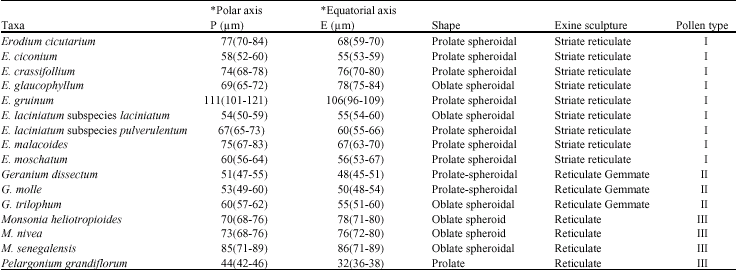
| Fig. 1-8: SEM micrographs of pollen morphology and exine sculpture of studied taxa of Geraniaceae |
| Fig. 1: Erodium cicutarium, equatorial view |
| Fig. 2: E. cicutarium, exine pattern |
| Fig. 3: E. ciconium, equatorialand polar view |
| Fig. 4: E. ciconium, exine pattern |
| Fig. 5: E. crassifolium, polar view |
| Fig. 6: E. crassifolium, exinepattern |
| Fig. 7: E. gruinum, polar viewFig. |
| Fig. 8: E. gruinum, exine pattern |
 |
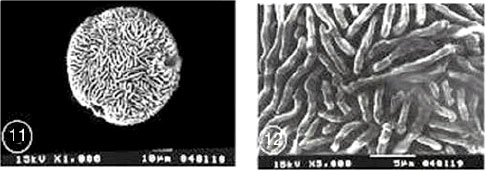 |
 |
 |
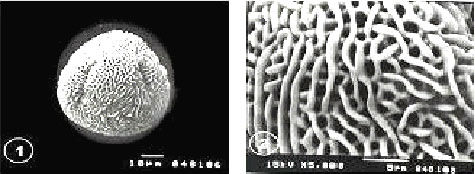
| Fig. 9-16: SEM micrographs of pollen morphology and exine sculpture of studied taxa of Geraniaceae |
| Fig. 9: E. laciniatum subspecies laciniatum, equatorial and polar view |
| Fig. 10: E. laciniatum subspecies laciniatum, exine pattern |
| Fig. 11: E. laciniatum subspecies pulveruntum, equatorial view |
| Fig. 12: E. laciniatum subspecies pulveruntum exine pattern |
| Fig. 13: E. malocoides,polar view |
| Fig. 14: E. malocoides, exine pattern |
| Fig. 15: E. moschatum, polar view |
| Fig. 16: E. moschatum exine pattern |
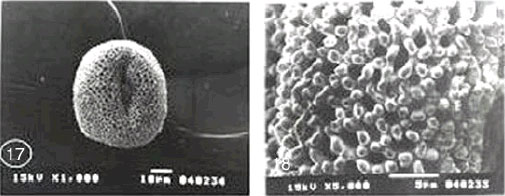 |
 |
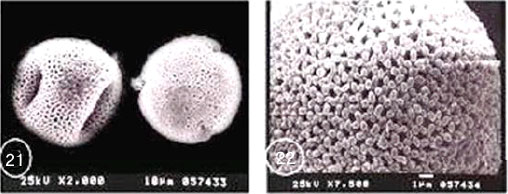 |
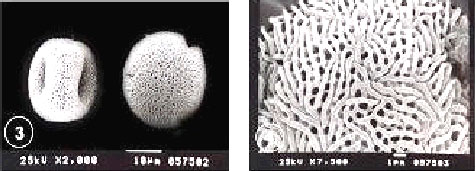
| Fig. 17-22:SEM micrographs of pollen morphology and exine sculpture of studied taxa of Geraniaceae |
| Fig. 17: Geranium dissectum general view |
| Fig. 18: G. dissectum exine pattern |
| Fig. 19: G. molle general view |
| Fig. 20: G. molle exine pattern |
| Fig. 21: G. trilophum, equatorial and polar view |
| Fig. 22: G. trilophum, exine pattern |
Key to the different pollen types and subtypes:
| 1 | a. Exine sculpture reticulate/gemmate...................................................................................................................type II |
| b. Exine sculpture not so............................................................................................................................................ 2 | |
| 2 | a. Exine sculpture striate to striatereticulate…......................................................................................................... type I |
| b. Exine sculpture reticulate (type III) ........................................................................................................................... 3 | |
| 3 | a. Muri articulate .......................................................................................................... Mansonia he liotropioides subtype |
| b. Muri not so ......................................................................................................................................................... 4 | |
| 4 | a.Lumina area 4.42 μm2…................................................................................................…. Monsonia senegalensis subtype |
| b. Lumina area 1.5 μm2 ................................................................................................. Pelargonium grandiflorum subtype |
 |
 |
 |
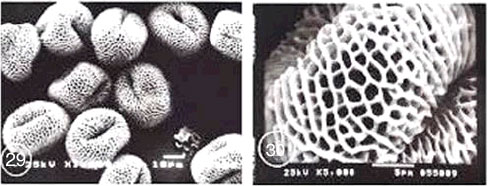 |
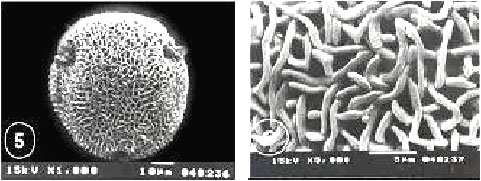
| Fig. 23-30: SEM micrographs of pollen morphology and exine sculpture of studied taxa of Geraniaceae |
| Fig. 23: Monsonia heliotropioides, polar view |
| Fig. 24: M. heliotropioides, exine pattern |
| Fig. 25: M. nivea, polar view |
| Fig. 26: M. nivea, exine pattern |
| Fig. 27: M. senegalensis, general view |
| Fig. 28: M. senegalensis, exine pattern |
| Fig. 29: Pelargonium grandiflorum, equatorial and polar view |
| Fig. 30: P. grandiflorum, exine pattern |
DISCUSSION
The variation of pollen described and documented above illustrates diverse pollen morphologies in Geraniaceae. These results were generally in accordance with those of Stafford and Blackmore (1991) as well as Perveen and Gaiser (1999), who described Geraniaceae as a eurypalynous family. Moreover the present study revealed that each genus has more or less its own characteristic pollen type. Based on the current study, three distinct pollen types in terms of surface ornamentations and exine structures were recognized viz., striate/reticulate, reticulate/gemmate and reticulate-type.
Pollen type-I (Striate/reticulate), is characterized by a striate reticulate tectum. All investigated Erodium species were found to fall into a general striate/reticulate ornamentation type, within which it was not possible to distinguish between the species. In the course of the current study, detailed SEM investigations have demonstrated a continuous range of sculptural variation occurs within pollen type I; thus, making it impossible to enable a subdivision on a non arbitrary basis. Bortenschlager (1967) carried out an extensive survey on the range of pollen types in the Gerniaceae. He examined the pollen morphology of 31 species of Erodium, of which 9 species were restudied in the course of this investigation. Bortenschlager recognized two basic pollen types, Geranium multiflorum and Erodium. According to the present results, pollen type I (Striate-Reticulate) is similar to the Erodium pollen type reported previously by Bortenschlager (1967). The latter distinguished three subtypes within his Erodium type based on the details of the pollen wall sculpture. The results of the present study, revealed no subtypes of pollen type I, in contrast to the findings of Bortenschlager (1967). On the other hand, the present results are in agreement with the finding of El-Oqlah (1983) who investigated the pollen morphology of 35 species of Erodium and pointed to the absence of pollen subtypes. Verhoeven and Venter (1987) reported a similar pollen type in Erodium species (except Erodium oxyrrhynchum ssp.) and more recently, Perveen and Gaiser (1999) studied the pollen morphology of 16 species of the family Geraniaceae from Pakistan and described the same pollen type in Erodium species.
Pollen type II (Reticulate/gemmate), is readily distinguished by the reticulate tectum which is heavily ornamented with bacula and gemmae. This pollen type is characteristic of Geranium species. The distinction between Geranium species included under this pollen type seemed to be difficult. Geranium trilophum in particular may be differentiated by larger size range 60 μm (57-62 μm) and oblate spheroid pollen. Otherwise the other two Geranium species were found to be quite similar to each other; both species showed the same pollen shape (prolate spheroid) and nearly the same size range (51-53 μm). Pollen type II is similar to the Geranium multiflorum type that described previously by Bortenschlager (1967) and also to the pollen grains of Geranium species described. On the other hand, the present result does not support El-Ghazaly (1991), who reported that the exine sculpture of Geranium molle is striato-reticulate while the present study showed a reticulate/gemmate exine sculpture for G. molle.
Pollen type III (Reticulate), is easily recognized by a coarsely reticulate tectum. Three species of Monsonia (Monsonia heliotropioides, M. nivea and M. senegalensis) beside Pelargonium grandiflorum were included under this pollen type. Venter (1979, 1983) as well as Verhoeven and Venter (1986) reported a similar type of pollen in several species of Monsonia. In addition, Stafford and Gibby (1992) in their extensive work in pollen morphology of Pelargonium recognized the same pollen type in many species of Pelargonium (as P. longifolium, P. luridum, P. patulum, P. pinnatum, P. crassipes and others). Pollen type III, was found to be variable both in the characters of exine sculpture, shape and size of pollen. These differences are sufficient for them to be categorized in separate subtypes: subtypeA (Monsonia heliotropioides), subtype B (Monsonia senegalensis) and subtype C (Pelargonium grandiflorum).
The first subtype A is characterized by heterobrochate reticulum, perforated nexine, articulate, fragmentimurate pollen and the presence of supratectal processes. This subtype is characteristic of Monsonia heliotropioides and M. nivea. The present work revealed that the distinction between M. heliotropioides and M. nivea palynologically is quite difficult as both species showed more or less similar pollen grains. This finding is in agreement with El-Ghazaly (1991), who pointed to the great similarity of their pollen grains. The second subtype B, with homomorphic reticulum, wide lumina and relatively straight and smooth muri was found to be characteristic to M. senegalensis. Venter (1979), on his monograph of Monsonia as well as El-Ghazaly (1991) who studied this species among others reported a similar type of pollen in the genus Monsonia. The species included under pollen subtypes A and B, could also be distinguished from each other not only by the variations in the pollen morphological characters, but also by the number of pollen content per anther. M. nivea and M. heliotropioides (pollen subtype A) showed a strikingly low number of pollen content comparing with that of M. senegalensis (pollen subtype B). A similar observation had been reported by Aldasoro et al. (2001); they studied the pollen morphology in combination with other morphological characters and gave an account on the phylogenetic relationships among 25 species of the old-world genus Monsonia.
The third pollen subtype C is distinguished by the reticulate exine sculpture, small lumina and laterally compressed undulating muri as well as prolate shape. This subtype is characteristic for Pelargonium grandiflorum. This type of pollen is similar to that described by Stafford and Gibby (1992).
CONCLUSION
The pollen morphological characters in the current study are considered to be diagnostic at the generic level and to some extent at the specific level. Moreover the present study confirmed the eurypalynous nature of the family. More studies are still needed in the future, utilizing more cosmopolitan material, to achieve more conclusive results.
REFERENCES
- Aedo, C., M. Garcia, A.L. Maria, J. Aldasoro and C. Navarro, 2007. Taxonomic revision of Geranium Subsect. Mediterranea (Geraniaceae). Syst. Bot., 32: 93-128.
Direct Link - Albers, F., 1991. Comparative karyological studies in Geraniaceae on family, genus and section level. Proceedings of the International Geraniaceae Symposium, September 24-26, 1990, University of Stellenbosch, Republic of South Africa, pp: 115-122.
Direct Link - Aldasoro, J.J., C. Navarro, P. Vargas and C. Aedo, 2001. Anatomy, morphology and cladistic analysis of Monsonia L. (Geraniaceae). Anales Jard. Bot. Madrid, 59: 75-100.
Direct Link - Aldasoro, J.J., C. Navarro, P. Vargas, L.L. Saez and C. Aedo, 2001. California, a new genus of Geraniaceae endemic to the Southwest of North America. Anales Jardin Botanico Madrid, 59: 209-216.
CrossRefDirect Link - Al-Quran, S.A., 2004. Pollen morphology of plantaginaceae in Jordan. Pak. J. Biol. Sci., 7: 1594-1602.
CrossRefDirect Link - El Oqlah, A.A., 1983. Pollen morphology of the genus Erodium l'Herit. in the Middle East Pollen Spores, 25: 383-394.
Direct Link - Erdtman, G., 1960. The acetolysis method: A revised description. Svenok. Bot. Tidskr, 54: 561-564.
Direct Link - Huysmans, S., S. Dessein, S. Erik and R. Elmar, 2003. Pollen morphology of NW European representatives confirms monophyly of Rubieae (Rubiaceae). Rev. Paleobot. Palynol., 127: 219-240.
CrossRefDirect Link - Link, D.A., 1991. The nectaries of geraniaceae. Proceeding of the International Geraniaceae Symposium, September 24-26, 1990, University of Stellenbosch, Republic of South Africa, pp: 215-225.
Direct Link - Mabberley, D.J., 1997. The Plant Book: A Portable Dictionary of Vascular Plants. 2nd Edn., Cambridge University Press, Cambridge, ISBN: 0521414210, pp: 858.
CrossRefDirect Link - Mulder, Ch., 2003. The Northwest European pollen flora, 61, Aizoaceae. Rev. Paleobot. Palynol., 123: 41-45.
Direct Link - Perveen, A. and M. Gaiser, 1999. Pollen flora of Pakistan-XV Geraniaceae. Turk. J. Bot., 23: 263-269.
Direct Link - Price, R.A. and J.D. Palmer, 1993. Phylogenetic relationships of the geraniaceae and geraniales from rbcL sequence comparisons. Ann. Mssouri Bot. Garden, 80: 661-671.
CrossRefDirect Link - Punt, W., P.P. Hoen, S. Blackmore, S. Nilsson and A. Le Thomas, 2007. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol., 143: 1-81.
CrossRefDirect Link - Stafford, P.J. and M. Gibby, 1992. Pollen morphology of the genus Pelargonium (Geraniaceae). Rev. Palaeobot. Palynol., 71: 79-109.
CrossRefDirect Link - Venter, H.J.T., 1979. A monograph of Monsonia L. (Geraniaceae): Meded Landbouhoogesch. Wageningen, 79: 1-128.
Direct Link - Venter, H.J., 1983. Phytogeography and interspecies relationships in Monsonia (Geraniaceae). Bothalia, 14: 865-869.
CrossRefDirect Link - Verhoeven, R.L. and H.J.T. Venter, 1987. Pollen morphology of Erodium in Southern Africa. S. Afr. J. Bot., 53: 279-283.
CrossRefDirect Link