
Badr E. El-Bialy
Department of Forensic Medicine and Toxicology, Egypt
Eman E. Abdeen
Department of Bacteriology, Mycology and Immunology, Faculty of Veterinary Medicine, University of Sadat City, Sadat City, Egypt
Nermeen B. El-Borai
Department of Forensic Medicine and Toxicology, Egypt
Eman M. El-Diasty
Animal Health Research Institute, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2016 | Volume: 19 | Issue: 1 | Page No.: 26-35







ABSTRACT
Aflatoxins (AFs), widely distributed food-borne mycotoxins, affect quality and safety of food and cause economic losses in livestock. In this study, the protective effect of Bee Pollen (BP) against some immunotoxic hazards elucidated from eating of AFs-containing diet was investigated in Wistar rats. Rats were randomly classified into four groups and treated for 30 days, Group 1; control negative, Group 2; Total AFs (3 mg kg–1 basal diet), Group 3; BP (20 g kg–1 basal diet) and Group 4; AFs+BP in basal diet. The immunoprotective effect of BP was revealed in terms of increasing (relative to levels seen in Group 2 rats that consumed the AFs diet) serum total protein and globulin levels, restored normal neutrophil (PMN)/lymphocyte ratio, increased PMN phagocytic activity and increased lymphocyte proliferative capacity. Also, the use of the BP reduced spleen H2O2 levels and increased GSH content while maintaining normal levels of NO formation. Histopathologic analysis showed that the AFs caused lymphocytic depletion in the spleen; however, BP induced lymphocytic hyperplasia and reduced the levels of AFs-inducible cellular exhaustion or depletion. These results provide evidence of a protective effect of BP against some immunotoxic actions induced in situ by consumption of AFs.
PDF Abstract XML References Citation
Received: October 21, 2015;
Accepted: November 25, 2015;
Published: December 15, 2015
How to cite this article
Badr E. El-Bialy, Eman E. Abdeen, Nermeen B. El-Borai and Eman M. El-Diasty, 2016. Experimental Studies on Some Immunotoxicological Aspects of Aflatoxins Containing Diet and Protective Effect of Bee Pollen Dietary Supplement. Pakistan Journal of Biological Sciences, 19: 26-35.
DOI: 10.3923/pjbs.2016.26.35
URL: https://scialert.net/abstract/?doi=pjbs.2016.26.35
DOI: 10.3923/pjbs.2016.26.35
URL: https://scialert.net/abstract/?doi=pjbs.2016.26.35
INTRODUCTION
Mycotoxin-producing fungi are significant invaders and destroyers of agricultural products and seeds in the field, during storage and processing, or in the markets (Jimoh and Kolapo, 2008). Mycotoxins, secondary metabolites produced by various fungi (Chandra and Sarbhoy, 1997; Maheshwar et al., 2009), contaminate foods and feedstuffs affecting their quality and safety and exert serious health hazard to poultry, animals and humans (Mokhles et al., 2007; Iheshiulor et al., 2011). Aflatoxins (AFs), widely distributed food-borne mycotoxins, are produced by a large number of Aspergillus species, mainly A. flavus and A. parasiticus. It is estimated that ≈25% of the food crops in the world are affected by mycotoxins. Beans, rice, corn, soybeans, wheat, cotton seed, barley and peanuts are the crops most often at risk of being contaminated with AFs (Caloni and Cortinovis, 2011; Reza et al., 2012).
Aflatoxins can also cause economic losses in livestock due to increases in disease susceptibility or decreases in productivity. Aflatoxicosis in farm animals is commonly under estimated due to a lack of specific early clinical signs. Humans and animals that consume AFs-contaminated feed eventually develop various health problems including growth retardation, hematologic disorders, hepato-, nephro-, geno- and immunotoxicity and even death (Wilson et al., 1997; Bintvihok, 2002; Gong et al., 2002; Fapohunda et al., 2008; Guindon et al., 2007). An increase in generation of Reactive Oxygen Species (ROS) and in lipid peroxidation (LPO) has been suggested as key underlying events associated with many of these toxic manifestations (Lee et al., 2005).
With regard to immunotoxicity, AFs can cause immunosuppression through depression of T-or B-lymphocyte activity (Reddy et al., 1987), Nitric Oxide (NO) and antibody productions, phagocytic activity as well as an impairment of several other major macrophage/neutrophil functions (Neldon-Ortiz and Qureshi, 1992; Moon et al., 1999), including modification of their ability to synthesize/release cytokines and perform phagocytic activities (Jakab et al., 1994; Marin et al., 2002).
Bee-Pollen (BP) is a natural product collected by honeybees from selected flower species, mixed with nectar and bee secretions (Le Blanc et al., 2009; Nakajima et al., 2009). The BP has been used for many years in both traditional medicine and as a dietary supplement mainly due to its nutritional properties and health benefits (Isla et al., 2001; Kroyer and Hegedus, 2001; Almeida-Muradian et al., 2005). While BP is replete with proteins, sterols, unsaturated fatty acids, vitamins, lipids, carbohydrates and minerals that have nutritional value (Campos et al., 2003), its health/therapeutic effects have been attributed mainly to its phenolic and flavonoids constituents (Le Blanc et al., 2009; Xu et al., 2009) that impart anti-oxidant (Marghitas et al., 2009), microbial (Garcia et al., 2001; Basim et al., 2006) and inflammatory effects (Akkol et al., 2010) and also seem to act as immunostimulants (De Oliveira et al., 2013; El-Asely et al., 2014).
Because of these latter effects, the present study was undertaken to determine if BP could impart protective efficacy against aspects of immunotoxic effect induced in situ by AFs exposure. To assess this, rat models were fed (for 30 days) a diet containing AFs in the presence/absence of BP and then a variety of immune system-related endpoints were evaluated.
MATERIALS AND METHODS
Animals: A total of 32 male Wistar rats (120-150 g) were purchased from Al-Zyade Experimental Animals Production Center (Giza, Egypt). All animals were kept in 50×30×30 polypropylene cages (four animal in each cage) in ventilated pathogen free rooms maintained at 22-25°C with a 60% relative humidity and a 12 h light: dark cycle. All rats had ad libitum access to rodent chow purchased from Al-Zyade Experimental Animals Production Center and filtered tap water. The rats were quarantined for 2 week prior to the start of experiments to acclimatize. The Research Ethical Committee of the Faculty of Veterinary Medicine, University of Sadat City, (Sadat City, Egypt) approved all aspects of the experimental design here.
Aflatoxins: Aspergillus flavus isolated from broiler feed was obtained from the Mycology Department of the Animal Health Research Institute, Giza, Egypt and confirmed by sub-culture onto Malt Extract and Czapek Dox agar and incubation for 8 day at 25°C. The A. flavus was screened for aflatoxins (AFs) production using Thin Layer Chromatography (TLC) (Munimbazi and Bullerman, 1998). Presence of various AFs was qualitatively confirmed by the appearance of blue fluorescence on the plate and comparison of the spot’s Retention Factor (RF) value versus the RF of a known standard.
Crushed yellow corn, obtained commercially from Al-Ahram Company poultry rations (Giza, Egypt) was verified as completely free from fungal or mycotoxins contaminations by gross and TLC examination. The corn was placed in flasks and then autoclaved at 121°C for 15 min on three successive days. The corn was then treated with 10 mL spore suspension (at 107 spores/mL) and the whole mixture fermented by incubation at 28-30°C for 21 days. After incubation, the flask was dried in a 60°C oven for 24 h to kill the fungus and then the product was powdered using a grinder. A representative 25 g sample of the yield was assessed for AFs content using AOAC official methods (AOAC., 1980). Routinely, the corn produce was found to bear ≈150 mg total AFs/kg corn. This corn was then incorporated into basal diet at a level that would provide a desired dose of 3 mg of total AFs/kg diet.
Reagents: Bee Pollen (BP) was purchased from an apiary in Sadat City, Egypt. Kits to measure serum total protein and albumin, as well as levels of reduced glutathione (GSH), hydrogen peroxide (H2O2) and Nitric Oxide (NO) in spleen homogenates, were purchased from Biodiag-nostic Company (Dokki, Egypt). All the other chemicals, biological agents, tissue culture media and reagents were purchased from Oxoid Company (United Kingdom) and Sigma (Cairo).
Treatments and rat sample collection: For the studies, after the acclimation period, the rats were randomly allocated into four groups (n = 8/group). Group 1: Rats (control) to be fed basal diet. Group 2: Rats to be fed basal diet containing AFs at 3 mg total AFs/kg diet based on Abdel-Wahhab et al. (2007). Group 3: Rats to be fed basal diet containing 2% (20 g kg–1 diet) Bee Pollen (BP) based on Khalil and El-Shiekh (2010). Group 4: Rats to be fed basal diet containing AFs and BP. All rats were fed the various diets for 30 days.
At the completion of the feeding period, all rats were fasted overnight and blood samples were collected from retro-orbital veinus plexus under light ether anesthesia and then euthanized to permit tissue samples collection. Some samples of blood were collected into heparinized tubes for later use in evaluation of phagocytic activity and lymphocyte proliferation. Other samples were collected into EDTA coated tubes for use in differential leukocyte counts and calculation of blood neutrophil/lymphocyte ratios. Additional samples were collected into tubes without anti-coagulant and centrifuged at 3000 rpm for 15 min to isolate serum for use in biochemical analysis. Specimens of spleen from each rat were then collected and stored at -20°C for later biochemical investigations or placed in 10% formalin for histopathology.
Assessment of BP protective effects against AFs-induced immunotoxicity: Serum samples were analyzed for total protein and albumin using commercial kits. From these values, serum globulin level was calculated using Globulin = Total protein-Albumin (in g dL–1). Differential counts were performed using routine hematological procedures (Feldman et al., 2000) to permit estimation of neutrophil/lymphocyte ratios. For the analysis of GSH, H2O2 and NO contents in splenic tissues, samples of the isolated organ were homogenized using a Dounce Tissue Grinder (Omni International, Kennesaw Georgia) (Tissue homogenate was prepared by rinsing the tissue in Phosphate Buffered Saline (PBS) of pH 7.4 containing 0.16 mg mL–1 heparin to remove any red blood cells and clots then 1 g tissue was homogenized in 5-10 mL cold PBS, pH 7.4 in the grinder that surrounded by ice, after that the samples were centrifuged at 4°C, 4000 rpm for 15 min). Thereafter, the samples were analyzed for their GSH, H2O2 and NO levels using commercial kits with following their instructions.
Phagocytic activity: Phagocytosis by polymorphonuclear cells (neutrophils; PMN) was assessed by using heat-killed Candida albicans, according to the method described by Wilkinson (1981). In brief, 100 μL C. albicans (106/mL), 100 μL rat serum (isolated from naïve rats) and 100 μL (heparinized) blood from a given rat were combined and incubated at 37°C for 30 min. The mixture was then centrifuged at 1,000 rpm for 5 min and the resulting supernatant then discarded. Smears of the sedimented materials were then prepared and stained with Giemsa stain. Using a light microscope, the total number of PMN that engulfed C. albicans per the total number of PMN (100 PMN) examined in a given slide field was determined and the results expressed as percentage with Phagocytic Activity (PA). Among the PMN that did ingest yeast, total individual organisms were counted; from this data, the Phagocytic Index (PI) (i.e., average total number candida ingested/phagocytically-active cells) was determined. In all cases, a minimum of two slides/rat were evaluated.
Lymphocyte proliferation: Heparinized rat blood samples were diluted 1:1 using HBSS (Hank's balanced salt solution) in sterile tubes. The separation of lymphocytes was done by layering of blood in Ficol (2:1) and centrifuged at 400×g at 4°C for 30 min to give packed blood cells with granulocyte, interface layer (which contain lymphocytes) and upper plasma layer. The interface layer was carefully aspirated using sterile glass Pasteur pipette, then placed in sterile tubes containing 2 mL RPMI 1640 medium. Cells were washed 3 times with RPMI 1640 medium by centrifugation at 400×g for 10 min at 4°C. After the last wash, the sediment lymphocytes were suspended in 1 mL of RPMI 1640 medium without Fetal Calf Serum (FCS). The RBCs contamination, if any, was removed by the distilled water lysis method. Lymphocyte numbers were then calculated using a hemocytometer; cells were then placed into wells of a 96 well plate (at 106 cells/150 μL culture medium [RPMI 1640+10% FBS]). Proliferative responses were then estimated using triplicate sets of cells stimulated with phytohemagglutinin (15 μg PHA/mL, final concentration in well), control cells (spontaneous proliferation) would be assessed in cells that received medium only. All cells were then cultured at 37°C (in a 5% CO2 incubator) for 72 h before 10 μL MTT (3[4,5-dimethylthiazol-2-y1]-2,5-diphenyltetrazolium bromide; 5 mg mL–1) was added to each well. The cells were incubated a further 4 h at 37°C before 95% DMSO was added to each well (100 μL/well) to dissolve the formazan that formed in viable reproducing cells. The absorbance (OD) in each well was measured at 570 nm in a Versa Max microplate reader (Molecular Devices, Sunnyvale, CA) and relative proliferation determined by comparing the ODtreated rat vs. ODcontrol rat (Rai-el-Balhaa et al., 1985).
Histopathology: Spleens collected at necropsy were fixed overnight in 10% neutral formalin and then embedded in paraffin. Sections (5 μm thick) were then prepared and stained with hematoxylin and eosin (Bancroft et al., 1996). The samples were then examined using a light microscope to characterize any damage in the samples.
Statistics: Values are presented as Mean±Standard Error (SE). All statistical analyses were performed using SPSS (Statistical package for Social Sciences) Version 16 released on 2007, the significance between AFs-group with control negative group and AFs+BP-treated group with AFs group was considered at p<0.05.
RESULTS
Serum total protein level: The present study revealed that the consumption of aflatoxins (AFs; Group 2) daily in the diet for 30 days resulted in significant reduction in total protein in the serum of rats versus (vs) values in control rats consuming the basal diet only (Group 1) (Table 1). The AFs exposure had no significant effect on levels of globulin or albumin, though levels of globulin were reduced by ≈ 18%. Consumption of bee pollen (BP; Group 3 rats) alone as a dietary supplement had no significant impact on any of these parameters. When consumed along with the AFs, the BP enabled the rats to maintain normal total protein levels, have nearly normal total globulin levels (i.e., slightly lower vs. Group 1 values) and actually have significantly elevated (even vs. control) total albumin levels in their blood compared to values seen in the rats fed diet containing the AFs only.
| Table 1: | Serum total protein, albumin and globulin levels in experimental rats |
 | |
Values shown are Mean±SE (n = 6 animals/group),aValue significantly different from Group 1 at p<0.05,bValue significantly different from Group 2 at p<0.05, Group 1: Control, Group 2: AF in diet, Group 3: BP in diet, Group 4: AF+BP | |
| Table 2: | Splenic GSH, NO and H2O2 levels in treated rats |
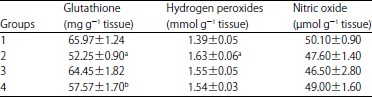 | |
Values shown are means±SE (n = 6 animals/group),aValue significantly different from Group 1 at p<0.05,bValue significantly different from Group 2 at p<0.05, Group 1: Control, Group 2: AF in diet, Group 3: BP in diet, Group 4: AF+BP | |
| Table 3: | Neutrophil, lymphocyte and neutrophil/lymphocyte ratios in experimental rats’ blood |
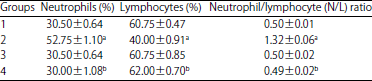 | |
Values shown are Mean±SE (n = 6 animals/group), aValue significantly different from Group 1 at p<0.05, bValue significantly different from Group 2 at p<0.05, Group 1: Control, Group 2: AF in diet, Group 3: BP in diet, Group 4: AF+BP | |
Levels of reduced glutathione, hydrogen peroxide and nitric oxide in spleen tissue: Analysis of spleen homogenates showed that consumption of the AFs induced significant reductions in reduced glutathione (GSH) contents and elevations in hydrogen peroxide (H2O2) levels compared to values in samples from Group 1 control rats (Table 2). The AFs caused a non-significant decrease in splenic nitric oxide (NO) levels. Co-consumption of BP with AFs (Group 4 rats) appeared to mitigate the effects of AFs on splenic GSH (significant increase) and H2O2 levels in relation to aflatoxicated rats.
Neutrophils, lymphocytes percentages and neutrophils lymphocytes ratio N/L: The assessments of PMN and lymphocytes levels in the blood of the various treated rats (Table 3) revealed that exposure to AFs itself resulted in (relative to in Group 1 rats) significant neutrophilia and lymphocytopenia, with an accordingly-increased N/L ratio. Consumption of BP alone as a dietary supplement had no significant effect on any of these parameters. Co-consumption of BP with AFs appeared to mitigate the effects of AFs on each of these endpoints and induced significant elevation in neutrophils, lymphocytes percentages and neutrophils lymphocytes ratios N/L.
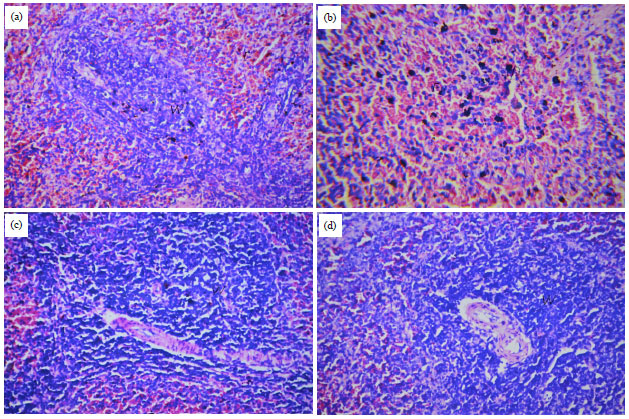 | |
| Fig. 1(a-d): | (a) Normal red and white pulps of spleen of control rats, (b) Lymphoid depletion in the white pulps with congestion and pigmented material in the red pulps, (c) Lymphoid hyperplasia in white pulps of rats treated with BP group and (d) Lymphoid hyperplasia in white pulps of rats treated with AFs and BP (H and E, X40) |
| Table 4: | Phagocytic activity/index and lymphocyte proliferation by cells from treated rats |
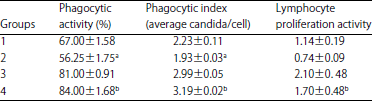 | |
Values shown are Mean±SE (n = 6 animals/group), aValue significantly different from Group 1 at p<0.05, bValue significantly different from Group 2 at p<0.05, Group 1: Control, Group 2: AF in diet, Group 3: BP in diet, Group 4: AF+BP | |
Phagocytic activity/index and inducible lymphocyte proliferation: From a functional standpoint, the circulating PMN and lymphocytes of rats fed a diet containing AFs had significant decreases in, respectively, phagocytic activity/index and inducible lymphocyte proliferation ex vivo compared with corresponding cells isolated from control rats (Table 4). Interestingly, in these sets of evaluations, it appeared that consumption of BP alone as a dietary supplement or as a co-exposure agent had a significant effect on each of these parameters. In each case, the measured values were well above those for cells from the Group 1 (control) or Group 2 (AFs-exposed rats).
Spleen histopathology: Histopathological examination of spleen (Fig. 1) indicated that compared to the organ from control rats (Fig. 1a), consumption of the AFs-bearing diet resulted in lymphoid depletion in the white pulp and congestion and increases in pigmented materials present in the red pulp (Fig. 1b). In rats that consumed the BP as just a dietary supplement, there were indications of lymphoid hyperplasia in the white pulp (Fig. 1c). In the spleens of rats that co-consumed AFs and BP, there was also clear evidence of the hyperplasia, but not as much lymphoid depletion or damage to the red pulp as in the tissues of the Group 2 rats (Fig. 1d).
DISCUSSION
Fungi are significant destroyers of foodstuffs and grains during storage, rendering them unfit for human consumption by retarding their nutritive value and often by producing mycotoxins (Satish et al., 2007).
Nutritional stress factors have a negative impact on the oxidant/anti-oxidant balance that contributes to regulation of various metabolic pathways essential for maintenance of immuno-competence. Mycotoxins, including several AFs, are considered to be among the most important feed-borne stress factors leading to disturbance of the oxidant/anti-oxidant balance and cause immunosuppression (Silvotti et al., 1997).
As noted above, mycotoxins elicit a wide spectrum of toxicological effects. Of particular interest is the capacity of some mycotoxins to alter normal immune function when present in food at levels below those that induced overt toxicities (Surai and Dvorska, 2005). The immune system is primarily responsible for defense against invading organisms. Immune response suppression by AFs eventually could impair immune function in developing animals (Silvotti et al., 1997), impaired host resistance to infection (Jiang et al., 2005) and induced re-activation of chronic infections (Venturini et al., 1996; Kubena et al., 2001).
Aflatoxins inhibit both humeral and cell mediated immunity. Aflatoxin Bl inhibits protein synthesis and cell proliferation (Sharma, 1993). Since immunoglobulins are protein in nature, a decrease in total protein and globulin might result in reduced antibody production which ultimately results in decreased immunity.
Aflatoxins have been shown to cause lowering of serum total protein levels in rats (El-Nekeety et al., 2014), rabbits (Yousef et al., 2003) and broilers (Raju and Devegowda, 2000). It is quite likely the decreased synthesis/secretion of proteins by cells might be related to a formation of AFs adducts with DNA, RNA and organelle proteins, all outcomes that potentially inhibit normal gene transcription/translation processes in cells (Wang and Groopman, 1999). Thus, the results here in the AFs-exposed rats were in keeping with previous reports in the literature. With regard to BP, previous studies showed that BP administration led to elevations in serum total protein levels (Khalil and El-Shiekh, 2010; Attia et al., 2014; Capcarova et al., 2013). These increases were thought likely be due to the contributions from protein and amino acids constituents of the BP (Campos et al., 2003), thus the use of the BP was able to protect the rats from expected AFs-induced reductions in these values.
The present study also revealed there was a significant neutrophilia and lymphocytopenia (with increasing N/L ratios) in AFs-intoxicated rats. Several studies have demonstrated that AFs can induce lymphocytopenia, monocytopenia and heterophilia in chickens (Basmacioglu et al., 2005; Samuel et al., 2009) and in rats and mice (Abdel-Wahhab et al., 2002; Tuzcu et al., 2010; Gupta et al., 2011). The neutrophilia could be attributed to a general inflammation and alterations in bone marrow cellularity/function in response to the AFs (Abdel-Wahhab and Aly, 2003; Hassan et al., 2012). Pier and Mcloughlin (1985), Oguz et al. (2000), Mehrzad et al. (2011) and Sahar et al. (2011) previously reported that AFs could cause suppressed phagocytosis by macrophages. These authors also indicated that the toxins cause thymic aplasia and suppress cell-mediated immunity and leukocyte migration. Thus, the present findings of decreased phagocytic activity/indices in the PMN of AFs-exposed rats would be in keeping with those studies. With regard to BP, the current results were in agreement with those of Attia et al. (2014), who showed that consumption of BP significantly increased the phagocytic capacity and index in granulocytes of rabbits. Moreover, Li et al. (2009) indicated that BP was a potential immuno-stimulant in that BP consumption led to significant enhancement of phagocytic capacity and index values for peritoneal macrophages in rats. Indeed, in the current study, the PMN from rats given the BP for 30 days (with or without AFs) had significant increases in phagocytic activity and indices above those in cells of control rats and of AFs-intoxicated hosts.
In vitro/ex vivo lymphocyte proliferation is often used to assess potential immuno-suppressive effects of toxicants (Hayek et al., 1996). The findings here of AFs-induced reductions in ex vivo lymphocyte proliferation are in agreement with those of Meissonnier et al. (2008), who reported inhibition of lymphocyte proliferation by cells isolated from AFs-intoxicated pigs. The current results are also in line with the findings of AFB1-induced decreases in lymphocyte proliferation and cytokine production by cells isolated from exposed experimental animals (Abbes et al., 2010). As with the PMN-related functional parameters, the consumption of BP here led to strong increases in ex vivo lymphocyte proliferation. While these outcomes could simply be related to the increased presence of amino acids, vitamins and trace elements that stimulate proliferation and differentiation of immune cells (Akter et al., 2006), it is also possible that polysaccharide constituents in the BP act to stimulate T-lymphocyte formation (Wang et al., 2005) and this carried over to the post-harvest protocols.
Within the spleen, there are several immune (and non-immune cell types). However, the ability to generate NO and H2O2, is relegated to the immune cells present. Unfortunately, the same cannot be said with regard to GSH, as erythrocytes and other local cells have substantial GSH contents (Rossi et al., 2009; Van Zwieten et al., 2014). The observed reductions in NO levels in the spleens of rats fed the AFs-only-bearing diet could be related to changes in protein synthesis in the hosts, there could be less inducible Nitric Oxide Synthase (iNOS) produced within the cells. It is also possible that the observed reductions in NO levels may have evolved secondary to inhibition of iNOS itself, as was previously documented by Cheng et al. (2002) and Chatterjee and Ghosh (2012). The small improvements over splenic NO levels in the rats fed BP alone or BP along with AFs in the diet could of course arisen not as a result of changes in the presence/activity of iNOS but from effects on NO by phenolics and flavonoids present in the BP (Leja et al., 2007; Akkol et al., 2010; Maruyama et al., 2010). These classes of agents have an anti-oxidant effect that has been attributed to their ability to scavenge free radicals or by acting as reducing agents/hydrogen donors (Caldwell, 2003; Capcarova et al., 2013).
This effect can also now help to explain the findings here whereby co-consumption of the BP prevented the significant reductions in splenic GSH and elevations in splenic H2O2 levels seen with the AFs alone. Moita et al. (2013) showed that BP could act as an anti-inflammatory agent by helping to scavenge endogenous ROS. As anti-oxidants, phenolic and flavonoids contents (Le Blanc et al., 2009; Xu et al., 2009) in the BP could serve as ROS scavengers (Abdella et al., 2009). As a consequence of induced H2O2 formation, it would not be unexpected that there would be peroxidative damage to cells in the organs of the exposed rats. Indeed, because reduced GSH is critical to mitigating/repairing peroxidative damage to membranes on affected cells, it was understandable that the spleens of rats fed the AFs alone-bearing diet had significantly reduced levels of this tripeptide. Therefore, by reducing the levels of ROS, the BP by this activity alone could have led to less peroxidative damage in the rats’ tissues and so prevented the sharp decreases expected from co-consumption of the AFs. Interestingly, there is also an alternative explanation for the observed effects on GSH levels in the spleens. Specifically, Myhrstad et al. (2002) showed that flavonoids could increase the expression of the rate-limiting enzyme in the synthesis of GSH, as c-glutamylcysteine synthetase. If this was to also be happening in the rats co-fed BP with AFs, this would allow for a secondary means of preventing loss of GSH in these hosts (secondary to fact these hosts also have their protein synthesizing capacities augmented above those in rats only exposed to the Afs).
CONCLUSION
Collectively, the current data support out contention that bee pollen may be a promising dietary source of anti-oxidants and could potentially impart immuno-protective effects during aflatoxicosis.
ACKNOWLEDGMENTS
The authors would like to thank all the workers and laboratory staff for their cooperation and efforts to complete this study.
REFERENCES
- Abbes, S., J.B. Salah-Abbes, M.A. Abdel-Wahhab and R. Ouslati, 2010. Immunotoxicological and biochemical effects of aflatoxins in rats prevented by tunisian montmorillonite with reference to HSCAS. Immunopharmacol. Immunotoxicol., 32: 514-522.
CrossRefDirect Link - Abdella, E.M., A. Tohamy and R.R. Ahmad, 2009. Antimutagenic activity of Egyptian propolis and bee pollen water extracts against cisplatin-induced chromosomal abnormalities in bone marrow cells of mice. Iran. J. Cancer Prev., 2: 175-181.
Direct Link - Abdel-Wahhab, M.A. and S.E. Aly, 2003. Antioxidants and radical scavenging properties of vegetable extracts in rats fed aflatoxin-contaminated diet. J. Agric. Food Chem., 51: 2409-2414.
CrossRefDirect Link - Abdel-Wahhab, M.A., S.A. Nada and F.A. Khalil, 2002. Physiological and toxicological responses in rats fed aflatoxin-contaminated diet with or without sorbent materials. Anim. Feed Sci. Technol., 97: 209-219.
Direct Link - Abdel-Wahhab, M.A., E.A. Omara, M.M. Abdel-Galil, N.S. Hassan, S.A. Nada, A. Saeed and M.M. El-Sayed, 2007. Zizyphus spina-christi extract protects against aflatoxin B1-initiated hepatic carcinogenicity. Afr. J. Tradit. Complement. Altern. Med., 4: 248-256.
PubMedDirect Link - Sahar, T.A., A.H. Abeer, A.H. Manal and K.M. Mogda, 2011. Influence of rosemary extract on immune responses and oxidative stress in mice intoxicated by aflatoxins. Nat. Sci., 9: 54-63.
Direct Link - Akkol, E.K., D.D. Orhan, I. Gurbuz and E. Yesilada, 2010. In vivo activity assessment of a honey-bee pollen mix formulation. Pharm. Biol., 48: 253-259.
CrossRefPubMedDirect Link - Akter, S., M.Z.I. Khan, M.R. Jahan, M.R. Karim and M.R. Islam, 2006. Histomorphological study of the lymphoid tissues of broiler chickens. Bangl. J. Vet. Med., 4: 87-92.
Direct Link - Almeida-Muradian, L.B., L.C. Pamplona, S. Coimbra and O.M. Barth, 2005. Chemical composition and botanical evaluation of dried bee pollen pellets. J. Food Compos. Anal., 18: 105-111.
CrossRefDirect Link - Attia, Y.A., A.M. El-Hanoun, F. Bovera, G. Monastra, W.S. El-Tahawy and H.I. Habiba, 2014. Growth performance, carcass quality, biochemical and haematological traits and immune response of growing rabbits as affected by different growth promoters. J. Anim. Physiol. Anim. Nutr., 98: 128-139.
CrossRefPubMedDirect Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Basim, E., H. Basim and M. Ozcan, 2006. Antibacterial activities of Turkish pollen and propolis extracts against plant bacterial pathogens. J. Food Eng., 77: 992-996.
CrossRefDirect Link - Basmacioglu, H., H. Oguz, M. Ergul, R. Col and Y.O. Birdane, 2005. Effect of dietary esterified glucomannan on performance, serum biochemistry and haematology in broilers exposed to aflatoxin. Czech J. Anim. Sci., 50: 31-39.
Direct Link - Bintvihok, A., 2002. New insights to controlling mycotoxin danger in ducks. Feed Technol., 6: 28-29.
Direct Link - Caldwell, C.R., 2003. Alkylperoxyl radical scavenging activity of red leaf lettuce (Lactuca sativa L.) phenolics. J. Agric. Food Chem., 51: 4589-4595.
CrossRefPubMedDirect Link - Caloni, F. and C. Cortinovis, 2011. Toxicological effects of aflatoxins in horses. Vet. J., 188: 270-273.
CrossRefDirect Link - Campos, M.G., R.F. Webby, K.R. Markham, K.A. Mitchell and A.P. da Cunha, 2003. Age-induced diminution of free radical scavenging capacity in bee pollens and the contribution of constituent flavonoids. J. Agric. Food Chem., 51: 742-745.
CrossRefPubMedDirect Link - Capcarova, M., A. Kolesarova, A. Kalafova, B. Galik, M. Simko, M. Juracek and R. Toman, 2013. The role of dietary bee pollen in antioxidant potential in rats. Eur. J. Vet. Sci., 29: 133-137.
Direct Link - Chandra, R. and A.K. Sarbhoy, 1997. Production of aflatoxins and zearalenone by the toxigenic fungal isolates obtained from stored food grains of commercial crops. Indian Phytopathol., 50: 458-468.
Direct Link - Chatterjee, D. and P. Ghosh, 2012. Sub-cytotoxic concentration of AflatoxinB2 prevents NO-mediated increased mitochondrial membrane potential and intracellular killing of Candida albicans in macrophages. Adv. Life Sci., 2: 52-56.
CrossRefDirect Link - Cheng, Y.H., T.F. Shen and B.J. Chen, 2002. Induction of changes in morphology, reactive nitrogen/oxygen intermediates and apoptosis of duck macrophages by aflatoxin B1. Asian-Aust. J. Anim. Sci., 15: 1639-1645.
Direct Link - El-Asely, A.M., A.A. Abbass and B. Austin, 2014. Honey bee pollen improves growth, immunity and protection of Nile tilapia (Oreochromis niloticus) against infection with Aeromonas hydrophila. Fish Shellfish Immunol., 40: 500-506.
CrossRefDirect Link - El-Nekeety, A.A., S.H. Abdel-Azeim, A.M. Hassan, N.S. Hassan, S.E. Aly and M.A. Abdel-Wahhab, 2014. Quercetin inhibits the cytotoxicity and oxidative stress in liver of rats fed aflatoxin-contaminated diet. Toxicol. Rep., 1: 319-329.
CrossRefDirect Link - Fapohunda, S.O., C.N. Ezekiel, O.A. Alabi, A. Omole and S.O. Chioma, 2008. Aflatoxin-mediated sperm and blood cell abnormalities in mice fed with contaminated corn. Mycobiology, 36: 255-259.
CrossRefPubMedDirect Link - Li, F., Q. Yuan and F. Rashid, 2009. Isolation, purification and immunobiological activity of a new water-soluble bee pollen polysaccharide from Crataegus pinnatifida Bge. Carbohydr. Polym., 78: 80-88.
CrossRefDirect Link - Garcia, M., C. Peres-Aquillue, T. Juan, M.I. Juan and A. Herrera, 2001. Pollen analysis and antibacterial activity of spanish honeys. Food Sci. Technol. Int., 7: 155-158.
CrossRefDirect Link - Guindon, K.A., L.L. Bedard and T.E. Massey, 2007. Elevation of 8-hydroxydeoxyguanosine in DNA from isolated mouse lung cells following In vivo treatment with aflatoxin B1. Toxicol. Sci., 98: 57-62.
CrossRefDirect Link - Gupta, R., V. Sharma and S. Sharma, 2011. Chemopreventive potential of tinospora cordifolia root extract against aflatoxin b1-induced toxicity in swiss albino mice. Int. J. Biol. Med. Res., 2: 1115-1121.
Direct Link - Hassan, Z.U., M.Z. Khan, A. Khan, I. Javed and Z. Hussain, 2012. Effects of individual and combined administration of ochratoxin A and aflatoxin B1 in tissues and eggs of White Leghorn breeder hens. J. Sci. Food Agric., 92: 1540-1544.
CrossRefDirect Link - Iheshiulor, O.O.M., B.O. Esonu, O.K. Chuwuka, A.A. Omede, I.C. Okoli and I.P. Ogbuewu, 2011. Effects of mycotoxins in animal nutrition: A review. Asian J. Anim. Sci., 5: 19-33.
CrossRefDirect Link - Isla, M.I., M.I.N. Moreno, A.R. Sampietro and M.A. Vattuone, 2001. Antioxidant activity of Argentine propolis extracts. J. Ethnopharmacol., 76: 165-170.
CrossRefDirect Link - Jakab, G.J., R.R. Hmieleski, A. Zarba, D.R. Hemenway and J.D. Groopman, 1994. Respiratory aflatoxicosis: Suppression of pulmonary and systemic host defenses in rats and mice. Toxicol. Applied Pharmacol., 125: 198-205.
CrossRefDirect Link - Jiang, Y., P.E. Jolly, W.O. Ellis, J.S. Wang, T.D. Phillips and J.H. Williams, 2005. Aflatoxin B1 albumin adduct levels and cellular immune status in Ghanaians. Int. Immunol., 17: 807-814.
CrossRefPubMedDirect Link - Jimoh, K.O. and A.L. Kolapo, 2008. Mycoflora and aflatoxin production in market samples of some selected Nigerian foodstuffs. Res. J. Microbiol., 3: 169-174.
CrossRefDirect Link - Khalil, F.A. and N.M. El-Sheikh, 2010. The effects of dietary Egyptian propolis and bee pollen supplementation against toxicity if sodium fluoride in rats. J. Am. Sci., 6: 310-316.
Direct Link - Kroyer, G. and N. Hegedus, 2001. Evaluation of bioactive properties of pollen extracts as functional dietary food supplement. Innov. Food Sci. Emerg. Technol., 2: 171-174.
CrossRefDirect Link - Kubena, L.F., R.H. Bailey, J.A. Byrd, C.R. Young, D.E. Corrier, L.H. Stanker and G.E. Rottinghaust, 2001. Cecal volatile fatty acids and broiler chick susceptibility to Salmonella typhimurium colonization as affected by aflatoxins and T-2 toxin. Poult. Sci., 80: 411-417.
CrossRefDirect Link - Le Blanc, B.W., O.K. Davis, S. Boue, A DeLucca and T. Deeby, 2009. Antioxidant activity of Sonoran desert bee pollen. Food Chem., 115: 1299-1305.
CrossRefDirect Link - Lee, J.K., E.H. Choi, K.G. Lee and H.S. Chun, 2005. Alleviation of aflatoxin B1-induced oxidative stress in HepG2 cells by volatile extract from Allii Fistulosi Bulbus. Life Sci., 77: 2896-2910.
CrossRefDirect Link - Leja, M., A. Mareczek, G. Wyzgolik, J. Klepacz-Baniak and K. Czekonska, 2007. Antioxidative properties of bee pollen in selected plant species. Food Chem., 100: 237-240.
CrossRefDirect Link - Marghitas, L.A., O.G. Stanciu, D.S. Dezmirean, O. Bobis, O. Popescu, S. Bogdanov and M.G. Campos, 2009. In vitro antioxidant capacity of honeybee-collected pollen of selected floral origin harvested from Romania. Food Chem., 115: 878-883.
CrossRefDirect Link - Marin, D.E., I. Taranu, R.P. Bunaciu, F. Pascale and D.S. Tudor et al., 2002. Changes in performance, blood parameters, humoral and cellular immune responses in weanling piglets exposed to low doses of aflatoxin. J. Anim. Sci., 80: 1250-1257.
Direct Link - Maruyama, H., T. Sakamoto, Y. Araki and H. Hara, 2010. Anti-inflammatory effect of bee pollen ethanol extract from Cistus sp. of Spanish on carrageenan-induced rat hind paw edema. BMC Complement. Altern. Med., Vol. 10.
CrossRef - Maheshwar, P.K., S.A. Moharram and G.R. Janardhana, 2009. Detection of fumonisin producing Fusarium verticillioides in paddy (Oryza sativa L.) using Polymerase Chain Reaction (PCR). Braz. J. Microbiol., 40: 134-138.
CrossRefDirect Link - Mehrzad, J., G. Klein, J. Kamphues, P. Wolf, N. Grabowski and H.J. Schuberth, 2011. In vitro effects of very low levels of aflatoxin B1 on free radicals production and bactericidal activity of bovine blood neutrophils. Vet. Immunol. Immunopathol., 141: 16-25.
CrossRefDirect Link - Meissonnier, G.M., P. Pinton, J. Laffitte, A.M. Cossalter and Y.Y. Gong et al., 2008. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Applied Pharmacol., 231: 142-149.
CrossRefPubMedDirect Link - Moita, E., A. Gil-Izquierdo, C. Sousa, F. Ferreres and L.R. Silva et al., 2013. Integrated analysis of COX-2 and iNOS derived inflammatory mediators in LPS-stimulated RAW macrophages pre-exposed to Echium plantagineum L. bee pollen extract. PloS One, Vol. 8.
CrossRef - Mokhles, M., M.A. Abd El Wahhab, M. Tawfik, W. Ezzat, K. Gamil and M. Ibrahim, 2007. Detection of aflatoxin among hepatocellular carcinoma patients in Egypt. Pak. J. Biol. Sci., 10: 1422-1429.
CrossRefPubMedDirect Link - Moon, E.Y., D.K. Rhee and S. Pyo, 1999. Inhibition of various functions in murine peritoneal macrophages by aflatoxin B1 exposure in vivo. Int. J. Immunopharmacol., 21: 47-58.
CrossRefDirect Link - Munimbazi, C. and L.B. Bullerman, 1998. Isolation and partial characterization of antifungal metabolites of Bacillus pumilus. J. Applied Microbiol., 84: 959-968.
CrossRefPubMedDirect Link - Myhrstad, M.C.W., H. Carlsen, O. Nordstrom, R. Blomhoff and J.O. Moskaug, 2002. Flavonoids increase the intracellular glutathione level by transactivation of the γ-glutamylcysteine synthetase catalytical subunit promoter. Free Rad. Biol. Med., 32: 386-393.
CrossRefPubMedDirect Link - Neldon-Ortiz, D.L. and M.A. Qureshi, 1992. The effects of direct and microsomal activated aflatoxin B1 on chicken peritoneal macrophages in vitro. Vet. Immunol. Immunopathol., 31: 61-76.
CrossRefDirect Link - Oguz, H., T. Kececi, Y.O. Birdane, F. Onder and V. Kurtoglu, 2000. Effect of clinoptilolite on serum biochemical and haematological characters of broiler chickens during aflatoxicosis. Res. Vet. Sci., 69: 89-93.
CrossRefDirect Link - De Oliveira, M.C., D.M. da Silva, F.C. Loch, P.C. Martins, D.M.B. Dias and G.A. Simon, 2013. Effect of bee pollen on the immunity and tibia characteristics in broilers. Revista Brasileira de Ciencia Avicola, 15: 323-327.
CrossRefDirect Link - Rai-el-Balhaa, G., J.L. Pellerin, G. Bodin, A. Abdullah and H. Hiron, 1985. Lymphoblastic transformation assay of sheep peripheral blood lymphocytes: A new rapid and easy-to-read technique. Comp. Immunol. Microbiol. Infect. Dis., 8: 311-318.
CrossRefDirect Link - Raju, M.V. and G. Devegowda, 2000. Influence of esterified-glucomannan on performance and organ morphology, serum biochemistry and haematology in broilers exposed to individual and combined mycotoxicosis (aflatoxin, ochratoxin and T-2 toxin). Br. Poult. Sci., 41: 640-650.
PubMedDirect Link - Reddy, R.V., M.J. Taylor and R.P. Sharma, 1987. Studies of immune function of CD-1 mice exposed to aflatoxin B1. Toxicology, 43: 123-132.
PubMedDirect Link - Rossi, R., D. Giustarini, S. Fineschi, G. De Cunto, G. Lungarella and E. Cavarra, 2009. Differential thiol status in blood of different mouse strains exposed to cigarette smoke. Free Radical Res., 43: 538-545.
CrossRefDirect Link - Samuel, A.O., O. Olubukola and A.O. Matthew, 2009. Hematological and immunological effect on chicken exposed to aflatoxin. Vet. World J., 2: 5-7.
Direct Link - Satish, S., D.C. Mohana, M.P. Ranhavendra and K.A. Raveesha, 2007. Antifungal activity of some plant extracts against important seed borne pathogens of Aspergillus sp. J. Agric. Technol., 3: 109-119.
Direct Link - Reza, S.S.M., A. Masoud, T. Ali, G. Faranak and N. Mahboob, 2012. Determination of aflatoxins in nuts of Tabriz confectionaries by ELISA and HPLC methods. Adv. Pharmaceut. Bull., 2: 123-126.
CrossRefDirect Link - Silvotti, L., C. Petterino, A. Bonomi and E. Cabassi, 1997. Immunotoxicological effects on piglets of feeding sows diets containing aflatoxins. Vet. Rec., 141: 469-472.
CrossRefPubMedDirect Link - Tuzcu, M., E. Sur, I. Celik, Y. Oznurlu and M.K. Ciftci, 2010. Effects of aflatoxin on the proportions of peripheral blood leukocytes and Alpha-Naphtyl Acetate Esterase (ANAE) positive lymphocytes in the mouse. Kafkas Universitesi Veteriner Fakultesi Dergisi, 16: 337-341.
CrossRefDirect Link - Van Zwieten, R., A.J. Verhoeven and D. Roos, 2014. Inborn defects in the antioxidant systems of human red blood cells. Free Radic. Biol. Med., 67: 377-386.
CrossRefDirect Link - Venturini, M.C., M.A. Quiroga, M.A. Risso, C. Di Lorenzo, Y. Omata, L. Venturini and H. Godoy, 1996. Mycotoxin T-2 and aflatoxin B1 asimmunosuppressors in mice chronically infected with Toxoplasma gondii. J. Comp. Pathol., 115: 229-237.
CrossRefDirect Link - Wang, J.S. and J.D. Groopman, 1999. DNA damage by mycotoxins. Mutat. Res., 424: 167-181.
CrossRefDirect Link - Wang, J., G.M. Jin, Y.M. Zheng, S.H. Li and H. Wang, 2005. [Effect of bee pollen on development of immune organ of animal]. China J. Chin. Materia Medica, 30: 1532-1536, (In Chinese).
PubMedDirect Link - Wilson, A.S., D.P. Williams, C.D. Davis, M.D. Tingle and B.K. Park, 1997. Bioactivation and inactivation of aflatoxin B1 by human, mouse and rat liver preparations: Effect on SCE in human mononuclear leucocytes. Mutat. Res./Fundam. Mol. Mech. Mutagen., 373: 257-264.
CrossRefDirect Link - Xu, X., L. Sun, J. Dong and H. Zhang, 2009. Breaking the cells of rape bee pollen and consecutive extraction of functional oil with supercritical carbon dioxide. Innov. Food Sci. Emerg. Technol., 10: 42-46.
CrossRefDirect Link - Nakajima, Y., K. Tsuruma, M. Shimazawa, S. Mishima and H. Hara, 2009. Comparison of bee products based on assays of antioxidant capacities. BMC Complementary. Altern. Med., Vol. 9.
CrossRefDirect Link - Yousef, M.I., M.H. Salem, K.I. Kamel, G.A. Hassan and F.D. El-Nouty, 2003. Influence of ascorbic acid supplementation on the haematological and clinical biochemistry parameters of male rabbits exposed to aflatoxin B1. J. Environ. Sci. Health B, 38: 193-209.
PubMed