
J.P. Pandey
Department of Animal Sciences, School of Life Sciences,
University of Hyderabad, Hyderabad-5000 46, India
R.K. Tiwari
Department of Zoology, R.N. Govt. Postgraduate College, Gyanpur-221 304, S.R.N. Bhadohi, India
Dinesh Kumar
Department of Zoology, Banaras Hindu University, Varanasi-221, India
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 3 | Page No.: 200-206







ABSTRACT
The bioefficacy of some neem-based insecticides (NBIs) (neemazal, multineem and nimbecidine) was evaluated using fifth instar larvae and pupae of Danais chrysippus. All the NBIs caused reduction in total haemocyte count and much variation in their normal profile. This caused disintegration of haemocytes leading to reduction in immune response. In addition, the NBIs produced ecdysial stasis thereby resulting in the larval-pupal intermediates, pharate adults and deformed imagoes. A reduction in body weight of treated larvae was recorded too.
PDF Abstract XML References Citation
How to cite this article
J.P. Pandey, R.K. Tiwari and Dinesh Kumar, 2008. Reduction in Haemocyte Mediated Immune Response in Danais chrysippus Following Treatment with Neem-Based Insecticides. Journal of Entomology, 5: 200-206.
DOI: 10.3923/je.2008.200.206
URL: https://scialert.net/abstract/?doi=je.2008.200.206
DOI: 10.3923/je.2008.200.206
URL: https://scialert.net/abstract/?doi=je.2008.200.206
INTRODUCTION
The importance of neem tree (Azadirachta indica A. Juss) as antimicrobial and insect repellent is known to Indians since time immemorial. The azadirachtin, an active ingredient of this tree, because of its bioefficacy and biodegradability has now been considered as an alternative to conventional insecticides that are known to cause hazardous effects on human health and environment. Despite having an array of biological effects like antifeedancy, repellency, antifecundity, growth regulatory and prothoracicotropic hormone (PTTH) inhibitory activities as reported by a number of workers (Schmutterer, 1990; Singh, 1996; Cowles, 2004; Medina et al., 2004; Pandey et al., 2006; Shah et al., 2007), the effects of neem-based insecticides (NBIs) on haemocytes, the cells responsible for cellular immunity, have not been much studied (Azambuja et al., 1991; Sharma et al., 2003; Tiwari et al., 2006). The present study was, therefore, undertaken to investigate the effect of some NBIs on haemocyte’s population, their morphology as well as on metamorphic development of plain tiger butterfly, Danais chrysippus.
MATERIALS AND METHODS
The early larval instars of the plain tiger butterfly, Danais chrysippus Linn (Lepidoptera: Nymphalidae) were collected from Ak plant (Calotropis gigantea) from the field in the month of February and September. These larvae were reared on fresh leaves of the said plant in a BOD incubator maintained at 27±1°C, 75±5% RH and 16L: 8D photoperiod in laboratory (Pandey and Tiwari, 2005; Pandey et al., 2008). Fifth instar larvae (24 h old) and pupae (4-6 h old) from the said lab culture were used. The NBIs used in the present study were neemazal T’S (Azadirachtin 1%, other limonoids 3%, oil fatty acids glycerol esters 46.3%, polyethylene monosorbitol oleate 49.7%-EID Parry India Ltd.), multineem (seed extract containing 0.03% Azadirachtin-Multiplex Fertilizers Pvt. Ltd., Bangalore, India) and nimbecidine (0.03% Azadirachtin, neem oil 90.57%, hydroxyl 5%, epichlorohydrate 0.5%, aromax 3%). To examine their effects on haemocytes and metamorphic moulting, various concentrations (2.5, 5, 7, 10, 15, 20, 25 and 30%) of these insecticides were prepared by diluting them with acetone. While all the dilutions of insecticides were applied topically in different doses (vide Tables) with the help of glass micropipette on the dorsal and ventral surfaces at three different body regions (cephalic, thoracic and abdominal) of V instar larvae, prepupae and pupae of experimental group to investigate their effects on metamorphic moulting, only 50 μL of 5% of all the NBIs was used to study their toxic effect on haemocytes. The controls were treated with acetone only. Total haemocyte count (THC) and differential haemocyte count (DHC) were made at 24 hourly intervals in both the groups. The methods of haemolymph collection, smear formation, staining and counting of cells were similar to those applied earlier (Tiwari et al., 2006). The mean number of circulating haemocytes per mm3 was calculated using the formula of Jones (1962). Besides, the effects of NBIs on haemolymph cell’s morphology along with survival, moulting, degrees of development and wing shape in test insects were also studied. The body weight of the larvae of both the groups was also recorded at 24 h intervals till pupation to assess the toxic effect of NBIs.
RESULTS
Effect on Haemocyte Count
Application of 50 μL of 5% NBIs prevented the normally occurring increase in the number of haemocytes during fifth instar larval development resulting in over-all reduction in THC compared to control larvae (Table 1). Besides, the DHC revealed a much variation (Table 2).
| Table 1: | Effects of neem based insecticides (5% conc.) on THC in V instar larvae of D. chrysippus (Values are mean±SD for 20 larvae) |
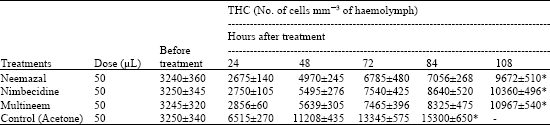 | |
| * = Indicates prepupa formation | |
| Table 2: | Effects of topical application of some neem based insecticides (5% conc. and 50 μL dose) on DHC in V instar larvae of D. chrysippus (Values are mean±SD for 10 larvae) |
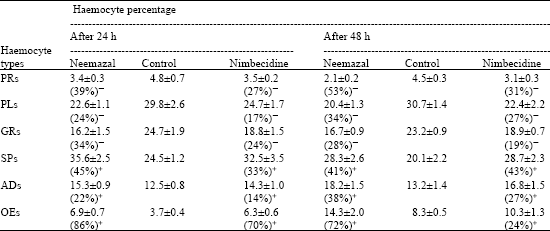 | |
| Value in parenthesis showed % decrease (¯) and % increase (+) in number of different haemocyte types | |
While a reduction occurred in the counts of prohaemocytes (PRs), plasmatocytes (PLs) and granulocytes (GRs); the spherulocytes (SPs), adipohaemocytes (ADs) and oenocytoids (OEs) showed rise in their counts in treated larvae. Neemazal caused 28% reduction in GRs and 53% in PRs after 48 h of treatment. Further, this NBI caused 72% rise in OEs, 41% in SPs and 38% in ADs respectively.
Effect on Haemocyte Morphology
NBIs caused different degrees of deformity in almost all the cell types in a dose dependent manner. Cellular clumping, vacuolization of all cells, loss of pseudopods in PLs and nuclear fragmentation especially in GRs were seen. Phagocytic capsules were not frequently seen showing apparent reduction in their number. Some GRs and SPs were found with lysed cellular contents coming out of them leaving empty spaces within the cells and many exocytotic vesicles on their periphery. A few GRs were seen only with their outer skeleton but their nuclei and other cytoplasmic contents were not visible perhaps due to degradation.
Effect on Morphogenetic Development
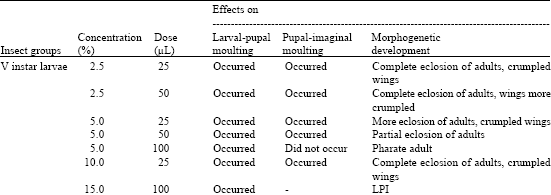
Topical application of NBI on ventral surface of cephalic and thoracic regions of the larvae was more effective than that on dorsal surface of abdominal region. The treatment caused movement of the head on the wall of rearing glass vials, increased defecation and jerking of the body intermittently. The effects of NBIs of different concentrations and doses on insect’s survival, moulting, degrees of development and wing shape were observed and the results were presented in Table 3-5.
| Table 3: | Effects of nimbecidine on morphogenetic development of D. chrysippus * |
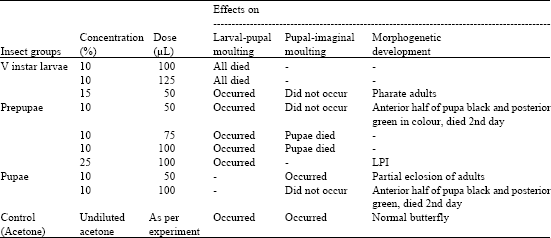 | |
| *: 15 insects were used in each experimental and corresponding control groups separately | |
| Table 4: | Effects of multineem on morphogenetic development of D. chrysippus * |
 | |
 | |
| *: 15 insects were used for each experimental and corresponding control groups, separately | |
| Table 5: | Effects of neemazal on morphogenetic development of D. chrysippus* |
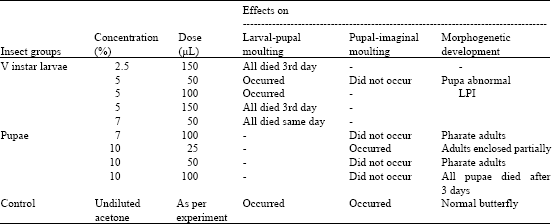 | |
| *: 15 insects were used for each experimental and corresponding control groups, separately | |
| Table 6: | Effects of neem-based insecticides on body weight of D. chrysippus larvae (Values are mean±SD for 20 insects) |
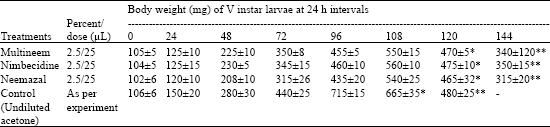 | |
| *: Prepupa formation, **: Pupa formation | |
Effect on Body Weight
Besides a delay of about one day in pupation, NBI treated larvae showed less consumption of food in comparison to acetone treated controls. While control larvae fed ad libitum attained a body weight of 715±25 mg, neemazal treated larvae could attained only 540±25 mg causing a reduction of 175 mg in their weight. Multineem and nimbecidine treated larvae, on the other hand, were heavier in weight than neemazal treated larvae (Table 6).
DISCUSSION
The response of haemocytes against biological agents as well as toxins has been studied in many insects. The resulting effects thereof are phagocytosis, encapsulation and distortion of cell contour or cellular disintegration (Saxena and Tikku, 1990). The drastic reduction in THC in present insect following NBIs application is similar to the reports of Azambuja et al. (1991) in Rhodnius prolixus, Sharma et al. (2003) in Spodoptera litura and Tiwari et al. (2006) in Dysdercus koenigii. The decrease in THC number may be due to the clumping of haemocytes, the toxic effect of NBIs and/or to their inhibitory effect on endocrine glands. Sharma et al. (2003) reported about 50% reduction in THC after 72 h of oral feeding of neem gold in S. litura but 61% decline was seen in D. koenigii following topical application of neemazal (Tiwari et al., 2006). Present results reveal that neemazal produced 59 and 56% reduction in THC after 24 and 48 h, respectively, which is more effective as compared to two other NBIs (vide Table 1). It could be probably because of more azadirachtin content (1% in the former and 0.03% in the latter two). However, Figueiredo et al. (2006) found no significant difference in THC between azadirachtin treated R. prolixus nymphs and their controls. The maximum drop of PR percentage (53%) was followed by PLs (34%) and GRs (28%) respectively, 48 h after neemazal treatment (Table 2). This indicates the maximum participation of PRs in over-all decline in cell count more likely by the inhibition of their mitotic divisions (Salehzadeh et al., 2003). The decline in PL- and GR-percentage was reported to be caused by their involvement in phagocytosis and nodule formation (Sharma et al., 2003). But the reduction in number of phagocytotic capsules in NBI treated D. chrysippus larvae led to suggest some other reasons for decline in their counts. Similar drastic reduction in encapsulation/ phagocytosis response were also found in Drosophila larvae infected by eggs of parasitic wasps (Sorrentino et al., 2002) and azadirachtin treated R. prolixus nymphs (Figueiredo et al., 2006). Based on their studies, Figueiredo et al. (2006) have suggested for the first time that the phagocytosis is modulated by ecdysone. The reduction in number of these cell types in the present study, therefore, seems to be due to the toxic effect of NBIs rather than their involvement in phagocytic actions.
A large number of abnormalities observed in NBIs treated Danais larvae at cellular and developmental levels (see Results) have also been reported by earlier workers (Schmutterer, 1988; Singh, 1996; Sahayaraj and Paulraj, 2001; Sharma et al., 2003; Tiwari et al., 2006). Further, the synthesis and release of PTTH by the brain was also reported to be deficient in azadirachtin treated R. prolixus nymphs (Garcia et al., 1990). The factors for these developmental and cellular defects are reported to lie in the endocrine system and the hormones secreted by them (Hoffmann, 1970; Schmutterer, 1990; Koul and Isman, 1991; Koul, 1996; Tiwari et al., 2006). Further, our results reveal the reduction of body weight in NBIs treated larvae. This is suggestive of their antifeedant property as reported for azadirachtin treated cutworm, Peridroma saucia (Koul and Isman, 1991) and Aza treated larvae of the lemon-butterfly, Papilio demoleus (Pandey et al., 2006).
It is, therefore, suggested that the NBIs might act negatively on the prothoracic glands via brain thereby causing reduction in phagocytic response of haemocytes and reduction in body weight. Further, NBIs might have direct toxic effects on haemocytes leading to necrotic results. The over-all effect of the NBIs could be adverse on the immune activity of the present insect threatening its survival.
ACKNOWLEDGMENTS
RKT thanks University Grants Commission, New Delhi for financial assistance and Principal of the college for providing necessary laboratory facilities. Authors also express their gratitude to anonymous referees for their fruitful suggestions and critical evaluation of the manuscript.
REFERENCES
- De Azambuja, P.D., E.S. Garcia, N.A. Ratcliffe and J.D. Warthen, 1991. Immune-depression in Rhodnius prolixus induced by the growth inhibitor azadirachtin. J. Insect Physiol., 37: 771-777.
CrossRef - Cowles, R.S., 2004. Impact of azadirachtin on vine weevil (Coleoptera: Curculionidae) reproduction. Agric. For. Entomol., 6: 291-294.
CrossRefDirect Link - Figueiredo, M.B., D.P. Castro, N.F.S. Nogueira, E.S. Garcia and P. Azambuja, 2006. Cellular immune response in Rhodnius prolixus: Role of ecdysone in hemocyte phagocytosis. J. Insect Physiol., 52: 711-716.
CrossRefDirect Link - Garcia, E.S., N. Luz, P. Azambuja and H. Rembold, 1990. Azadirachtin depresses the release of prothoracicotropic hormone in Rhodnius prolixus larvae: Evidence from head transplantations. J. Insect Physiol., 36: 679-682.
CrossRefDirect Link - Hoffmann, J.A., 1970. Regulations endocrines de la production et de la differenciation des hemocytes chez un insecte Orthoptere: Locusta migratoria migratoroides. Gen. Comp. Endocrinol., 15: 198-219.
CrossRefDirect Link - Jones, J.C., 1962. Current concepts concerning insect hemocytes. Am. Zool., 2: 209-246.
CrossRefDirect Link - Koul, O. and M.B. Isman, 1991. Effects of azadirachtin on the dietary utilization and development of the variegated cutworm, Peridroma saucia. J. Insect Physiol., 37: 591-598.
CrossRef - Medina, P., F. Budia, P. Del Estal and E. Vinuela, 2004. Influence of azadirachtin, a botanical insecticide on Chrysoperla carnea (Stephens). Reproduction, toxicity and ultrastructural approach. J. Econ. Entomol., 97: 43-50.
Direct Link - Pandey, J.P. and R.K. Tiwari, 2005. Feeding, brain implantation and 20-hydroxyecdysone treatment reverses the effect of starvation and ventral nerve cord severance on haemocyte counts in larvae of the plain tiger butterfly Danaus chrysippus (Lepidoptera: Nymphalidae). Int. J. Trop. Insect Sci., 25: 295-300.
CrossRefDirect Link - Pandey, J.P., G. Damodar, D. Manohar, A. Jagota and A.D. Gupta, 2006. Various Effects of Neem Based Pesticide Aza During the Postembryonic Development of Lemon-butterfly, Papilio demoleus (Lepidoptera: Papilionidae). In: Frontiers in Molecular Endocrinology of Invertebrates and Vertebrates, Gupta, A.D. and B. Senthilkumaran (Eds.). Department Animal Science, University, Hyderabad, pp: 6-12.
- Pandey, J.P., R.K. Tiwari and D. Kumar, 2008. Temperature and ganglionectomy stresses affect haemocyte counts in plain tiger butterfly, Danais chrysippus L. (Lepidoptera: Nymphalidae). J. Entomol., 5: 113-121.
CrossRefDirect Link - Salehzadeh, A., A. Akhkha, W. Cushley, R.L.P. Adams, J.R. Kusel and R.H.C. Strang, 2003. The antimitotic effect of the neem terpenoid azadirachtin on cultured insect cells. Insect Biochem. Mol. Biol., 33: 681-689.
CrossRefDirect Link - Saxena, B.P. and K. Tikku, 1990. Effect of plumbagin on haemocytes of Dysdercus koenigii F. Proc. Indian Acad. Sci. (Anim. Sci.), 99: 119-124.
CrossRefDirect Link - Schmutterer, H., 1988. Potential of azadirachtin-containing pesticides for integrated pest control in developing and industrialized countries. J. Insect Physiol., 34: 713-719.
CrossRef - Schmutterer, H., 1990. Properties and potential of natural pesticides from the neem tree, Azadirachta indica. Annu. Rev. Entomol., 35: 271-297.
CrossRefDirect Link - Shah, F.A., M. Gaffney, M.A. Ansari, M. Prasad and T.M. Butt, 2007. Neem seed cake enhances the efficacy of the insect pathogenic fungus Metarhizium anisopliae for the control of black vine weevil, Otiorhynuchs sulcatus(Coleoptera: Curculionidae). Biol. Control, 44: 111-115.
CrossRefDirect Link - Sharma, P.R., O.P. Sharma and B.P. Saxena, 2003. Effect of neem gold on haemocytes of the tobacco armyworm, Spodoptera litura (Fabricius) (Lepidoptera; Noctuidae). Curr. Sci., 84: 690-695.
Direct Link - Sorrentino, R.P., Y. Carton and S. Govind, 2002. Cellular immune responses to parasite infection in the Drosophila lymph gland is developmentally regulated. Dev. Biol., 243: 65-80.
CrossRefDirect Link - Tiwari, R.K., J.P. Pandey and D. Kumar, 2006. Effect of neem based insecticides on metamorphosis, haemocytes and reproductive behaviour in red cotton bug, Dysdercus koenigii Fab. (Heteroptera: Pyrrhocoridae). Entomon, 31: 267-275.
Direct Link