
Hemlata Bisht
Department of Botany and Microbiology, School of Life Sciences, HNBG Central University, Srinagar (G), Uttarakhand, 246-174, India
Vinay Prakash
Department of Seed Science and Technology, School of Agriculture and Allied Sciences, Chauras Campus, HNBG Central University, Srinagar (G), Uttarakhand, 246-174, India
A.R. Nautiyal
High Altitude Plant Physiology Research Centre, School of Agriculture and Allied Sciences, HNBG Central University, Srinagar (G), Uttarakhand, 246-174, India
Research Journal of Seed Science
Year: 2012 | Volume: 5 | Issue: 2 | Page No.: 63-70







ABSTRACT
Quercus semecarpifolia is a multipurpose tree species but facing the problem of poor regeneration in natural habitat. The seeds of Q. semecarpifolia shows the recalcitrant behavior as the seeds shed at high moisture content, short viable, desiccation and chilling sensitive and skip the maturation drying stage. To understand the regeneration potential of the species, seeds were examined under different temperature and light/dark conditions in intact form and after scarification and stratification. The finding indicates that the light, temperature, scarification and stratification have considerable effect on regeneration of Q. semecarpifolia when tested for radicle emergence and seedling recruitment under laboratory conditions. Higher (35°C) as well as lower (5°C) temperature did not favour radicle emergence, its growth and seedling establishment; however 15°C is best suited temperature to emerge radicle and root shoot development at which around 91% seeds resulted into seedlings. Scarification of seeds hardly matters for enhancement of radicle growth and seedling production significantly but light matters a lot. Light exhibits highest (90%) and fastest radicle growth (within 10 days) as well as shoot emergence (within 30 days) and survival percentage if compared with the germination in dark. Prechilling treatment at 0°C lost total viability even within 24 h.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: February 06, 2012;
Published: February 24, 2012
How to cite this article
Hemlata Bisht, Vinay Prakash and A.R. Nautiyal, 2012. Factors Affecting Regeneration Potential of Quercus semecarpifolia, Smith: A Poor Regenerated Oak of Himalayan Timberline. Research Journal of Seed Science, 5: 63-70.
URL: https://scialert.net/abstract/?doi=rjss.2012.63.70
URL: https://scialert.net/abstract/?doi=rjss.2012.63.70
INTRODUCTION
Quercus semecarpifolia, a high level Himalayan oak species forms timberline in many of the Western Himalayan regions. The species has wider distribution range from 1800 to 3500 m and phytosociological studies declare Q. semecarpifolia as the single abundant species or less associated species in the study area (Bisht, 2001). The species has a potential as a multipurpose tree for the production of fodder, tannin, fuel wood and wood for agricultural implements in the mountain areas, so the settlements in nearby areas highly dependent on the species for their livelihood. The species is considered as most older and overexploited plant of sub-alpine zones (Singh et al., 2010). Low regeneration, short viability, failure of a good seed crop every year, climate change, regular anthropogenic exploitation and the high magnitude of human pressure on the species could be dangerous to the existence of the species and in very near future the species may disappear from the subjected regions. Some of the studies were conducted on phytosociology and ecology of the species (Singh et al., 2010; Singh and Rawat, 2010; Bhatt and Ram, 2005; Shrestha, 2003) but the studies are limited on regeneration and factors affecting it particularly in the Western Himalayan perspective. The species need immediate attention for conservation of its genetic resources because of great role of this tree in the economy as well as ecology of Himalaya.
The seeds of Q. semecarpifolia behave as a recalcitrant seeds like many other oak species (Bisht, 2001). Similar to all recalcitrant seeds Q. semecarpifolia seeds shed at very high moisture content, undergo a biphasic seed development and lack maturation drying, short viable, very big in size, short storable, desiccation as well as chilling sensitive too (Bisht, 2001). Moreover the process of germination in these seeds was found to progress very slowly and was interesting as well as different than other oaks and associated species. In this species sprouting takes place immediately after shedding like white oaks (Tilki, 2010). After emergence radicle first grew to a length varying from 12-20 cm and subsequently began to grow in diameter. Shoot and root later emerged from thick portion of the radicle and the remaining portion of the radicle, towards seed gradually degenerated and seed was automatically detached from the radicle (Bisht, 2001). All these characteristics and low regeneration status of the species in nature make the seed interesting and initiate us to propose the study to determine the regeneration potential and impact of different factors on seedling establishment to develop conservation strategies for Q. semecarpifolia. Regeneration studies are very acceptable approach to understand the seed behavior and its requirement for germination (Chauhan et al., 2006) and seedling recruitment, for conservation (Sharma and Sharma, 2010; Srivastava et al., 2011), reintroduction and mass propagation (Zaman et al., 2011), technology development (Kumar et al., 2011) and effective management studies (Wei et al., 2009) of any of the species.
MATERIALS AND METHODS
Seeds (acorn sensu stricto) of Q. semecarpifolia (brown oak) were collected from the forest floor under the canopy of healthy tree, growing in the natural stand of Western Himalayan region near Duggalbitta (2590 masl) in district Rudraprayag, Uttarakhand, India. To collect the mature and immediately fallen fresh seeds, the forest floor was first swept to remove earlier fallen seeds and the seeds abscised within 24 h were collected for the regeneration studies. Natural shedding and easy sloughing out of cupule (a cup like structure by which approximately half of the seed enclosed) was considered as the maturity index of the seed.
To determine the effect of temperature on germination, mature seeds were kept at 5, 15, 25 and 35°C temperature in seed germinator. In addition to this, to determine the effect of light and dark, the seeds of Q. semecarpifolia were kept under light and dark conditions in the intact form and after removing the pericarp (scarified seeds) at 15°C temperature, which is the best suited temperature for germination. Mature intact seeds in light (IL), mature intact seeds in dark (ID) and mature scarified seeds in light (SL) and mature scarified seeds in dark (SD) were the conditions to analyze light/dark and scarification effect on regeneration.
Seeds were also sown after prechilling treatment for which, seeds were stored at 0°C in moist vermiculite and then sown to regenerate after 24 h to understand the behavior of seeds under the snow conditions in natural habitat during winter if the seeds not germinate early and lays as it is in situ.
For all the studies, the seeds were placed in styrofoam seedling trays filled with the mixture of vermiculite and sand (3:1) in triplicate, each replicate comprising 100 seeds. The sowing substratum (vermiculite and sand mixture) moistened regularly with distilled water. Radicle emergence resulted shoot/root emergence was noted as the criterion for germination because all the seeds of the Q. semecarpifolia already emerges radicle immediately after shedding from tree but all can not produce seedlings even after radicle emergence. Increase in radicle length and expansion in radicle diameter in the crucial and important stage resulted into seedlings. So, the observations were made for increase in radicle length, expansion in radicle diameter leading shoot/root emergence resulting successful seedling establishment in all these laboratory conditions. Statistical analysis was carried out by using ANOVA.
RESULTS
Seed germination leading radicle emergence, its growth and seedling production in Q. semecarpifolia showed disappointing results under natural condition on the forest floor (Bisht, 2001) in contrast to laboratory where not only the higher percentage of seeds resulted into germination but also the seedlings establishment was high. As high as 92% seeds had radicle emergence at laboratory within 16 days after sowing, resulting shoot emergence in 90% seeds within a month and only 2% degenerated after radicle emergence (Fig. 1, 2) but in situ only 5% seedlings were recovered even after next growing season (Bisht, 2001).
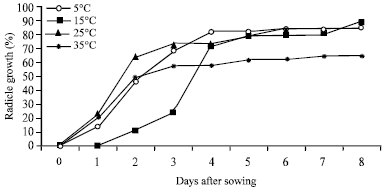 | |
| Fig. 1: | Percent seedling emergence in Q. semecarpifolia seed sown at 5, 15, 25 and 35°C temperature under laboratory conditions |
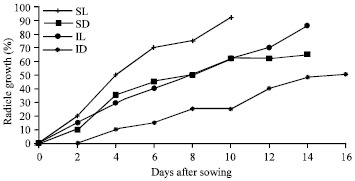 | |
| Fig. 2: | Percent radicle growth in intact and scarified seeds of Q. semecarpifolia under laboratory conditions in light and dark |
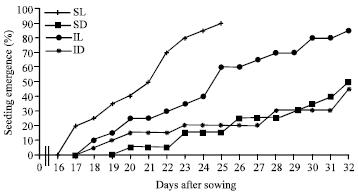 | |
| Fig. 3: | Percent seedling emergence from intact and scarified seeds of Q. semecarpifolia under laboratory conditions in light and dark |
Freshly collected seeds of Q. semecarpifolia when tested for the germinability under different temperatures revealed that the higher (35°C) as well as lower (5°C) temperature did not favour germination in these seeds. All the seeds emerge radicle irrespective of the conditions applied. Maximum percentage of seedling (90) was achieved at 15°C as compared to 86% at 5°C as well as 25°C (Fig. 1). Elevated temperature further reduced radicle growth of these seeds as only 66% seed achieved required length of radicle and later recovered as seedlings at 35°C. With temperature the days for onset of germination also varied. Germination under different temperature for 8 days had 80-99% correlation coefficient with significant variation with F = 4.22 (F critical = 3.00, p = 0.01) due to different temperature conditions, however, the variation was less pronounced. This is due to more or less similar germination percentage at different temperature conditions.
Seeds sown after removing the pericarp (scarified seeds) did not affect regeneration considerably but the scarified/intact seeds under light and dark conditions at room temperature exhibited significantly different results in respect of radicle growth and shoot emergence and has been depicted in Fig. 2. Highest and fastest radicle growth was recorded in the seeds sown in scarified state under the light (92%) within 10 days. Contrary to this, intact seeds in dark had lowest and slowest performance and took 16 days to reach upto maximum (50%). The results are highly significant (F = 30.44, F critical = 3 at p level<0.01) due to around 87-89% positive correlation coefficient between ID (mature intact seeds in dark) and SL (mature scarified seeds in light) as well as ID (mature intact seeds in dark) and SD (mature scarified seeds in dark) and maximum 99% positive correlation between SD and SL.
Percent seedling emergence from intact and scarified seeds of Q. semecarpifolia under laboratory conditions in light and dark has been depicted in Fig. 3. Shoot emergence also followed trend similar to that of radicle emergence and its growth wherein shoot emergence in light germinated excised seeds was earliest and in highest percentage (90% within 25 days) while dark germinated intact as well as excised seeds had lowest and slowest emergence of shoots (45 and 55%, respectively within 32 days). Although, removal of pericarp does not seem to be essential for the germination of the seeds of Q. semecarpifolia, it had promoting effect on germinability of these seeds. Emergence of shoots took place after 17 days in SL, 18 days in ID and IL and 25 days in SD conditions. Also, unlike the radicle emergence differences in the percentage of seeds resulting into shoot emergence in light germinated intact seeds and dark germinated excised seeds were considerably high and not only the final percentage varied in the two but even initially the differences were high.
DISCUSSION
Regeneration studies on natural regeneration status of Q. semecarpifolia conducted previously by Thakuri (2010), Singh and Rawat (2010), Tashi (2004), Shrestha (2003), Bisht (2001) and Vetaas (2000) and agreed on very low regeneration under different microclimatic conditions. High percentage of seeds degenerated on the forest floor even after sprouting, percentage of such seeds ranging from 74-96% depending on the condition in which seeds were sown in-situ (Bisht, 2001). Unlike in situ, 90% seedlings were recovered within one month in laboratory conditions. This is due to promoting effect of light and temperature on germination of the seeds in controlled laboratory condition which must have ecophysiological implications. In natural conditions also, seeds must germinate immediately during rainy season to avoid the desiccation thereafter and escape to harsh chilling temperate environment as the seed of Q. semecarpifolia are chilling and desiccation sensitive (Bisht, 2001). Light had significant influence on radicle growth and seedling establishment of Q. semecarpifolia seeds wherein a higher percentage of seeds germinated under light than in dark like many other temperate tree species viz., Q. robur (Kuhne and Bartsch, 2007) Betula papyrifera (Bevington and Hoyle, 1981), Picea marina (Farmer et al., 1984) and Pinus contorta (Li et al., 1994).
However, in most of these cases, stratification of the seeds resulted in identical germination percentages in light as well as dark. Absolute darkness and 90 days prechilling treatment ceased germination but alteration of light dark conditions enhance germination in a woody species, Vaccinium arctostaphylos (Shahram, 2007). Evidences of seed germination enhancement in continuous dark and reduction in continuous light were presented but in presence of growth regulators in Digitaria exilis by Idu et al. (2008). It is evident from these studies that even if light is not essential for germination but it may have a promoting or retarding effect on germinability of seeds.
Studies reviewed by Baskin and Baskin (1998) indicate that seeds of different temperate and sub-alpine species have different responses to light and dark as well as varying temperature regimes. For example, a trans Himalayan medicinal plant, Bunium persicum germinate only at 4°C and shifting of seeds from 4-25°C ceased germination (Sharma and Sharma, 2010) but Q. semecarpifolia respond well at 15°C in presence of light however lower (>15°C) as well as higher (<30°C) temperature did not favour the regeneration similar to the Campsis radicans (Chachalis and Reddy, 2000). However, in crop plants like Glycine max and Pisum sativum reduction in percentage and rate of germination was noticed at high temperature (Beena and Jayaram, 2010). But the germination in Brassica tournefortii does not affected by light at optimum temperature but at low temperature, light inhibit germination (Chauhan et al., 2006).
Regeneration process in this species in nature begins with radicle emergence which takes place immediately after seed shedding during spring in the month of July-August unlike most of the oak species viz., Q. leucotrichophora, Q. robur, Q. glauca etc. in which seed shedding occurred during winter in November-December. Radicle emergence followed by thickening of radicle and most of the seeds remain in this condition over winter. Seedling emergence (roots and shoot emergence) from this swollen portion takes place after a longitudinal split only in the subsequent year in natural condition (Bisht, 2001) when light intensity and temperature conditions increase in the timberline after snowmelt. It can be presumed that this thick portion of radicle requires a chilling treatment for seedling emergence as an adaptive strategy to over-winter under its natural conditions/habitat where such conditions prevail subsequent to its germination (Bisht, 2001). However, the freshly collected seeds of Q. semecarpifolia sown in the laboratory at high temperature conditions (30°C) underwent normal course of events leading to seedling emergence and did not require any chilling or low temperature cycle/condition for the same. The process of regeneration in these seeds was found to progress slow and low in nature but in favourable light and temperature conditions in laboratory high as well as early seedling establishment occurred. This rules out any such possibility of low temperature requirement in these seeds but probably an adaptive strategy against typical temperate conditions. The seeds of some other temperate plants viz., Quercus, Trillium, Viburnum, Convallaria and Polygonatum species also exhibit epicotyle dormancy where seeds germinate and put out a radicle in the autumn without prior chilling (Baskin and Baskin, 1998). However, development of the epicotyle depends upon a chilling treatment and this does not normally occur during the spring. Seedlings of these species do not emerge above the ground before the second spring after seed dispersal (Panneerselvam, 1998).
Moreover, scarification of the seeds in Q. semecarpifolia did not contribute to enhance the regeneration but in other Himalayan oaks in Q. glauca and Q. leucotrichophora, removal of pericarp improved germinability of the seeds (Rawat et al., 1998). Pericarp in this case posed as mechanical resistant for cotyledon expansion restricting water uptake by the seed during radicle emergence. But in Q. semecarpifolia scarification did not affect regeneration may be because of high moisture content and non requirement of water for radicle emergence as the seed emerge radicle immediately after shedding without water.
Q. semecarpifolia seeds lost their viability on prechilling treatment at 0°C even within 24 h. This is because of ice crystal formation in the cells at/below 0°C due to high moisture content of the seeds at shedding (Bisht, 2001). Contrast to this, in Jasminus fruticans seed germination enhances after 3 months cold stratification (Pipinis et al., 2009) and seed dormancy in a temperate oak, Q. ilex, broken by stratification of scarified seeds and seedling growth also positively affected (Ghasemi and Khosh-Khui, 2007). Similar to a deciduous Iranian mountain tree, Pistacia khinjuk (Baninasab and Rahemi, 2008). Stratification of lower altitude plant, Pterocarya fraxinifolia also maximize seed germination rate as well as percentage (Cicek and Tilki, 2008). Scarification and stratification were efficient to promote germination and seedling growth in many of the angiosperms and gymnosperms (Esen et al., 2007) and also not only in herbaceous plant but in tree species too viz. Rubia tinctorum (Sadeghi et al., 2009), Solanum nigrum (Suthar et al., 2009), Prunus scoparia and P. webbii (Heidari et al., 2008), Parkia biglobosa (Okunlola et al., 2011), Convolvulus oxyphyllus and Aeluropus lagopoides (Zaman et al., 2011), Lupinus leptophyllus (Alderete-Chavez et al., 2010). These observations declare that the requirement/factors of seed germination vary individually irrespective of the similarity in plant habit or habitat. So, the development, management and implementation of strategies for conservation must be species specific and also area specific.
CONCLUSION
It is evident from findings that although radicle emergence took place in a high percentage of seeds irrespective of the conditions, only a few could result in production of a seedling even after radicle emergence. Although, only a few seeds failed to emerge radicle, the deterioration of radicle caused maximum mortality. Scarification does not affect radicle growth as well as seedling emergence significantly but light and temperature contribute to enhance the seedling establishment in totality. Identification of responsible factors for poor regeneration even after radicle emergence and the mechanism associated with it would further help in understanding the ecophysiology of its regeneration and to develop conservational strategies of this species and restoration of timberline ecosystem.
ACKNOWLEDGEMENT
We thank Ministry of Non-Conventional Energy Sources, Government of India and Council for Scientific and Industrial Research, New Delhi, India for the financial assistance during the study.
REFERENCES
- Alderete-Chavez, A., D.A. Rodriguez-Trejo, V. Espinosa-Hernandez, E. Ojeda-Trejo and N. de la Cruz-Landero, 2010. Effects of different scarification treatments on the germination of Lupinus leptophyllus Seeds. Int. J. Botany, 6: 64-68.
CrossRefDirect Link - Bevington, J.M. and M.C. Hoyle, 1981. Phytochrome action during prechilling induced germination of Betula papyrifera, Marsh. Plant Physiol., 67: 705-710.
Direct Link - Bhatt, J. and J. Ram, 2005. Seed characteristics and germination in Quercus leuco-trichophora A. Camus along the elevation gradient in the Uttranchal Himalaya. Bull. Nat. Inst. Ecol., 15: 207-213.
Direct Link - Chachalis, D. and K.N. Reddy, 2000. Factors affecting Campsis radicans seed germination and seedling emergence. Weed Sci., 48: 212-216.
CrossRefDirect Link - Chauhan, B.S., G. Gill and C. Preston, 2006. African mustard (Brassica tournefortii) germination in southern Australia. Weed Sci., 54: 891-897.
Direct Link - Cicek, E. and F. Tilki, 2008. Influence of stratification on seed germination of Pterocarya fraxinifolia (Poiret) spach, a relic tree species. Res. J. Bot., 3: 103-106.
CrossRefDirect Link - Esen, D., O. Yildiz, M. Sarginci and K. Isik, 2007. Effects of different pretreatments on germination of Prunus serotina seed sources. J. Environ. Biol., 28: 99-104.
PubMedDirect Link - Farmer, R.E., P. Charrette, I.E. Searle and D.P. Tarjan, 1984. Interaction of light, temperature and chilling in the germination of black spruce. Can. J. For. Res., 14: 131-133.
Direct Link - Ghasemi, M. and M. Khosh-Khui, 2007. Effect of stratification and growth regulators on seed germination and seedling growth of Quercus ilex L. J. Plant Sci., 2: 341-346.
CrossRef - Idu, M., J.U. Chokor and O. Timothy, 2008. Effects of various hormones on the germination of Fonio-Digitarie exilis L. Int. J. Bot., 4: 456-460.
Direct Link - Kuhne, C. and N. Bartsch, 2007. Germination of acorns and development of oak seedlings (Quercus robur L.) following flooding. J. For. Sci., 53: 391-399.
Direct Link - Li, X.J., P.J. Burton and C.L. Leadem, 1994. Interactive effects of light and stratification on the germination of some British Columbia conifers. Can. J. Bot., 72: 1635-1646.
Direct Link - Okunlola, I., R.A. Adebayo and A.D. Orimogunje, 2011. Methods of braking seed dormancy on germination and early seedling growth of African locust bean (Parkia biglobosa) (JACQ.) Benth. J. Hortic. For., 3: 1-6.
Direct Link - Shrestha, B.B., 2003. Quercus semecarpifolia Sm. in the himalayan region: Ecology, exploitation and threats. Himalayan J. Sci., 1: 126-128.
Direct Link - Singh, G., G.S. Rawat and D. Verma, 2010. Comparative study of fuelwood consumption by villagers and seasonal Dhaba owners in the tourist affected regions of Garhwal Himalaya, India. Energy Policy, 38: 1895-1899.
CrossRef - Suthar, A.C., V.R. Naik and R.M. Mulani, 2009. Seed and seed germination in Solanum nigrum Linn. Am-Eurasian J. Agric. Environ. Sci., 5: 179-183.
Direct Link - Tilki, F., 2010. Influence of acorn size and storage duration on moisture content, germination and survival of Quercus petraea (Mattuschka) Liebl. J. Environ. Biol., 31: 325-328.
Direct Link - Vetaas, O.R., 2000. The effect of environmental factors on the regeneration of Quercus semecarpifolia Sm. in central Himalayan, Nepal. Plant Ecol., 146: 137-144.
Direct Link - Wei, S., C. Zhang, X. Li, H. Cui, H. Huang, B. Sui, Q. Meng and H. Zhang, 2009. Factors affecting buffalobur (Solanum rostratum) seed germination and seedling emergence. Weed Sci., 57: 521-525.
Direct Link