
T. Ling
School of Environmental and Municipal Engineering, Institute of Environmental Ecology, Lanzhou Jiaotong University, Lanzhou 730070, Peoples� Republic of China
Y. Fangke
School of Environmental and Municipal Engineering, Institute of Environmental Ecology, Lanzhou Jiaotong University, Lanzhou 730070, Peoples� Republic of China
R. Jun
Demonstration Center for Experimental, Teaching of Environmental Science and Technology in Colleges of Gansu Province, Lanzhou 730070, Peoples� Republic of China
Research Journal of Phytochemistry
Year: 2010 | Volume: 4 | Issue: 4 | Page No.: 225-233







ABSTRACT
The present investigation reports the results of the effects of mercury on seed germination, coleoptile growth and root elongation of four vegetables. The effects of seven serial doses of mercury (0, 0.1, 0.2, 0.4, 0.8, 1.6 and 3.2 mM) on four vegetable crops were studies. Seeds were surface sterilized and washed with distilled water. The seeds were germinated in Petri dishes with double layer of filter paper soaked in distilled water (control) and 0.1-3.2 mM HgCl2 solutions. The seeds were set under a photoperiod of 12 h and 32±1/25±1°C day/night temperature. The root and shoot length of seedling were measured and the germination percentage was recorded after 96 h. The different concentration of Hg2+ showed reduction in coleoptile growth and root elongation with increase in concentration of Hg2+. All the treated species were significantly inhibited when the Hg2+ concentration up to 0.8 mM. Brassica oleracea was the most sensitive species to mercury among the four test species, B. campestris was the most resist species to mercury pollution. These four vegetables were more sensitive to mercury stress in coleoptile growth and root elongation than seed germination.
PDF Abstract XML References Citation
Received: April 20, 2010;
Accepted: June 30, 2010;
Published: July 17, 2010
How to cite this article
T. Ling, Y. Fangke and R. Jun, 2010. Effect of Mercury to Seed Germination, Coleoptile Growth and Root Elongation of Four Vegetables. Research Journal of Phytochemistry, 4: 225-233.
URL: https://scialert.net/abstract/?doi=rjphyto.2010.225.233
URL: https://scialert.net/abstract/?doi=rjphyto.2010.225.233
INTRODUCTION
Following the industrialization, the anthropogenic factor has become the most important source of heavy metals in the environment. Heavy metal contamination of soil, water and air has caused serious environmental hazard in the biosphere due to rapid industrialization and urbanization. Heavy metals might accumulate in the food chains, with risks for the health of animals and humans, which are less sensitive to metal toxicity than plants, but they are capable of concentrating heavy metals in certain tissues and organs (Xiong, 1998; Peralta et al., 2001; Parmar et al., 2002; Liu et al., 2006; Street et al., 2007; Emese et al., 2009). The influence of metals on development and reproduction of plants can be firstly quantified by determining the germination traits of seeds and growth performance of seedling. In the presence of high concentrations of some heavy metals, most of plant species performed the reduction of seed germination and seedling growth (Patra and Sharma, 2000; Peralta et al., 2001; Abedin and Meharg, 2002; Jeliazkova et al., 2003; Shafiq and Iqbal, 2005; Marchiol et al., 2006; Marques et al., 2007; Ashraf and Ali, 2007; Hussain et al., 2007; Mahmood et al., 2007; Marques et al., 2007; Singh Sengar et al., 2008; Jun et al., 2009).
Mercury (Hg) is a toxic heavy metal that is of significant concern as an environmental pollutant. Since mercury is not very phytotoxic in normally occurring concentrations. In polluted regions, mercury is a non-degradable toxic heavy metal pollutant when it is accumulated by plants. The information is scare about its uptake mechanism and growth inhibition. There are a wide range of sources that emit mercury to the atmosphere. Approximately half of the atmospheric budget of vapor-phase mercury is attributed to anthropogenic sources and half to natural sources (Nriagu, 1989). Mercury is readily transported in the atmosphere (Carpi et al., 1997) and has an atmospheric half-life of approximately one year (Lindqvist and Pehkonen, 1999).
Mercury is not essential to living cells and performs no known biological function. Mercury has a strong affinity for sulfur and mercury’s primary mode of toxic action in living organisms is thought to be the interference of enzyme function and protein synthesis by binding to sulfhydryl groups (Sharma, 1985; Garcia and Reyes, 2001; Patra et al., 2004). Maximum work has been carried out on seed germination and seedling growth of different plant species in field exposed to mercurials (Sharma, 1985; Fargasova, 1994; Helal, 1995; Mishra and Choudhuri, 1997a, b, 1998, 1999; Bonifacio and Montano, 1998; Jain et al., 1998; Al-Yemeni, 2001; Munzuroglu and Gechil, 2002; Neelima and Reddy, 2003; Li et al., 2005; Bhanumathi and Jayabalan, 2007; Devi et al., 2007; Bandana et al., 2008; Umadevi et al., 2009). The present study was undertaken with a view to find out the toxic effect of mercury to seed germination, root elongation and coleoptile growth of four vegetables. With the specific aim of determining: 1) what degree mercury inhibits the four vegetables’ seed germination and seedling growth? 2) Which vegetables are more sensitive to mercury?
MATERIALS AND METHODS
The effects of seven serial doses of mercury (0, 0.1, 0.2, 0.4, 0.8, 1.6 and 3.2 mM) on four vegetable crops were studies. Seeds of Cabbage (Brassica rapa), Cole (B. napus), Head Cabbage (B. oleracea) and Spinach (Spinacia oleracea) were offered by Gansu Academy of Agricultural Sciences of China. The experiment was conducted in Institute of Environmental Ecology of Lanzhou Jiaotong university from Gansu for 4 months. Seeds were surface sterilized in 0.5% sodium hypochlorite solution for 20 min and washed thoroughly with distilled water. The seeds were germinated in Petri dishes (diameter is 150 mm) with double layer of filter paper soaked in distilled water (control) and 0.1-3.2 mM HgCl2 solutions. The seeds were set under a photoperiod of 12 h and 32±1/25±1°C day/night temperature. The root and shoot length of seedling were measured and the germination percentage was recorded after 96 h. A 1-mm radical emergence from seeds was considered seed germination. The root and coleoptile length of 10 germinated seeds which selected stochastically were measured at 96 h under each treatment.
Data are the results from six separate analyses with 50 seeds in each Petri. Statistical analysis was performed based on STATISTICA. The data were analyzed through one-way Analysis of Variance (ANOVA) to determine the effect of treatments and Duncan’s multiple comparison test were performed to determine the statistical significance of the differences between means of treatments.
RESULTS AND DISCUSSION
Germination under Mercury Stress
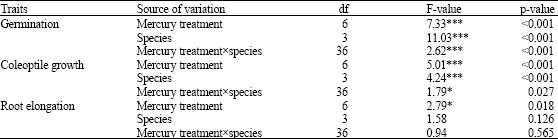
The germination percentage of vegetable seeds occurred at all mercury treatments from 0.0 mM to 3.2 mM for each species (Table 2). A two-way ANOVA showed that seed germination was significantly affected by different species (F3 =7.33, p<0.001), mercury treatment (F5 =11.03, p<0.001) and interaction between species and mercury treatment (F36 =2.62, p<0.001) (Table 1). The germination percentage was significantly different among mercury treatments in four vegetable species (one-way ANOVA: F6,36 = 13.26, p<0.001 for Brassica oleracea; F6,36 = 14.29, p<0.001 for B. campestris; F6,36 = 56.62, p<0.001 for B. rapa; F6,36 = 18.69, p<0.001 for Spinacia oleracea). For all species, the lowest germination percent always occurred in highest mercury concentration treatments. Compared with the control, higher concentration mercury treatments (0.8, 1.6, 3.2 mM) significantly inhibited germination for Brassica oleracea, B. rapa and Spinacia oleracea. The germination of all four species was not affected by lower mercury concentration treatments (0.1 mM) and was significantly inhabited by the highest mercury treatment (3.2 mM) comparing control and other treatments.
| Table 1: | The analysis of variance for the effects of different species, mercury treatments and their interaction on seed germination (%), coleoptile growth and root elongation for four vegetable species |
 | |
| *Means significant correlation at 0.05 level. **Means significant correlation at 0.01 level. ***Means significant correlation at 0.001 level | |
| Table 2: | Influence of mercury on seed germination, coleoptile growth and root elongation of four vegetables |
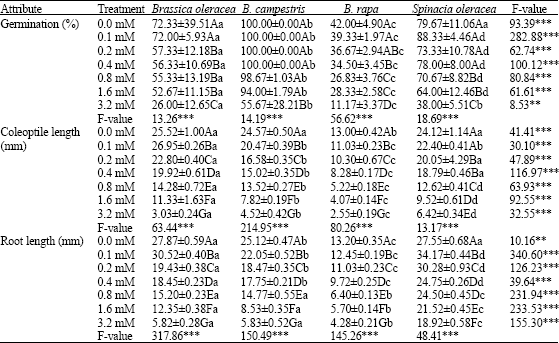 | |
| Results are Means±SD. For each attribute the mean values with the same lowercase letters among species in same column are not significantly different at 5% level of probability (Duncan’s multiple comparisons test) and with the same upper-case letters among treatments in same row are not significantly different. *Means significant correlation at 0.05 level. **Means significant correlation at 0.01 level. ***Means significant correlation at 0.001 level | |
| Table 3: | Correlations of mercury concentration (Y) to germination percentage (X), root and shoot length (X) |
 | |
B. rapa performed poorer germination at all mercury treatments and control (Table 2).
The germination was significantly different among species at six kinds of mercury treatments and one control. At control, the germination percentages of Brassica oleracea and Spinacia oleracea did not perform significant difference, but significantly lower than B. campestris and higher than B. rapa. At 0.1, 0.2, 0.4, 0.8 and 1.6 mM mercury treatments, there was significant difference of the germination percentages among four species and B. campestris always performed the highest germination percentage, B. rapa performed the lowest germination percentage. At the highest concentration mercury treatments (3.2 mM), Brassica oleracea and B. rapa did not performed significant difference and were significantly lower than other 2 species (Table 2). There was a significant negative correlation between the mean percent germination and mercury concentration for all four vegetable species (Table 3).
Coleoptile Growth under Mercury Stress
A two-way ANOVA showed that coleoptile growth was significantly affected by different mercury treatment (F6 =5.01, p<0.001) and species (F3 =4.25, p<0.001), but not by the interaction between species and mercury treatment (Table 1). The coleoptile length was significantly different among mercury treatments in each vegetable species (one-way ANOVA: F6,36 = 63.44, p<0.001 for Brassica oleracea; F6,36 = 214.95, p<0.001 for B. campestris; F6,36 = 80.26, p<0.001 for B. rapa; F6,36 = 13.17, p<0.001 for Spinacia oleracea). The coleoptile growth of all 4 vegetables dramatically and significantly decreased with the addition of mercury concentration. Compared with the control, lower concentration mercury treatments (0.1 mM) significantly stimulated the coleoptile growth of Brassica oleracea, but other 3 species did not perform same responses. For Brassica oleracea, there was significant difference of coleoptile growth among 6 treatments and the control, but the highest coleoptile length appeared at 0.1 mM mercury treatment and the lowest coleoptile length appeared at the highest concentration treatment (3.2 mM). For B. campestris and B. rapa, the highest coleoptile length appeared at control and the lowest coleoptile growth was always caused by the highest concentration treatment (3.2 mM). The lowest concentration mercury treatment (0.1 mM) did not significantly effect the coleoptile growth of Spinacia oleracea. Comparing the lower concentration treatments and control, three higher concentration treatments (0.8, 1.6 and 3.2 nM) performed siginificantly lower coleoptile growth and significant difference each other (Table 2).
The coleoptile growth was significantly different among species at six kinds of mercury treatments and one control. At control, Brassica oleracea, B. campestris and Spinacia oleracea did not performed significant difference in the coleoptile growth and their coleoptile lengths were significantly higher than the coleoptile length of B. rapa. At 0.1 mM treatment, B. campestris and Spinacia oleracea did not performed significant difference in the coleoptile growth and their coleoptile lengths were significantly higher than Brassica oleracea and B. rapa and there was significant difference between Brassica oleracea and B. rapa. At 0.2 and 0.4 mM treatments, the coleoptile lengths of Brassica oleracea and Spinacia oleracea were significantly higher than the coleoptile lengths of B. campestris and B. rapa. At higher concentration mercury treatments (0.8, 1.6 and 3.2 mM), there was significant difference of the coleoptile growth among four species and At 0.8, 1.6 and 3.2 mM treatments, Brassica oleracea always performed the biggest coleoptile growth, B. rapa performed the smallest coleoptile growth (Table 2). There was a significant negative correlation between the mean coleoptile length and mercury concentration for all four vegetable species (Table 3).
Root Elongation under Mercury Stress
A two-way ANOVA showed that root elongation was significantly affected by different mercury treatment (F6 =2.80, P=0.018), but not by the species and the interaction between species and mercury treatment (Table 1). The root elongation was significantly different among mercury treatments in each vegetable species (one-way ANOVA: F6,36 = 317.86, p<0.001 for Brassica oleracea; F6,36 = 150.49, p<0.001 for B. campestris; F6,36 = 145.26, p<0.001 for B. rapa; F6,36 = 48.41, p<0.001 for Spinacia oleracea). The root elongation of all 4 vegetables dramatically and significantly decreased with the addition of mercury concentration. Compared with the control, lower concentration mercury treatments (0.1 mM) significantly stimulated the root elongation of Brassica oleracea and Spinacia oleracea, but other 2 species did not perform same responses. For Brassica oleracea, there was significant difference of root elongation among 6 treatments and the control, but the highest root elongation appeared at 0.1 mM mercury treatment and the lowest root elongation appeared at the highest concentration treatment (3.2 mM). For B. campestris and B. rapa, the highest root length appeared at control and the lowest root length was always caused by the highest concentration treatment (3.2 mM). Comparing the lower concentration treatments and control, three higher concentration treatments (0.8, 1.6 and 3.2 nM) performed significantly lower root elongation and significant difference each other (Table 2).
The root elongation was significantly different among species at six kinds of mercury treatments and one control. At control, Brassica oleracea and Spinacia oleracea did not performed significant difference in the root elongation and were significantly higher than B. campestris and B. rapa. At lower concentration mercury treatments (0.1, 0.2, 0.4 mM), there was significant difference of the root elongation among four species, Brassica oleracea always performed the biggest root elongation, B. rapa performed the smallest root elongation. At higher concentration mercury treatments (0.8, 1.6 and 3.2 mM), B. campestris and Spinacia oleracea did not performed significant difference in the root elongation and their root lengths were significantly higher than B. rapa and lower than Spinacia oleracea (Table 2). There was a significant negative correlation between the mean root length and mercury concentration for all four vegetable species (Table 3).
In this study, we have examined the toxicity of mercury on seed germination, coleoptile growth and root elongation in four vegetable species. Depending on the Hg2+ concentration and the plant species, the inhibition effects of mercury on vegetable were different. Seed germination, root and coleoptile growth of vegetable are susceptible to Hg2+ pollution. The inhibitory effect on root and coleoptile growth of vegetables was more pronounced at higher concentrations of Hg2+. Root and coleoptile length of seedlings decreased with increasing concentrations of Hg2+. Mercury toxicity for shoot and root cell may result from the displacement of other cations from binding sites in the plasma membranes and cell wall (Sharma, 1985).
In this study, we have examined the toxicity of mercury on seed germination, coleoptile growth and root elongation in four vegetable species. Depending on the Hg2+ concentration and the plant species, the inhibition effects of mercury on vegetable were different. Seed germination, root and coleoptile growth of vegetable are susceptible to Hg2+ pollution. The inhibitory effect on root and coleoptile growth of vegetables was more pronounced at higher concentrations of Hg2+. Root and coleoptile length of seedlings decreased with increasing concentrations of Hg2+. Mercury toxicity for shoot and root cell may result from the displacement of other cations from binding sites in the plasma membranes and cell wall (Sharma, 1985).
Seed germination had been no significantly inhibited by the lower levels ( = 0.4 mM) of Hg2+, but at the same Hg2+ concentration, the root and shoot growth was significantly decreased comparing to the control. In general, our results were consistent with previous reports regarding the effect of heavy metals (Fargasova, 1994; Helal, 1995; Mishra and Choudhuri, 1997a, b, 1999; Bonifacio and Montano, 1998; Al-Yemeni, 2001; Jeliazkova and Craker, 2002; Munzuroglu and Gechil, 2002; Li et al., 2005). Among the different vegetable seedling, the shoot appeared to suffer the most severe growth inhibition. To this concern, it had been suggested that the reduction in coleoptile growth could be due to inhibition of mercury on coleoptile cell division or elongation, or on the extension of cell cycle. Our results indicated that mercury affected root and coleoptile growth more than seed germination was not a reliable indicator for metal tolerance in early root development of plants. Radicle and plumule growth, the primary plant organs that sense, contact and accumulate heavy metal(s) from the substrate, are thought to be a reliable indicator of metals tolerance in plants (Patra et al., 2004; Ozdener and Kutbay, 2009), may be it’s because of the seed coat is able to reduce the amount of mercury entering the seed, but after the seed germination, radicle and plumule have no barrier to protect. Some other studies have the same conclusion (Leon et al., 2005).
Mercury has inhibitory effect on the germination and seedling growth of the test species. Germination inhibition due to heavy metals has been reported by many workers (Fargasova, 1994; Helal, 1995; Mishra and Choudhuri, 1997a, b, 1999; Bonifacio and Montano, 1998; Al-Yemeni, 2001; Jeliazkova and Craker, 2002; Munzuroglu and Gechil, 2002; Li et al., 2005). Ionic toxicity might be the cause of drastic effects of salts on seed germination (Mishra and Choudhuri, 1997a), or it might be due to osmotic effect (Shaukat et al., 1999). Mercury also causes alterations in protein structure, alterations in calcium transport, along with the inhibition of glucose transport and enzyme function. The direct mechanism involving mercury’s inhibition of cellular enzymatic processes by binding with the hydroxyl radical in amino acids appears to be a major part of the connection to allergic/immune reactive conditions (Sharma, 1985; Garcia and Reyes, 2001; Patra et al., 2004).
From this study it is concluded that seed germination, root and coleoptile growth of the vegetable species were significantly reduced by mercury and different species show different levels of tolerance to mercury, Brassica oleracea was the most sensitive species to mercury among the four test species, B. campestris was the most resist plants to mercury pollution and mercury stress is more sensitive to the coleoptile growth and root elongation.
ACKNOWLEDGMENTS
This research was financially supported by the National Natural Science Foundation (No. 30970490) and by The Program for Changjiang Scholars and Innovative Research Team in University (IRT0966).
REFERENCES
- Abedin, M.J. and A.A. Meharg, 2002. Relative toxicity of arsenite and arsenate on germination and early seedling growth of rice (Oryza sativa L.). Plant Soil, 243: 57-66.
CrossRefDirect Link - Al-Helal, A.A., 1995. Effect of cadmium and mercury on seed germination and early seedling growth of rice and alfalfa. J. Univ. Kuwait (Sci.), 22: 74-83.
Direct Link - Al-Yemeni, M.N., 2001. Effects of cadmium, mercury and lead on seed germination and early seedling growth of Vigna ambacensis L. Indian J. Plant Physiol., 6: 147-151.
Direct Link - Marques, P.G.C., O.S.S. Rangel and M.L. Castro, 2007. Effect of arsenic, lead and znic on seed germination and plant growth in black nightshade (Solanum nigrum L.) vs. clover (Trifolium incarnatum L.). Fresenius Eviromental Bull., 16: 896-903.
Direct Link - Ashraf, R. and T.A. Ali, 2007. Effect of heavy metals on soil microbial community and mung beans seed germination. Pak. J. Bot., 39: 629-636.
Direct Link - Bandana, B., K.S. Anaytullah, A.K. Srivastava, S.K. Kuril and P.K. Singh, 2008. Effect of mercuric chloride on seed germination, seedling growth and enzyme activities in maize (Zea mays L.). Indian J. Plant Physiol., 13: 284-290.
Direct Link - Bhanumathi, P. and N. Jayabalan, 2007. Phytotoxic effects of mercuric acetate on peanut (Arachis hypogaea L.). Adv. Plant Sci., 20: 121-124.
Direct Link - Bonifacio, R.S. and M.N.E. Montano, 1998. Inhibitory effects of mercury and cadmium on seed germination of Enhalus acoroides (L.f.) Royle. Bull. Environ. Contamination Toxicol., 60: 45-51.
CrossRef - Carpi, A., S.E. Lindberg, E.M. Prestho and N.S. Bloom, 1997. Methyl mercury contamination and emission to the atmosphere from soil amended with municipal sewage sludge. J. Environ. Q., 26: 1650-1655.
Direct Link - Devi, P.U., S. Murugan, S. Akilapriyadharasini, S. Suja and P. Chinnaswamy, 2007. Effect of mercury and effluents on seed germination, root-shoot length, amylase activity and phenolic compounds in Vigna unguiculata. Nat. Environ. Pollut. Technol., 6: 457-462.
Direct Link - Emese, S., A. Rita, G. Katalin and M.G. Gabriella, 2009. Change of bioaccumulation of toxic metals in vegetables. Commun. Soil Sci. Plant Anal., 40: 285-293.
CrossRef - Fargasova, A., 1994. Effect of Pb, Cd, Hg, As and Cr on germination and root growth of Sinapis alba seeds. Bull. Environ. Contam. Toxicol., 52: 452-456.
CrossRefDirect Link - Garcia, E.M. and R.E. Reyes, 2001. Synthesis pattern of an Hg-binding protein in Acetabularia calyculus during short-term exposure to mercury. Bull. Environ. Contamination Toxicol., 66: 357-364.
Direct Link - Sengar, R.S., M. Gautam, S.K. Garg, R. Chaudhary and K. Sengar, 2008. Effect of lead on seed germination, seedling growth, chlorophyll content and nitrate reductase activity in mung bean (Vigna radiata). Res. J. Phytochem., 2: 61-68.
CrossRefDirect Link - Hussain, M., G. Yasin, A. Ali and R. Ahmed, 2007. Amelioration of toxic effects of lead (Pb) and chromium (Cr) in four black gram (Vigna mungo L.) Hepper cultivars with the application of kinetin. Pak. J. Agric. Sci., 44: 251-258.
Direct Link - Jain, R.K., H. Kumar and N.P. Saxena, 1998. Effects of cobalt and mercury on seedling vigour in Brassica campestris var. toria P. T. 303. J. Indian Botanical Soci., 77: 13-18.
Direct Link - Jeliazkova, E.A. and L.E. Craker, 2003. Seed germination of some medicinal and aromatic plants in a heavy metal environment. J. Herbs Spices Med. Plants, 10: 105-112.
CrossRef - Jeliazkova, E., L.E. Craker and B.S. Xing, 2003. Seed germination of anise, caraway, and fennel in heavy metal contaminated solutions. J. Herbs Spices Med. Plants, 10: 83-93.
CrossRef - Jun, R., T. Ling and Z. Guanghua, 2009. Effects of chromium on seed gemination, root elongation and coleoptile growth in six pulses. Int. J. Environ. Sci. Technol., 6: 571-578.
Direct Link - Leon, V., J. Rabier, R. Notonier, R. Barthelemy and X. Moreau et al., 2005. Effects of three nickel salts on germinating seeds of Grevillea exul var. rubiginosa, an endemic serpentine Proteaceae. Ann. Bot., 95: 609-618.
CrossRef - Li, W., M.A. Khan, S. Yamaguchi and Y. Kamiya, 2005. Effects of heavy metals on seed germination and early seedling growth of Arabidopsis thaliana. Plant Growth Regul., 46: 45-50.
CrossRef - Liu, W.X., H.H. Li, S.R. Li and Y.W. Wang, 2006. Heavy metal accumulation of edible vegetables cultivated in agricultural soil in the suburb of Zhengzhou city, peoples republic of China. Bull. Environ. Contamination Toxicol., 76: 163-170.
CrossRef - Mahmood, T., K.R. Islam and S. Muhammad, 2007. Toxic effects of heavy metals on early growth and tolerance of cereal crops. Pak. J. Bot., 39: 451-462.
Direct Link - Marchiol, L., S. Assolari, G. Pellet and G. Zerbi, 2006. Germination and seedling growth of Indian mustard exposed to cadmium and chromium. Ital. J. Agron., 1: 45-49.
Direct Link - Mishra, A. and M.A. Choudhuri, 1997. Ameliorating effects of salicylic acid on lead and mercury-induced inhibition of germination and early seedling growth of two rice cultivars. Seed Sci. Technol., 25: 263-270.
Direct Link - Mishra, A. and M.A. Choudhuri, 1997. Differential effects of Pb+2 and Hg+2 on inhibition of germination of seeds of two rice cultivars. Indian J. Plant Physiol., 2: 41-44.
Direct Link - Mishra, A. and M.A. Choudhuri, 1998. Amelioration of lead and mercury effect on germination and rice seedling growth by antioxidants. Biol. Plant., 41: 469-473.
CrossRef - Mishra, A. and M.A. Choudhuri, 1999. Monitoring of phytotoxicity of lead and mercury from germination and early seedling growth indices in two rice cultivars. Water Air Soil Pollut., 114: 339-346.
CrossRef - Munzuroglu, O. and H. Geckil, 2002. Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Arch. Environ. Contam. Toxicol., 43: 203-213.
CrossRefDirect Link - Neelima, P. and K.J. Reddy, 2003. Differential effect of cadmium and mercury on growth and metabolism of Solanum melongena L. seedlings. J. Environ. Biol., 24: 453-460.
PubMed - Nriagu, J.O., 1989. A global assessment of natural sources of atmospheric trace metals. Nature, 338: 47-49.
CrossRefDirect Link - Parmar, N.G., S.D. Vithalani and S.V. Chanda, 2002. Alteration in growth and peroxidase activity by heavy metals in Phaseolus seedling. Acta Physiol. Plant., 24: 89-95.
Direct Link - Patra, M., N. Bhowmik, B. Bandopadhyay and A. Sharma, 2004. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot., 52: 199-223.
CrossRefDirect Link - Patra, M. and A. Sharma, 2000. Mercury toxicity in plants. Botanical Rev., 66: 379-422.
CrossRefDirect Link - Peralta, J.R., J.L. Gardea-Torresdey, K.J. Tiemann, E. Gomez, S. Arteaga, E. Rascon and J.G. Parsons, 2001. Uptake and effects of five heavy metals on seed germination and plant growth in Alfalfa (Medicago sativa L.). Bull. Environ. Contam. Toxicol., 66: 727-734.
CrossRefDirect Link - Shafiq, M. and M.Z. Iqbal, 2005. The toxicity effects of heavy metals on germination and seedling growth of Cassia siamea Lamk. J. New Seeds, 7: 95-105.
CrossRef - Sharma, S.S., 1985. Effect of mercury on germination and seedling growth, mobilization of food reserves and activity of hydrolytic enzymes in Pisum sativum. Environ. Exp. Botany, 25: 189-193.
CrossRef - Shaukat, S.S., M. Mushtaq and Z.S. Siddiqui, 1999. Effect of cadmium, chromium and lead on seed germination, early seedling growth and phenolic contents of Parkinsonia aculeata L. and Pennisetum americanum (L.) Schumann. Pak. J. Biol. Sci., 2: 1307-1313.
CrossRefDirect Link - Street, R.A., M.G. Kulkarni, W.A. Stirk, C. Southway and J. van Staden, 2007. Toxicity of metal elements on germination and seedling growth of widely used medicinal plants belonging to Hyacinthaceae. Bull. Environ. Contam. Toxicol., 79: 371-376.
CrossRefDirect Link - Umadevi, P., N. Kannikaparameswari, S. Selvi and S. Murugan, 2009. Effect of mercury exposure on Vigna unguiculata (cowpea) seeds. Nat. Environ. Pollut. Technol., 8: 53-58.
Direct Link - Xiong, Z.T., 1998. Lead uptake and effects on seed germination and plant growth in a Pb hyperaccumulator Brassica pekinensis Rupr. Bull. Environ. Contam. Toxicol., 60: 285-291.
CrossRefDirect Link