
J.P. Pandey
Laboratory of Silkworm Physiology, Central Tasar Research and Training Institute, Central Silk Board, P.O. Piska-Nagri, Ranchi-835003, India
P.K. Mishra
Laboratory of Silkworm Physiology, Central Tasar Research and Training Institute, Central Silk Board, P.O. Piska-Nagri, Ranchi-835003, India
D. Kumar
Laboratory of Silkworm Physiology, Central Tasar Research and Training Institute, Central Silk Board, P.O. Piska-Nagri, Ranchi-835003, India
B.M.K. Singh
Laboratory of Silkworm Physiology, Central Tasar Research and Training Institute, Central Silk Board, P.O. Piska-Nagri, Ranchi-835003, India
B.C. Prasad
Laboratory of Silkworm Physiology, Central Tasar Research and Training Institute, Central Silk Board, P.O. Piska-Nagri, Ranchi-835003, India
Research Journal of Immunology
Year: 2010 | Volume: 3 | Issue: 2 | Page No.: 169-177







ABSTRACT
Insect blood cells (Hemocytes) are very vital component of the insect immune system but elaborate studies on hemocyte profile of economically very important sericigenous insect A. mylitta under altered physical conditions were unnoticed by researchers. Hence, an attempt was made to investigate the effects of high, low and cyclic temperature treatment/stress on cellular (hemocyte) immune responses such as Total Hemocyte Count (THC), Differential Heamocytes Count (DHC) and related changes in cell contour/morphology in fifth instar larvae of A. mylitta. Based on morphology staining reaction and response towards temperature stress six types of hemocytes were identified in A. mylitta. They are prohemocytes (PRs), plasmatocytes (PLs), granulocytes (GRs), spherulocytes (SPs), oenocytoids (OEs) and adipocytes (ADs). Besides, vermicytes (Ves) and podocytes (POs) were occasionally observed in smears of moulting phase of late fifth instar larvae. Presence of POs is unique in the present insect and it has not been reported earlier. Variations in Differential Hemocyte Counts (DHC) were observed when the ambient regime of temperature 26±1°C was changed to Chilling (4°C), heating (50°C) and cyclic 20, 26 and 32°C to cause temperature stress. Reduction in PLs and GRs were observed during Cyclic Temperature Treatment (CTT) in comparison to normal temperature but chilling and heating temperature did not showed the similar trends. Decrease in PLs and increase in GRs were observed after chilling temperature and increase in PLs and decrease in GRs were noticed after heating temperature. More mitotically dividing PRs were seen during elevated temperature stress. Low temperature lead to clumping of hemocytes and high temperature to its spread and nuclear fragmentation. Significant drop in THC was observed at low temperature but surprisingly at heating temperature significant increase was noticed. THC was found to decline in 120 h CTT larvae.
PDF Abstract XML References Citation
How to cite this article
J.P. Pandey, P.K. Mishra, D. Kumar, B.M.K. Singh and B.C. Prasad, 2010. Effect of Temperature on Hemocytic Immune Responses of Tropical Tasar Silkworm, Antheraea mylitta D. Research Journal of Immunology, 3: 169-177.
DOI: 10.3923/rji.2010.169.177
URL: https://scialert.net/abstract/?doi=rji.2010.169.177
DOI: 10.3923/rji.2010.169.177
URL: https://scialert.net/abstract/?doi=rji.2010.169.177
INTRODUCTION
Hemocytes are very vital component of the insect immune system and are biochemically very sensitive having multiple functions such as nodule formation, phagocytosis and encapsulation as defense mechanism; synthesis and transport of nutrients and hormones for proper growth and wound healing by way of connective tissue formation (Mangalika et al., 2010). Hemocytes are also reported as a source of storage protein/ hexamerins. Most of the studies in Antheraea mylitta hemocytes have been made to count them in different instars and generations without much correlation with defense, growths and transport mechanisms. Elaborate studies on hemocyte profile of economically very important sericigenous insect A. mylitta under altered physical conditions were unnoticed by researchers.
A close link exists between insect development and temperature. In a number of insect species, the growth rate is increased and the developmental period is shortened as the temperature increases (Wigglesworth, 1972; Kiuchi et al., 2008). However, each species has its own range of temperatures for its normal growth and very high temperature slows down the growth, may leads to developmental failure, such as larval ecdysis and adult emergence (Chapman, 2002). The exact mechanisms underlying these effects are not well understood. In this study, we sought to record the effects of high, low and cyclic temperature on A. mylitta at hemocytic level. The insect blood cells (hemocytes) are known to play a key role in developmental processes during which they disassociate from tissues and reconstruct metamorphosing tissues. The studies conducted on hemocyte counts in different insects by various authors did not show any concurrence on the effect of temperature. Low temperature treatment to insect’s decreases THC (Tauber and Yeager, 1935; Tiwari and Shukla, 2000) and increase in THC at high temperature have been noticed (Tauber and Yeager, 1935; Rosenberger and Jones, 1960). In some cases no change in THC at low temperature has been reported (Arnold, 1952; Rosenberger and Jones, 1960). In addition, very little and contradictory information are available on effects of temperature on DHC (Arnold, 1952; Behera et al., 1999; Pandey et al., 2003). However, no such report is available in tropical tasar silkworm, A. mylitta. Hence, an attempt was made to investigate the effects of high, low and cyclic temperature treatment/stress on cellular (hemocyte) immune responses such as THC, DHC and related changes in cell contour/morphology in fifth instar larvae of A. mylitta.
MATERIALS AND METHODS
Insect Culture
Stock of tropical tasar silkworm, Antheraea mylitta Drury (Daba ecorace) was maintained in indoor condition at 26±1°C, 60±5% relative humidity with photoperiod exposure of 14:10 h light: dark in Vista Biocell environmental chamber throughout the fifth instar larvae as control. Works were carried out during the Year 2009 at Silkworm Physiology Laboratory, Central Tasar Research and Training Institute, P.O. Piska-Nagri, Ranchi 835303, India. Larvae were fed on fresh leaves of Asan, Terminalia tomentosa W and A.
Cyclic Temperature Treatment (CTT)
To create near outdoor fluctuating temperature condition as experienced during tasar silkworm rearing in rainy season, Vinstar larvae were treated with 20, 26 and 32°C in cyclic phase of 8 h each (20°C for 8 h, 26°C for 8 h and 32°C for 8 h during each day of V instar). The CTT were given in Vista Biocell environmental chamber and hemolymph smears were taken from 120 h old V instar larvae for observations. THC and DHC were observed, separately for control and temperature treated larvae.
Chilling and Heating
The 24 and 120 h old fifth instar larvae were exposed separately to a chilling temperature of 4-6°C for 1 h by placing them directly, on ice cubes in a refrigerator and heating temperature of 50°C for 1 h in incubator. THC and DHC were recorded by taking hemolymph smears of 120 h old treated larvae as 24 h old larvae under heating treatment (1 h 50°C) could not survive.
Smear Preparation and Staining
A. mylitta hemolymph samples were obtained from the desired stage. A drop of fresh hemolymph was collected by puncturing the prolegs (with the help of 70% ethanol sterilized needle) on a slide and mixed well with anti-coagulant. A thin uniform smear of hemolymph was spread on the slide by rubbing the edge of an inclined slide backward. Stock solution of Giemsa stain was prepared as per standard protocol (Tauber and Yeager, 1935). A portion of it was diluted 10 times with double distilled water (DDW). The air-dried smear was stained for 20 min and thereafter rinsed with DDW. Hemocytes were categorized based on morphology, staining reaction and response towards temperature as observed under light and phase contrast microscopy. The methods of smear formation, staining and hemocyte categorization were similar to those applied earlier (Pandey and Tiwari, 2004, 2005).
Total and Differential Hemocyte Count (THC and DHC)
For THC, the hemolymph was drawn into a thoma blood-cell pipette up to its graduated mark of 0.5 and diluted up to the 11th mark with Tauber-Yeager’s fluid (Tauber and Yeager, 1935). The pipette was then shaken for several minutes and the first three drops were discarded. A double line with improved Neubauer ruling haemocytometer was filled with diluted hemolymph and the hemocytes were counted at four corners and one central (1 mm2) square. When the distribution of cells in all the squares was not even, the sample was discarded and the procedure was repeated. The number of circulating hemocytes per cubic millimeter (mm3) was calculated using the formula of Jones (1962). Ten to 12 hemolymph determinations, each form oozing cut prolegs of 4-5 larvae were drawn and diluted and hemocytes were counted on a standard blood-cell counter under phase-contrast microscope. Permanent preparations of hemolymph smear, staining and calculation of relative percentage of different hemocyte types were similar to those adopted earlier (Pandey and Tiwari, 2004). DHC was conducted by counting of different categories of randomly selected 200 cells from stained smears of 10 insects was done both for treated and control larvae. The percentage of different cell types was calculated. The data were subjected to the statistical analysis by using Student’s t-test.
RESULTS
Observations on hemocyte categories and their THC, DHC, cell contours during V instar larvae were taken with the help of light and phase contrast microscopy. Based on morphology, staining reaction and response towards temperature stress, six types of hemocytes have been recognized in V instar larvae. They are PRs, PLs, GRs,SPs, ADs and OEs. The VEs and POs were occasionally observed in smears of moulting phase of late fifth instar larvae (Fig. 1).
Effect of Temperature Treatment on THC and DHC
Low temperature (4°C) and CTT caused a decrease in THC. These counts were always lower than control (Table 1). The total count of hemocytes declined to about 37% when larvae were subjected to low temperature at the age of 120 h. Similar declining trend of THC was observed in larvae treated with CTT (Table 1).
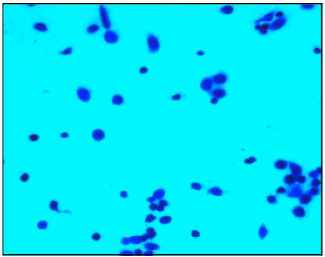 | |
| Fig. 1: | Hemolymph smear of A. mylitta showing different types of hemocytes x 450 |
| Table 1: | Changes in THC and DHC in V instar larvae of A. mylitta under diverse temperature regimes. (Values represent Mean±SD of 10-12 haemolymph determinations, each drawn from 10 larvae) |
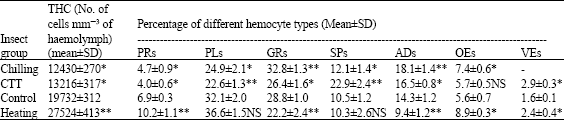 | |
| NS: Not Significant; p-values: *<0.05; **<0.01 | |
Contrary to this, heating of larvae at 50°C for an hour resulted into increased the THC by 1.3-1.5 folds. THC increased 19732±312 (control) to 27524±413 after heating. Variations in DHC were significant between three temperature treatments. The counts of PRs and PLs decreased under chilling treatments but a little rise in the number of GRs, SPs, ADs and OEs was recorded (Table 1). In heat-exposed larvae number of Prs, Pls and OEs rose while decline other cell types were observed (Table 1). Under CTT decrease in PRs, PLs and GRs percentages was observed but rise in the counts of GRs, SPs, ADs, OEs and VEs were recorded (Table 1). Short term heating (1 h at 50°C) showed a different pattern in relative percentage of various cell types. The heat exposure for a short period also induced cell damages under different conditions (cf Fig. 4-5).
Effect of Temperature Treatment on Hemocyte Morphology
Low temperature induced a little clumping of hemocytes (Fig. 2-3). Heating treatment caused damage to most of the cell types. Loss of cytoplasmic compactness of pseudopods in PLs, vacuolization in PLs and GRs fragmentations of nuclei including PRs, overall disintegration and cell death were some of the features affecting contour of the cells under heat-exposed larvae (1 h 50° C).
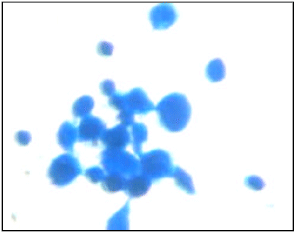
CTT induced damage of PLs and GRs, loss of cytoplasmic compactness of pseudopods in PLs, vacuolization in PLs and GRs (Fig. 6-7).
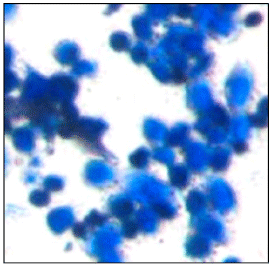 | |
| Fig. 2: | Low temperature induces the clumping of hemocytes in A. mylitta larvae x 675 |
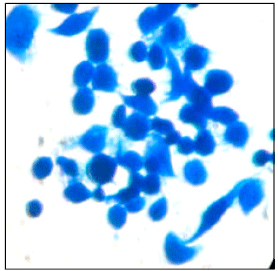 | |
| Fig. 3: | Hemocytes profile of control A. mylitta larvae x 675 |
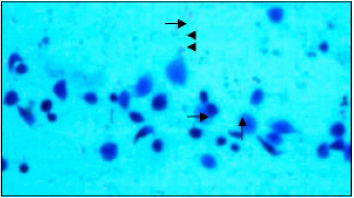 | |
| Fig. 4: | Loss of cytoplasmic compactness and pseudopods in hemocytes of A. mylitta after exposure to heating temperature x 675 |
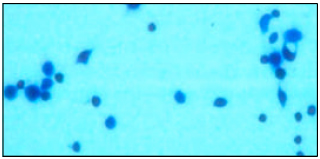 | |
| Fig. 5: | Hemocytes profile of control A. mylitta larvae x 675 |
 | |
| Fig. 6: | CTT induces the changes in hemocyte contour of A. mylitta larvae x 675 |
 | |
| Fig. 7: | Hemocytes profile of control A. mylitta larvae x 675 |
DISCUSSION
As accounts available on hemocytes on the whole are enormous, but most of the studies regarding hemocyte categories PRs, PLs, GRs,SPs, ADs, OEs, VEs and POs) amongst insects are not unanimous and are confined to one or two family (Pandey et al., 2003; Pandey and Tiwari, 2005). In present study, the change in pattern of THC in A. mylitta was observed under different temperature treatment. Variation in hemocytes under altered physiological conditions has been investigated by a number of researchers (Lackie, 1988; Gardiner and Strand, 2000; Pandey et al., 2003; Kiuchi et al., 2008). Cell division in insects depends on the developmental stage of the individual and is strongly regulated by the hemolymph concentration of hormones like 20-hydroxyecdysone (20-HE) and Juvenile Hormone (JH) particularly on the cell cycle and wing discs differentiations (Koyama et al., 2004; Kiuchi et al., 2008). It is reported that JH suppresses the growth and development of imaginal tissues after ecdysis in starved Manduca sexta larvae (Truman et al., 2006). Jones (1967) found an elevation in THC during larval development of G. mellonella in both heat-fixed and unfixed larvae though; the counts were significantly higher in heat-fixed insects. He has suggested that this increase in cell count is possibly due to loss of body fluid as a result of desiccation. Enhanced mitotic rate of Blaberus hemocytes exposed to 37°C was also reported by Tauber and Yeager (1935). However, the increase in THC in P. demoleus larvae after heating has been ascribed to more mitotic index which may be attributed to higher rate of synthesis and release of prothoracicotropic hormone (PTTH) from the brain and of ecdysone from prothoracic glands (PTG) (Tiwari and Shukla, 2000) as these physical factors are realized directly into the neurosecretory cells (NSCs) of the brain. The role of ecdysone or PTG in production, multiplication and differentiation of hemocytes has been shown earlier by a number of workers (Rao et al., 1984; Tiwari and Shukla, 2000). A comparable interpretation seems to be relevant for increased THC following high temperature in A. mylitta larvae. As defense mechanism of heat exposed A. mylitta larvae, an increase in number of hemocytes may be attributed possibly due to the detachment of hemocytes from tissue surfaces and higher rate of multiplication or production of hemocytes. The comparative reduction in THC in A. mylitta following chilling temperature treatment further supports the views of Tiwari and Shukla (2000) in P. demoleus. The decline in THC may be attributed to the clumping of the cells induced by chilling stress and thus rendering the hemocytes unavailable in circulating hemolymph. Clumping of hemocytes may be taken as a physiological response of hemocyte in transport system. In view of the fact that insects are cold-blooded-nature the low temperature might induce the clumping of hemocytes. The study of hemolymph cells in A. mylitta showed that the PLs and GRs are the only cell type which always gets affected at diverse temperature regimes. PLs and GRs were noticed to decrease under CTT with simultaneous increase in SPs. Trends of DHC showed that count of PLs, GRs and SPs were closely related to each other. The count of PRs declined in chilled and CTT, whereas it increased at high temperature. This shows that the PRs undergo mitotic divisions at high temperature. The stem-cell nature of PRs has been reported earlier by a number of workers. The variations in counts of other cell types are dependent to various physiological functions attributed to them as described in detail by Pandey and Tiwari (2005) in D. chrysippus larvae. The structural changes in haemolymph cells leading to their disintegration have been observed in A. mylitta larvae under stress conditions, corroborate the findings of Pandey et al. (2003).
In conclusion, the effects of temperature stress on hemocytes of A. mylitta studied perhaps for the first time. To understand the inhibitory effects of high and low temperature on insect growth at the cellular level, we investigated the influence of different temperature regimes on the larval hemocytes in the tasar silkworm, A. mylitta. Chilling caused a decline in the number of hemocytes, heating elicited an increase in their number. The relative percentage of different hemocyte types showed higher variation under different treatments and they have interdependence also. In addition temperature stress badly affected the cell contour also. The present study could only deal with the effect of temperature on THC, DHC and cell contour in order to initiate studies on establishing the immune response at cellular level in one of the most important commercial insect. Further studies are required to establish a correlation between temperature, hormonal status and hemocyte profile in order to investigate further in immune system of the insects.
ACKNOWLEDGMENTS
JP is thankful to Prof. R. K. Tiwari, Principal, K.N. Govt. P.G. College, Gyanpur-221304, India for vital suggestion and procedural support.
REFERENCES
- Jones, J.C., 1962. Current concepts concerning insect hemocytes. Am. Zool., 2: 209-246.
CrossRefDirect Link - Kiuchi, T., F. Aoki and M. Nagata, 2008. Effects of high temperature on the hemocyte cell cycle in silkworm larvae. J. Insect Physiol., 54: 454-461.
CrossRefDirect Link - Koyama, T., M. Iwami and S. Sakurai, 2004. Ecdysteroid control of cell cycle and cellular commitment in insect wing imaginal discs. Mol. Cell. Endocrinol., 213: 155-166.
Direct Link - Pandey, J.P. and R.K. Tiwari, 2005. Feeding, brain implantation and 20-hydroxyecdysone treatment reverses the effect of starvation and ventral nerve cord severance on haemocyte counts in larvae of the plain tiger butterfly Danaus chrysippus (Lepidoptera: Nymphalidae). Int. J. Trop. Insect Sci., 25: 295-300.
CrossRefDirect Link - Rao, C.G.P., A. Ray and P.S. Ramamurty, 1984. Effect of ligation and ecdysone on total haemocyte count in the tobacco caterpillar, Spodoptera litura (Noctuidae: Lepidoptera). Can. J. Zool., 62: 1461-1463.
CrossRefDirect Link - Rosenberger, C.R. and J.C. Jones, 1960. Studies on the total blood cell counts of the southern armyworm larvae, Prodenia eridania (Lepidoptera). Ann. Entomol. Soc. Am., 53: 351-355.
CrossRefDirect Link - Mangalika, P.R., T. Kawamoto, A. Takahashi-Nakaguchi and K. Iwabuchi, 2010. Characterization of cell clusters in larval hemolymph of the cabbage armyworm Mamestra brassicae and their role in maintenance of hemocytepopulations. J. Insect Physiol., 56: 314-323.
CrossRef - Tauber, O.E. and J.F. Yeager, 1935. On total hemolymph (blood) cell counts of insects I. Orthoptera, odonata, hemiptera and homoptera. Ann. Entomol. Soc. Am., 28: 229-240.
CrossRefDirect Link - Truman, J.W., K. Hiruma, J.P. Allee, S.G. Macwhinnie, D.T. Champlin and L.M. Riddiford, 2006. Juvenile hormone is required to couple imaginal disc formation with nutrition in insects. Science, 312: 1385-1388.
CrossRefPubMedDirect Link