
Rajesh Kumar
Rain Forest Research Institute, P.B.-136, Jorhat, Assam, India
Ashwani Tapwal
Rain Forest Research Institute, P.B.-136, Jorhat, Assam, India
Debu Moni Baruah
Rain Forest Research Institute, P.B.-136, Jorhat, Assam, India
Research Journal of Forestry
Year: 2012 | Volume: 6 | Issue: 1 | Page No.: 24-31







ABSTRACT
Soil is the major reservoir of C in the terrestrial environment and always under dynamic equilibrium due to various bio-geochemical processes active soil development. Litter accumulate forest floor and slowly decompose by the microbial activity to release the nutrients into the surrounding environment. Carbon dioxide is released from the soil to the atmosphere in heterotrophic respiration when the dead organic matter is used for substrates for soil micro-organisms and soil animals. The climatic factors such as rainfall, temperature and seasonal variations directly influence the occurrence and abundance of microbes that results in the variation of rate of leaf litter decomposition. This work was focused on variation in the rate of leaf litter decomposition in different seasons of the year of two plant species: Dipterocarpus tuberculatus and Dipterocarpus retusus at two different sites and seasonal variation in the microfungal quantity of the decomposed leaf litter was also enumerated. The results revealed a similar trend for the rate of decomposition for both species and a significant correlation between the abiotic factors and leaf litter decomposition.
PDF Abstract XML References Citation
Received: August 16, 2011;
Accepted: November 23, 2011;
Published: February 14, 2012
How to cite this article
Rajesh Kumar, Ashwani Tapwal and Debu Moni Baruah, 2012. Leaf Litter Decomposition Pattern in Dipterocarpus tuberculatus and Dipterocarpus retusus Forests of North East India. Research Journal of Forestry, 6: 24-31.
URL: https://scialert.net/abstract/?doi=rjf.2012.24.31
URL: https://scialert.net/abstract/?doi=rjf.2012.24.31
INTRODUCTION
Forest soils host large amounts of tree roots. The amount of C incorporated in the tree roots, accounts between 14 and 27 tonnes C ha-1 (Brunner and Godbold, 2007). Forests play an important role in the global carbon (C) cycle and management as well as climate can cause major effects on the balance of C between the atmosphere and the plant/soil system (Klemedtsson et al., 2010). In forests, roughly half of the soil respiration is autotrophic (mainly root respiration) while the remainder is heterotrophic, originating from decomposition of soil organic matter. Tree species induced changes may have a great impact on atmospheric CO2 concentrations (Berger, 2010). Carbon accumulation in soil is an important process of the carbon cycle in forest ecosystems. Anthropogenic disturbances could quickly affect soil carbon stocks and dynamics, especially where organic substances drive the soil-forming processes, such as in Podzols (Falsone et al., 2010).
Litter decomposition is one of the key biogeochemical processes in forest ecosystems (Swift et al., 1979). The rate of litter decomposition is largely a determining factor for productivity of forest ecosystems as plant nutrients became available for recycling within the system during the process. It is estimated that the nutrients released during litter decomposition can account for 69-87% of the total annual requirement of essential elements for forest plants (Waring and Schlesinger, 1985).
Litter cover also acts as a protective layer by buffering changes in soil water content (Ginter et al., 1979) and temperature (MacKinney, 1929) and hindering erosion (Lowdermilk, 1930), leaching (Lunt, 1951; Mo et al., 2003) and soil compaction (Benkobi et al., 1993). Litter decomposition is influenced by environmental factors and physiochemical properties of the parts such as stem wood, leaves, root, etc. of the species studied and the decomposer organisms present in the soil (Vesterdal, 1999; Wedderburn et al., 1999). Three factors which are site environmental condition (particularly climate), litter quality and the soil biota are known to control litter decomposition processes (Swift et al., 1979). Under certain circumstance, litter incorporation to the soil may reduce nutrients cycling and plant growth depending on the soil and litter chemical properties (Seneviratne, 2002). Once the incorporated litter was encapsulated forming humus, it contributes to increased C sequestration in the soil (Seneviratne, 2003). Allen et al. (2005) observed a higher net N mineralization in the soil when there was a below-ground interaction of the tree-crop system. Climate is known to be the dominant factor influencing litter decomposition on a large geographical scale or under unfavorable climatic conditions (Meentemeyer, 1978; Dyer et al., 1990; Austin and Vitousek, 2000). Therefore, litter decomposition in wet tropical regions is considered to be more rapid than in temperate regions, although decomposition rates vary widely in both climatic regions (Anderson and Swift, 1983; Aerts, 1997; Takeda, 1998). In contrast, under favorable climatic conditions, litter quality becomes a more important factor in determining litter decomposition processes (Bargali et al., 1993; Austin and Vitousek, 2000). Litter microbes are important components of litter decomposition. They act at various levels of litter decomposition. Fungi are the most important decomposer of plant material and 75% of the decomposition of various organic substrates is done mainly by different fungal species (Kjoller and Struwe, 1992). Population of microfungi varies in different seasons of the year (Behera and Mukerji, 1985). In addition to fungi bacteria are also responsible for the leaf litter decomposition (Mahasneh, 2001). In present study, variation in the rate of leaf litter decomposition in different seasons of the year of two plant species at two different sites, viz- Dipterocarpus tuberculatus near Moreh Town, Chandel District, Manipur and Dipterocarpus retusus at Deomali, Arunachal Pradesh were evaluated. Seasonal variation in the microfungal quantity present in the decomposed leaf litter was also enumerated. Dipterocarpus is a good timber yielding plant and it was selected for the study as the rate of decomposition of Dipterocarpus leaf litter is comparatively faster. Devi and Yadava (2007) reported that only 8.4% of the leaf litter left at the end of 12 months and complete decomposition of Dipterocarpus tuberculatus leaf litter occurred in 15 months.
The study was conducted in two different locations. The study site one is a South Bank Tropical Wet Evergreen Dipterocarpus retusus Forest (Champion and Seth, 1968) Tirap District, Deomali reserve forest, Arunachal Pradesh. Its altitude ranged between 150-600 msl and the mean temperature ranges between 22-36°C in summer and 10-25°C in winter. The rainfall starts with pre-monsoon in March and followed by the onset of Monsoon in May to its withdrawal during September (3/4th rainfall occurs during this period). The retreating monsoon and North East Monsoon also causes some rains in between. The total rainfall in this belt varies 2500-3500 mm and virtually having no dry month. The study site two is a tropical deciduous forest (Champion and Seth, 1968) located near Moreh town, Chandel District, between 23, 49’-24°28’N lat. and 94°09’- 94°31’ E long E long. in Manipur along the Indo-Myanmar border of Northeast India at 300-360 msl. There are three seasons-spring (March to May), rainy (June to October) and winter (November to February). The mean monthly maximum and minimum temperature ranged from 24.2 (January) to 36°C (May) and from 5.0 (January) to 23°C (August), respectively. The area receives 1245 mm annual rainfall, of which more than 80% occurs during the rainy season.
MATERIALS AND METHODS
The study was conducted from April 2008 to July 2009. To determine remaining weight of leaf litter, 120 (24 bags x 5 replicates) nylon bags of 20x20 cm area and 2 mm mesh size were prepared for each plant species. Freshly fallen leaves of the Dipterocarpus sp. were collected and air dried to get a constant weight of 10 g. These were placed in the bags and pinned to the forest floor with metal pins in 5 sub-plots each having 12x12 m area. Five bags were harvested every month (one bag from each sub-plot) until there was complete decomposition. Five more litter bags were placed in the same experimental sites (one bag in each sub-plot) every month and picked up the next month to study the weight loss rates per month.
Plot design of decomposition experiment: The extraneous particles were removed from the harvested litter bags by brushing them carefully and washed using tap water followed by distilled water with gentle agitation on a 1 mm mesh screen and oven dried at 60°C to get a constant weight. The weight loss%, the remaining weight% and the decomposition constant (k) were calculated following by Olson (1963).
Where:
| X0 | = | Weight of litter at time 0 |
| Xt | = | Weight of litter at time t |
| t | = | Time (usually in years) |
| k | = | Decomposition rate constant |
Every month, the fungal quantities were enumerated from decomposing leaf litter samples from each plot (5 sub-plots for each species). For this, samples were collected sterile polyethylene bags and brought back to laboratory in a portable icechest and stored at 4°C for further analysis. Serial dilution and plate count method was used to determine number of fungal colonies per g of dry leaf litter on Potato Dextrose Agar (PDA) medium at 25±3°C.
Total population of fungi per g of dry leaf litter was calculated using the following formula:
RESULTS AND DISCUSSION
Observations on litter decomposition for Dipterocarpus retusus and Dipterocarpus tuberculatus were recorded from April 2008-July 2009 and presented in Table 1 and Table 2. The results reveal almost similar pattern of decomposition in both species. A high value of k indicates that rate of decomposition of leaf litter of both the species is high. After 15th months, only 0.2% weight remained in case of Dipterocarpus tuberculatus and 0.4% weight remained in case of Dipterocarpus retusus. Similar findings were reported by Devi and Yadava (2007) in case of D. tuberculatus. Monthly variations in remaining litter mass were observed to be almost similar for both the species but little variation was observed in the month of September.
| Table 1: | Decomposition constant (k) value, percent mass loss, percent mass remaining and t values Dipterocarpus tuberculatus in year 2008-09 |
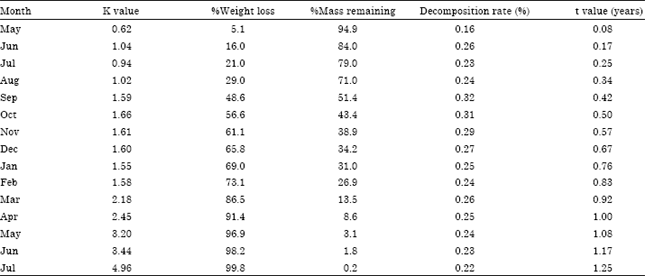 | |
| Table 2: | Decomposition constant (k) value, percent mass loss, percent mass remaining and t values Dipterocarpus retusus in year 2008-09 |
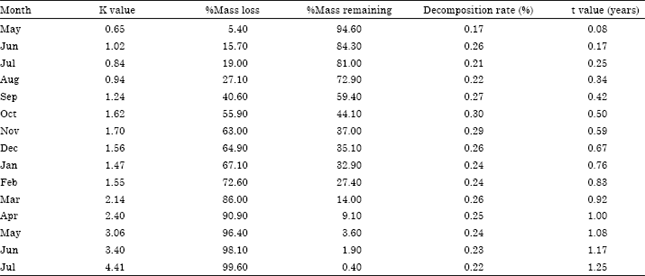 | |
Weight loss percentage of D. tuberculatus leaf litter between the months increased consistently and recorded maximum between July and September. Thereafter, it decreased and minimum percent mass loss is recorded in the month of November. Devi and Yadava (2007) also reported maximum loss of leaf litter in the months of August and September. Loss of D. retusus leaf litter was recorded maximum in August followed by July and minimum in November. For both the species weight loss percentage was recorded lower in winters (November to May). It is evident from both of the tables that loss of leaf litter was maximum during the rainy season (June to October) and considerably less in winter season (November to February) for both the species. Significant variations were observed in the mass loss percentage in different months and seasons for both the species. The difference in the rate of decomposition of the leaves of D. retusus and D. tuberculatus was negligible but the percent weight loss of D. tuberculatus leaf litter was slightly higher in the month of September. Results of t test revealed that the mass loss percentage of both the species correlates significantly.
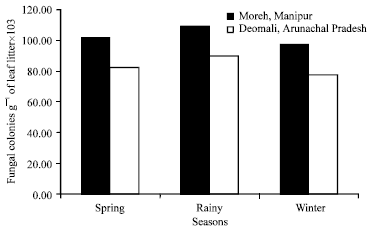 | |
| Fig. 1: | Average fungal colonies g-1 of leaf litter in different seasons from April 2008 to July 2009 |
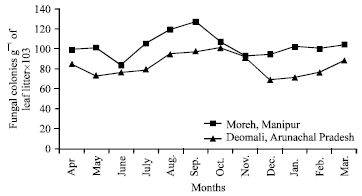 | |
| Fig. 2: | Seasonal variation in fungal colonies g-1 of leaf litter from April 2008-July 2009 |
In both species a significant variations were observed in the micro-fungal quantity in different months and seasons of the year. Maximum number of fungal colonies for both the species was observed in the rainy season and minimum in the winter season (Fig. 1). In case of D. tuberculatus the number of fungal colonies per gram of decomposed leaf litter was recorded maximum in the month of September and minimum in June, while for D. retusus it was maximum in October and minimum in January (Fig. 2). Higher rate of decomposition and number of fungal colonies were observed in rainy season. It may be due to leaching of water soluble nutrients from litter which became available for litter decomposing microfungi and other microorganisms. Three factors are known to control litter decomposition processes which are site environmental condition (particularly climate), litter quality and the soil biota (Swift et al., 1979). As a key factor of decomposition, higher number of fungal population may be a reason of faster decomposition of leaf litter. Significant correlation was observed between the abiotic factors and mass loss of leaf litter (Table 4). But the number of fungal colonies and the mass loss percent of the sampling months did not correlate statistically in Deomali, Arunachal Pradesh and significant correlation was observed in Moreh, Manipur (Table 3). Lesser weight loss and microfungal population may be due to cool and dry conditions. Moorhead et al. (1999) also reported that litter mass loss is higher in moist and warm environments than in cold or dry conditions. Variation in climatic factors alters the pattern of decomposition and occurrence of decomposer microbial populations. Compositional shifts in soil microbial communities, mediated by ecological interactions among soil saprotrophs, appear to lie at the biogeochemical heart of ecosystem response to environmental change.
| Table 3: | Correlation between rate of mass loss of leaf litter between both the species (n = 15) and number of fungal colonies with each species (n = 12) |
 | |
| *Not Significant, **Shows significance at p<0.05 level, ***Shows significance at p<0.01 level | |
| Table 4: | Correlation between rate of mass loss of leaf litter with soil moisture and rain fall |
 | |
| *Shows significance at p<0.05 level | |
Higher fungal population and high rate of decomposition has been recorded in warm and humid period of the year. Further investigations are required to determine the potent micro-fungi responsible leaf litter decomposition in a Dipterocarpus forest in a particular season.
ACKNOWLEDGMENTS
The authors are thankful to the Indian Council of Forestry Research and Education (ICFRE) for funding the research, project No: ICFRE/FP/13.
REFERENCES
- Mahasneh, A.M., 2001. Bacterial decomposition of Avicennia marina leaf litter from Al-khor (Qatar-Arabian Gulf). J. Biol., Sci., 1: 717-719.
CrossRefDirect Link - Allen, S.C., S. Jose, P.K.R. Nair, B.J. Brecke, V.D. Nair, D.A. Graetz and C.L. Ramsey, 2005. Nitrogen mineralization in a pecan (Carya illinoensis K. Koch)-cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Biol. Fertil. Soils, 41: 28-37.
Direct Link - Austin, A.T. and P.M. Vitousek, 2000. Precipitation decomposition and litter decomposability of Metrosideros polymorpha in native forests on Hawaii. J. Ecol., 88: 129-138.
CrossRef - Bargali, S.S., S.P. Singh and R.P. Singh, 1993. Patterns of weight loss and nutrient release from decomposing leaf litter in an age series of eucalypt plantations. Soil Biol. Biochem., 25: 1731-1738.
CrossRefDirect Link - Behera, N. and K.G. Mukerji, 1985. Seasonal variation and distribution of microfungi in forest soils of Delhi. Folia Geobot. Phytotax., 20: 291-311.
Direct Link - Brunner, I. and D.L. Godbold, 2007. Tree roots in a changing world. J. For. Res., 12: 78-82.
CrossRef - Ginter, D.L., K.W. McLeod and C. Sherrod Jr., 1979. Water stress in longleaf pine induced by litter removal. For. Ecol. Manage., 2: 13-20.
CrossRef - Lowdermilk, W.C., 1930. Influence of forest litter on run-off, percolation and erosion. J. For., 28: 474-491.
Direct Link - MacKinney, A.L., 1929. Effects of forest litter on soil temperature and soil freezing in autumn and winter. Ecology, 10: 312-321.
Direct Link - Meentemeyer, V., 1978. Macroclimate and lignin control of litter decomposition rates. Ecology, 59: 465-472.
Direct Link - Mo, J., S. Brown, S. Peng and G. Kong, 2003. Nitrogen availability in disturbed, rehabilitated and mature forests of tropical China. For. Ecol. Manage., 175: 573-583.
CrossRef - Moorhead, D.L., W.S. Currie, E.B. Rastetter, W.J. Parton and M.E. Harmon, 1999. Climate and litter quality controls on decomposition: An analysis of modeling approaches. Global Biogeochem. Cycles, 13: 575-589.
CrossRef - Olson, J.S., 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 44: 322-331.
CrossRefDirect Link - Devi, A.S. and P.S. Yadava, 2007. Wood and leaf litter decomposition of Dipterocarpus tuberculatus Roxb. in a tropical deciduous forest of Manipur, Northeast India. Curr. Sci., 93: 243-246.
Direct Link - Seneviratne, G., 2003. Global warming and terrestrial carbon sequestration. J. Biosci., 28: 653-655.
CrossRef - Wedderburn, M.E. and J. Carter, 1999. Litter decomposition by four functional tree types for use in silvopastoral systems. Soil Biol. Biochem., 31: 455-461.
CrossRef