
M. Faravani
Institute of Biological Sciences, University of Malaya, 50603, Kuala Lumpur, Malaysia
B.B. Baki
Institute of Biological Sciences, University of Malaya, 50603, Kuala Lumpur, Malaysia
Research Journal of Environmental Sciences
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 52-61







ABSTRACT
The effects of density on the growth rate and survival of individual plants as well as changes in population structure and self-thinning were experimentally evaluated in Melastoma malabathricum at five densities (19098, 76394, 152788, 229183 and 458365 seeds m-2). The biomass (dry weight) of root, stem and leaf was measured for seven times after drying from the 18 to 162th days after planting. It was applied at 20 day intervals. The relationship between the total dry matter weights per plant and plant densities of survivors for populations of Melastoma malabathricum showed that each population will start to thin along a line of slope from -3.7 to -1.2 until it reaches the maximum standing crop. Mortality during the phase of self-thinning is largely among individuals suppressed by the ensuing growth of neighbours, resulting in increased shading within the canopies of neighbouring plants. The total dry matter (m-2) was constant over a wide range of densities because individual plant displayed density-dependent reduction in growth rate and hence in individual plant size, in particular, because the reductions in mean plant weight compensated exactly for increase in density.
PDF Abstract XML References Citation
How to cite this article
M. Faravani and B.B. Baki, 2009. Population Structure and its Influence on Self-Thinning of Melastoma malabathricum L. Research Journal of Environmental Sciences, 3: 52-61.
DOI: 10.3923/rjes.2009.52.61
URL: https://scialert.net/abstract/?doi=rjes.2009.52.61
DOI: 10.3923/rjes.2009.52.61
URL: https://scialert.net/abstract/?doi=rjes.2009.52.61
INTRODUCTION
Thinning processes may occur in all crowded plant and animal populations, pure or mixed and this may play important roles in the demography of populations and community structure (Quinones et al., 2003). Intra-specific competition is a particular form of competition in which members of the same species vie for the same resource in an ecosystem (e.g., food, light, nutrients and space). This can be contrasted with inter-specific competition, in which different species compete (Solomon et al., 2002). In self-thinning of plant populations, size inequality decreases as a result of the predominant mortality of the smallest size class (Weiner et al., 2001; Rivera and Scrosati, 2008).
By understanding the factors limiting the growth of the individuals within the population, the response of the population as a whole can be deduced. This finding, termed the law of constant final yield (Polley et al., 2008) and this has been shown to hold for a large number of species. A widely accepted generalization about these paths is the self-thinning rule, or Power Law of Self-thinning (White and Harper, 1970; Lonsdale and Watkinson, 1982; Li, 2002; Simard and Zimonick, 2005), otherwise known as Self Thinning Law or the -3/2 Power Law or Yoda Law (Hara et al., 2006; Rivera and Scrosati, 2008; Wiegand et al., 2008).
The self-thinning rule relates plant mass to plant density in crowded, by a power-law equation with an exponent -3/2. It has been also called the -3/2 power law or Yoda`s law (Li et al., 2000; Rivera and Scrosati, 2008).
The model for self thinning was taken from Yoda et al. (1963) cited by Hara et al. (2006):
Log w = Log Κ + β Log Ï? |
where, w is the mean biomass per plant, where Ï? is the number of surviving plants, k and β are the self-thinning coefficient and exponent, respectively. The exponent β has been claimed to take the value -3/2 approximately regardless of species, age or site conditions and k varies with species and growth conditions and it is constant. The rule has been hotly debated whether a true self-thinning law exists in nearly pure stands of post fire chaparral and if so, is it a -3/2 or -4/3 law (Guo and Rundel, 1998; Zeide, 2001; Bi, 2004; McCarthy and Weetman, 2007). Lonsdale and Watkinson (1982) showed the -3/2 power law is a characteristic of shoots but not of whole plants. Recently, Chen et al. (2008) argued that the traditional slope of the upper boundary line, -3/2, has been challenged by -4/3 which is deduced from some new mechanical theories, like the metabolic theory.
More experimental or field studies should be carried out to identify the more accurate self-thinning exponent. In this study we wanted to check if the evidence could support acceptance of self-thinning rule as a quantitative biological law in Melastoma malabathricum. By using the slopes and intercepts of size-density relationships as variables, the slopes can be explained by simple geometric arguments. Here, we hypothesized that the sensitivity of plant may affect the rate of self-thinning, hence, in the plant community through resource capture and utilization.
MATERIALS AND METHODS
Plant Establishment
Seeds of M. malabathricum of the same cohorts were collected in September 2006 from the campus of the University of Malaya, Kuala Lumpur (3 ° 8` N; 101 ° 42` E) Malaysia. A synthetic community of M. malabthricum was established by sowing seeds directly in 26 cm depthx14 cm diameter in black plastic pots, previously filled with garden soil of Malacca series in an insect-proof house with 12 h of natural sunlight (mean midday radiation of 622 and 125 μmole photon m-2 sec-1 outdoor and inside in insect-proof house, respectively) and mean ambient temperatures of 33 ± 2 °C (day) and 25 ± 2 °C (night) at Rimba Ilmu, University of Malaya, Kuala Lumpur. The plants were watered once daily, in the morning from above with a fine rose. A seed germination test experiment was conducted to check seed vigour, seed viability and seed germination in advance, prior to experimentation. Five sowing densities viz. 19098, 76394, 152788, 229183 and 458365 seeds m-2, equal to 0.01, 0.040, 0.080, 0.12 and 0.240 g were applied in area each measuring 0.0154 m-2 and each density was accorded ten replicates.
Recording of Density and Individual Weight of Melastoma malabathricum
A circular quadrate was sampled in the center of each pot (using PVC pipe, internal diameter 3.0 cm) in the 40th (t1), 60th (t2), 80th (t3), 100th (t4) , 120th (t5),140th (t6) and 160th (t7) Days After Planting (DAP). The small quadrate (7.1 cm-2) was used in the populations of M. malabathricum with high density and the large quadrate (1.8 cm-2) in the populations with low density. The total No. of plants in the quadrate was recorded as number per m2. Samples were taken seven times on 18th to 162 days after planting (DAP) every 20 day intervals. All fresh plants were killed by liquid N2 to stop their metabolism. They were separated into stem, leaf and root before being dried for 48 h at 70 °C. After two weeks, the seed germination was considered as the initial densities in this study. Densities at this first observation were checked if they were less than the numbers we had expected to germinate, based on the weighed seed lots for each pot and on estimates of seeds pot-1 (Table 1). Therefore failures to germinate and mortality before emergence were not negligible. The mean seed germination rate at 25 °C was 40 in the laboratory tests. We have used principal component analysis with logarithmicized data to calculate the slope of the thinning lines shown in this study (Mohler et al., 1978).
RESULTS AND DISCUSSION
Mean density of established plants of M. malabathricum during the first measurement after germination was extremely lower than the initial sowing density in all density treatments. Substantial declines in density were observed with time by the first and ensuing harvests at the five density treatment over period of 160 days after sowing, the declines were density-dependent.
There was a strong, negatively density-dependent relationship with a linear relation y = 29.27- 3.56x, R2 = 0.94, p<0.05 for inhibition of germination by increasing sowing density (Fig. 1).
This negative relationship has been already reported in other plants by Lonsdale and Watkinson (1982). In these experiments, population numbers declined from the initial sowing densities not only as a result of self-thinning but also as a result of negatively density-dependent seed germination. At a sowing density of 458,365 seeds m-2, only 14% of the seeds germinated, in comparison with 30% at 19098 seeds m-2. There are, however, a number of reports of in most of these experiments, where overall density was held constant while seeds were aggregated in clumps of varying sizes. The CO2 produced by the roots might be acting as an inhibitor and also competition for resources between seeds and seedlings might be implicated (Inouye, 1980; Lonsdale and Watkinson, 1982).
As individuals grow in a competing population of plants, their mean biomass increases and their number decrease. Plotting mean plant biomass (dry weight, calculated by dividing total plant weight per pot by the number of survivors) (log) against the five density of survivors (plant m-2) in the populations with time of sowing showed that total plant weight decreased (Fig. 2, 3), starting in the lower right corner with a large number of plants of small size and moving up and to the left as the number of plants decline and the plants grow as a result of self thinning (Adler, 1996).
The relationship of the plant biomass and the plant density regimes were significant, in the regression models for self thinning (Table 1). It indicates the slope values and 95% confidence limits of slopes. The obtained slope value (-1.2) match well with findings based on a much longer observation period on stands of shrub species in a different area (Schlesinger and Gill, 1980). The self-thinning slope is more widely accepted, on theoretical grounds, as being -1/2 (or -3/2), as originally suggested by Yoda et al. (1963) and subsequent researchers, the regression slopes based on the data of this study are much closer to - 4/3 (based on mean biomass as shown in Fig. 2).
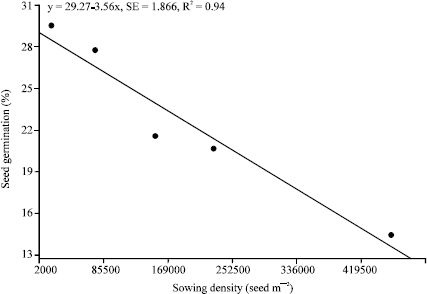 | |
| Fig. 1: | Seed germination as influenced by sowing density of Melastoma malabathricum |
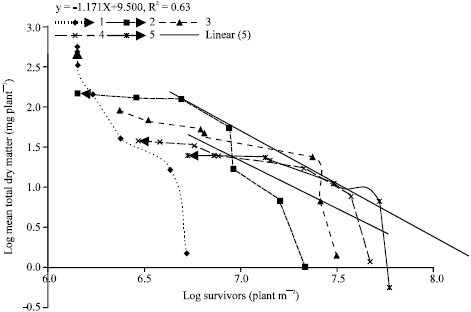 | |
| Fig. 2: | The relationship between the total dry matter weights per plant and plant densities of survivors for populations of Melastoma malabathricum sown at five densities (1-5) with the lines joint populations of the five sowing densities harvested on seven successive occasions indicating the trajectories, over time (40-160 DAP) and these population would have followed. Arrows indicate the directions of the trajectories, or the direction of self-thinning. At low densities especially, growth and hence mean dry weight is roughly independent of density but with ensuing plant growth over time, density-dependent reduction in growth compensation is denoted by variations in density, leading to achievement of fairly constant dry matter and slope of -1.17 |
| Table 1: | Gradient and intercept values for the thinning lines of populations of Melastoma malabathricum grown under various plant densities regimes and plant biomass (mg), calculated by principal components analysis |
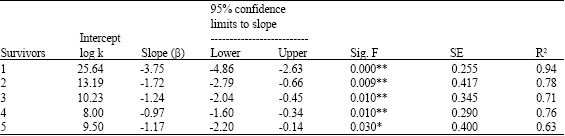 | |
| **, *Mean the relationship between two variables in the regression model is significant at p<0.01 or p<0.05. The equation of the lines is log w = log k + β log Ï? where, w is plant biomass (mg), Ï? is the density of survivors and k and β are constants | |
The slope of -1/3 (or -4/3) have also been reported more frequently in recent empirical studies (Bi, 2004; McCarthy and Weetman, 2007). The relation of the mean plant biomass and the survivors conforms to the power law with an ideal slope value is within the 95% confidence limits of the slopes of the significant models at p<0.05 (Table 1). The same results depicting the similar slope structure(s) have been already reported by Kays and Harper (1974), Westoby and Howell (1986) and Zhang et al. (2005).
The slope line is close to -1 but not exactly -1 and the relationship between dependent variable (w) and an independent or explanatory variable (p) is ultimately allometric (Marquet et al., 2005). In intra-specific competition, the resultant populations based on differential birth rates and death rates of plant (whether as genet or ramet entities), or number leaves or branches per plant, (as ramet entities), can regulate individual plant production or population at stable density from a very wide range of initial densities, bringing them to a narrow range of final densities and production per unit area and it therefore trends to keep density within certain limits (Fig. 2).
Figure 2 shows the fluctuations within plant biomass in single plant or per unit in populations of M. malabathricum whereas plant density decreases leads to parallel decrease in plant biomass. It is a schematic representation of the effects self-thinning on the numbers and individual plant weights with time in Melastoma malabathricum populations. It is envisaged that each population will start to thin along a line of slope from -3.7 to -1.2 until it reaches the maximum standing crop. Mortality during the phase of self-thinning is largely among individuals suppressed by the ensuing growth of neighbours, resulting in increased shading within the canopies of neighbouring plants. It also depicts the likely patterns by which populations of M. malabathricum might increase from an initially very small size of above-ground biomass (when the plant colonizes a previously unoccupied area) only to reach the asymptote as time progresses. If a succession of time intervals is taken singly, then each final density can be treated as the initial density for the next time-interval.
One-way ANOVA and Tukey`s Post Hoc Test (data not shown) indicate the mean plant was significantly (p<0.05) higher at low densities (Fig. 2). This phenomenon maybe caused by more photosynthetic activity among leaves because of leaf size as mean weight of leaves per plant was more at low density regimes. The total dry matter (m-2) was constant over a wide range of densities because individual plant displayed density-dependent reduction in growth rate and hence in individual plant size, in particular, because the reductions in mean plant weight compensated exactly for increase in density.
The relationship between mean shoot weight per plant (calculated by dividing total shoot weight per sample by the number of survivors) and the density of survivors in the populations conforms with the power law; thinning occurred along a line with a slope of -1.10 (R2 = 0.63, p<0.05) and 95% confidence limits to slope -2.07 (or -0.12) an intercept, log k, of 8.59 and then deflect from it as dead genets accumulate within the population (Fig. 3, Table 2).
Under the environmental regime of the present experiment, being conducted in the insect-proof house with mean midday radiation of 622 and 125 μmole photon m-2 sec-1 outdoor and inside in insect-proof house , respectively, therefore, with reduce light intensities, maximum yield is reduced; and the slope is not exactly -1 but it is close to -1 or - 4/3 (Table 1). In practice thought ,few self-thinning populations reach these maximum yields; and self-thinning populations with the slope of exactly -1 are rare (Begon et al., 1990; Guo and Rundel, 1998). Table 1 show all lines have a slope of approximating to -3/2 within the 95% confidence limits. The mean line slopes were less than -3/2 because of M. malabathricum is perennial shrub and the experiment conducted for a short period of 6 months.
The regression slopes, however, could change dramatically and if this happens, it reflects the integrated effects of all external influences like as disturbances, grazing, disease, drought, or other stress-inducing factors (Guo and Rundel, 1998). Different regression algorithms will produce different slope estimates, depending on the error structure of the data and the correlation between the bivariate data (Hamilton et al., 1995; Bi, 2004; Zhang et al., 2005).
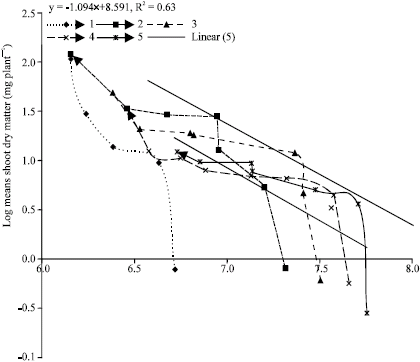 | |
| Fig. 3: | Self-thinning in Melastoma malabathricum populations sown at five densities (1-5) against mean shoot biomass (mg plant-1) with the lines joint populations of the five sowing densities harvested on seven successive occasions. They therefore indicate the trajectories , over time (40-160 DAP), that these populations would have followed. The arrow indicates the directions of the trajectories, i.e., the direction of self-thinning. The gradient of the thinning slope was -1.10 |
| Table 2: | Gradient and slope intercept values for the thinning lines within populations of Melastoma malabathricum grown under various plant densities regimes and mean shoot biomass (mg plant-1) , calculated by principal components analysis |
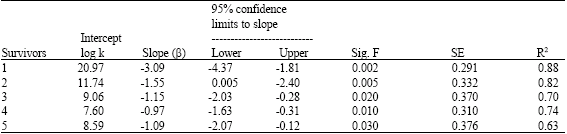 | |
| +Intercept (k), slope (β), R2 correlation coefficient and thinning populations by the reduced major axis using the equation Log w = log k- β log p, where, w is mean shoot dry weight per plant, Ï? is the density of survivors | |
The slope value for self-thinning has been argued that the -3/2 power rule should be the -4/3 law (Ogawa, 2001; Niklas, 2003; Niklas and Spatz, 2006) based on the metabolic theory (West et al., 1997). Other researchers (Adler, 1996; Chen et al., 2008) also provided a range of slope to describe the self-thinning rule.
| Table 3: | Allometric relationships between mean leaf mass (mg plant-1) and mean root for Melastoma malabathricum grown at five plant densities |
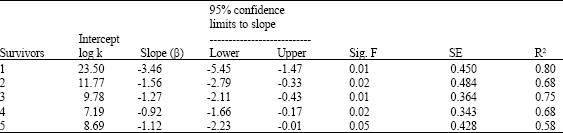 | |
| +Intercept (k), slope (β), R2 correlation coefficient and thinning populations by the reduced major axis using the equation Log w = log k-β log p | |
If succession of time intervals is taken singly, then each final density can be treated as the initial density for the next time interval. Thereafter, the biomass increased less and less with each time intervals until the population reached its carrying capacity (resources of the environment that can just maintain the population size in without a tendency to either increase or decrease (Begon et al., 1990).The biomass might therefore be expected to follow a sigmoidal curve due to ensuing competition vis- à -vis the onset of competition between survivors in high densities than low densities. This is a consequence of the hump in its recruitment rate curve, which is itself a consequence of intra-specific competition. When the log of average leaf weight per plant was plotted against the log of density of survivors for a crowded even-aged plant population, in such a way that the population`s trajectory was held under a line of slope -1.12 (R2 = 0.58, p<0.05) and 95% confidence limits to slope -2.23 (or -0.01) with an intercept, log k, of 8.69 (Table 3).
The results of this experiment suggests that there are limited resources available for M. malabathricum plant growth and that at high densities these are shared among the bigger number of competing individuals. If this would be the case, it is expected that provision of extra resources would allow greater growth of individuals` plants and greater yield per unit area. We have seen that intra-specific competition of M. malabathricum can, over a period of time, could influence the number of deaths, the number of births, among genets or ramets, or both, the amount of growth and the distribution of biomass within the population. With progressing the time, the individuals grow in size, their requirements increase and they therefore compete at an increasingly greater intensity. This, in turn, tends to gradually increase their risk of dying. Thus, the number that survived and the growth rate of the survivors are simultaneously influenced by density.
A slope of -3/2 indicates that in a growing, self-thinning population, mean plant weight increases faster than the decreases in density. A population following a -3/2 thinning line will therefore steadily increase its total weight (or yield). Eventually, of course, this increase will cease as yield cannot increase indefinitely. Instead, the thinning line might be expected to change from slope of -3/2 to a slope of -1, in such a way that the increase in mean plant weight is likely compensated by as the decrease in density.
A slope of -1 indicates that the further growth of survivors is exactly balanced by the deaths of other individuals. Upon reaching the asymptote with the maximum total yield possible, no further increase is possible for the said species in question in that environment (Begon et al., 1990) .
The mean weight of leaves per plant was influenced by the plant density treatments and the highest leaves weight per plant was recorded in the lowest plant density. As the surviving leaves in the canopy grow old, their photosynthetic activity falls below that required balancing respiratory load so that leaves, branch and eventually whole genets begin to die. Consequently, the number of live plants and the proportion of live matter within the population decreases (Donohue and Schmitt, 1999). Further statistical analysis using logarithmic regression on the relationship between plant height (cm) and log density of survivors as a function of time confirmed that the power law prevailed whereby thinning occurred along a line with a slope of -1.42 (R2 = 0.71, p<0.05) and within the 95% confidence limits to slope -2.46 (or -0.38), with an intercept, log k, of 10.36 (Table 4).
| Table 4: | Gradient and slope intercept values for the thinning lines of populations of Melastoma malabathricum grown under various plant densities regimes and plant height (cm), calculated by principal components analysis |
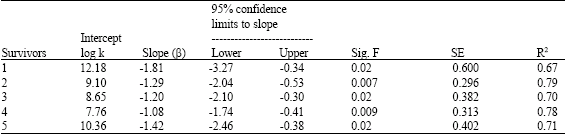 | |
| +Intercept (k), slope (β), R2 correlation coefficient and thinning populations by the reduced major axis using the equation Log w = log k- β log p, where, w is plant height, Ï? is the density of survivors | |
| Table 5: | Gradient and intercept for the thinning lines of populations of Melastoma malabathricum grown under various plant densities regimes and mean root biomass (mg plant-1), calculated by principal components analysis |
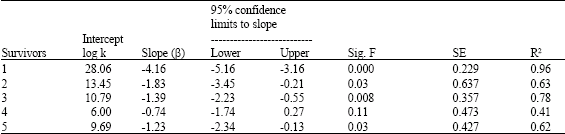 | |
| +Intercept (k), slope (β), R2 correlation coefficient and thinning populations by the reduced major axis using the equation Log w = log k- β� log p | |
Plant responses to crowding may be mediated by resource availability and/or by a specific environmental cue, with the ratio of red:far red wavelengths (R:FR) perceived by phytochrome (Donohue and Schmitt, 1999).
Root biomass was significantly (p<0.05) affected by density, being greater at low density than at high density (data not shown). Root biomass decreased with increasing plant density in all harvests, indicating that root growth was negatively related with plant density. The logarithmic models between different plant survivors at different densities and dry weight of root conforms to the power law; whereby thinning occurred along a line with a slope of -1.23 (R2 = 0.62, p<0.05) and within the 95% confidence limits with slope values registering -2.34 (or -0.13) and an intercept, log k, of 9.69. It showed that increased root competition could lower the slope and/or intercept of the self-thinning line transverse by plant populations (Table 5).
As root: shoot ratios were remarkably constant over the broad range of plant size achieved in the five densities. Thus, these plants do not exhibit above- versus below-ground biomass trade-offs in their ability to compete for light versus below-ground resources.
Plasticity in biomass allocation, root morphology and root distribution pattern has been found to be an important adaptive mechanism to acquire nutrient resources (Xie et al., 2001, 2007). As with shoot biomass, root biomass increased in low density levels. The rate of increase slowed or became negative with increasing the population density. The value of the self-thinning exponent was -1.23 at the final phase, 160 days after transplanting of stand development.
In summary, present results support the concept of competition-mediated self-thinning rule. The different self-thinning power is a plant response to the resource utilization and sensitivity to stress. Because regression slopes are affected by multiple factors, they can vary greatly among species and habitats. Regression analysis could be very helpful in identifying the presence of other factors that affect the development of ecological communities. More well-controlled experiments should be carried out in order to identify which is more accurate values between -3/2 and -4/3 or -1.
By considering all morphological attributes of the different growth forms suggests that growth form differentiation may be a plastic response to increasing levels of density stress. For example, increased density in plant populations commonly resulted in decrease in plant size as a result of increasing competition for limited resources (Harper, 1977). Likewise, the decreased allocation of resources to sexual reproduction is a common response to high levels of intra-specific competition (Harper, 1977).
ACKNOWLEDGMENT
We acknowledge the financial assistance through Fundamental Grant No. PS074/2007B given to the first author by the University of Malaya and to the Institute of Biological Sciences, University of Malaya for providing the facilities throughout this study.
REFERENCES
- Adler, F.R., 1996. A model of self-thinning through local competition. Proc. Nat. Acad. Sci. USA., 93: 9980-9984.
Direct Link - Bi, H., 2004. Stochastic frontier analysis of a classic self-thinning experiment. Aust. Ecol., 29: 408-417.
CrossRefDirect Link - Donohue, K. and J. Schmitt, 1999. The genetic architecture of plasticity to density in impatiens capensis. Evolution, 53: 1377-1386.
Direct Link - Guo, Q. and P.W. Rundel, 1998. Self-thinning in early postfire chaparral succession: Mechanisms, implications and a combined approach. J. Ecol., 79: 579-586.
CrossRef - Hamilton, N.R.S., C. Matthew and G. Lemaire, 1995. In defence of the-3/2 boundary rule: A re-evaluation of self-thinning concepts and status. Ann. Bot., 76: 569-577.
CrossRef - Hara, T., F. Koike and K. Matsui, 2006. Crowding effect in marine macrophytic algae populations. J. Plant Res., 99: 319-321.
CrossRefDirect Link - Inouye, R.S., 1980. Density-dependent germination response by seeds of desert annuals. Oecologia, 46: 235-238.
CrossRef - Kays, S. and J.L. Harper, 1974. The regulation of plant and tiller density in a grass sward. J. Ecol., 62: 97-105.
Direct Link - Li, B.L., H. Wu and G. Zou, 2000. Self-thinning rule: A causal interpretation from ecological field theory. Ecol. Model., 132: 167-167.
Direct Link - Li, X.L., 2002. Competition-density effect in plant populations. J. Forest. Res., 13: 48-50.
CrossRefDirect Link - Lonsdale, W.M. and A.R. Watkinson, 1982. Light and self-thinning. New Phytol., 90: 431-445.
Direct Link - Marquet, P.A., R.A. Quiñones, S. Abades, F. Labra and M. Tognelli et al., 2005. Review scaling and power-laws in ecological systems. J. Exp. Biol., 208: 1749-1769.
CrossRefDirect Link - McCarthy, J.W. and G. Weetman, 2007. Self-thinning dynamics in a balsam fir (Abies balsamea (L.) Mill.) Insect-mediated boreal forest chronosequence. For. Ecol. Manage., 241: 295-309.
Direct Link - Mohler, C.L., P.L. Marks and D.G. Sprugel, 1978. Stand structure and allometry of trees during self-thinning of pure stands. J. Ecol., 66: 599-614.
Direct Link - Niklas, K.J., 2003. Reexamination of a canonical model for plant organ biomass partitioning. Am. J. Bot., 90: 250-254.
Direct Link - Niklas, K.J. and H.C. Spatz, 2006. Allometric theory and the mechanical stability of large trees: Proof and conjecture. Am. J. Bot., 93: 824-828.
Direct Link - Ogawa, K., 2001. Time trajectories of mass and density in a chamaecyparis obtusa seedling population. For. Ecol. Manage., 142: 291-296.
CrossRef - Polley, H.W., B.J. Wilsey and J.D. Derner, 2008. Do species evenness and plant density influence the magnitude of selection and complementarity effects in annual plant species mixtures? Ecol. Lett., 6: 248-256.
CrossRef - Quinones, R.A., T. Platt and J. Rodriguez, 2003. Patterns of biomasssize spectra from oligotrophic waters of the Northwest Atlantic. Prog. Oceanogr., 57: 405-427.
CrossRef - Rivera, M. and R. Scrosati, 2008. Self-thinning and sizeinequality dynamics in a clonal seaweed (Sargassum lapazanum, Phaeophyeae). J. Phycol., 44: 45-49.
CrossRefDirect Link - Schlesinger, W.H. and D.S. Gill, 1980. Biomass, produc-tion and changes in the availability of light, water and nutrients during the development of the chaparral shrub, Ceanothus megacarpus after fire. J. Ecol., 61: 781-789.
CrossRef - Simard, S.W. and B.J. Zimonick, 2005. Neighborhood size effects on mortality, growth and crown morphology of paper birch. For. Ecol. Manage., 214: 251-265.
Direct Link - Weiner, J., P. Stoll, H. Muller-Landau and A. Jasentuliyana, 2001. The effects of density, spatial pattern and competitive symmetry on size variation in simulated plant populations. Am. Nat., 158: 438-450.
Direct Link - West, G.B., J.H. Brown and B.J. Enquist, 1997. A General model for the origin of allometric scaling laws in biology. Science, 276: 122-126.
CrossRef - Westoby, M. and J. Howell, 1986. Influence of population structure on self-thinning of plant populations. J. Ecol., 2: 343-359.
Direct Link - White, J. and J.L. Harper, 1970. Correlated changes in plant size and number in plant populations. J. Ecol., 58: 467-485.
Direct Link - Wiegand, K., D. Saltz, D. Ward and S.A. Levin, 2008. The role of size inequality in self-thinning: A pattern-oriented simulation model for arid savannas. Ecol. Model., 210: 431-445.
Direct Link - Xie, Y., S. An, B. Wu and W. Wang, 2006. Density-dependent root morphology and root distribution in the submerged plant vallisneria natans. Environ. Exp. Bot., 57: 195-200.
Direct Link - Xie, Z.M., Z.H. Ye and M.H. Wong, 2001. Distribution characteristics of fluoride and aluminum in soil profiles of an abandoned tea plantation and their uptake by six woody species. Environ. Exp. Bot., 26: 341-346.
Direct Link - Yoda, K., T. Kira, H. Ogawa and K. Hozumi, 1963. Self-thinning in overcrowded pure stands under cultivated and natural conditions. J. Inst. Polytech. Osaka City Univ., 14: 106-129.
Direct Link - Zeide, B., 2001. Natural thining and environmental change: An ecological process model. For. Ecol. Manage., 154: 165-177.
Direct Link - Zhang, H., G.X. Wang, Z.O. Liu, Z.X. Sheen and X.Z. Zhao, 2005. Sensitivity of response to abscisic acid affects the power of self-thinning in Arabidopsis thaliana. Bot. Bull. Acad. Sinica, 46: 347-353.
Direct Link