
Solomon Tefera
Department of Animal Production and Health, Faculty of Agriculture, University of Swaziland, PO M 205 Luyengo Campus, Swaziland, Southern Africa
B.J. Dlamini
Department of Animal Production and Health, Faculty of Agriculture, University of Swaziland, PO M 205 Luyengo Campus, Swaziland, Southern Africa
A.M. Dlamini
Department of Animal Production and Health, Faculty of Agriculture, University of Swaziland, PO M 205 Luyengo Campus, Swaziland, Southern Africa
Research Journal of Botany
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 49-64







ABSTRACT
A study was conducted to assess woody vegetation pattern in the semi-arid savannas of Swaziland under three land use systems (communal land, government ranch, game reserve) and three soil classes (vertisol, lithosol and raw mineral). Communal perceptions towards the indigenous use of woody plants were also assessed. A total of 90 woody plant species were identified. Fifty eight percent of the identified woody plants have browse value to livestock. The mean density of total woody plants combined ranged between 1352 TE ha-1 and 11 533 TE ha-1. In severely encroached area, the total woody plant density was greater (p< 0.001) on communal land (8578 TE ha-1) than in the ranch (5608 TE ha-1) and game reserve (4123 TE ha-1) and on raw mineral soil than lithosol. In less encroached areas, land use had no significant differences, but lithosol (2024 TE ha-1) had greater density (p< 0.05) than vertisol (1237 TE ha-1). The most important encroaching species in the study areas were: A. tortilis, A. nilotica, C. odorata, A. burkei, C. apiculatum, S. caffra and D. cinerea. All the sites in the current study were dominated by height class of >0.05 -2 m, which suggested the potential for long-term increase. There is an indication that bush encroachment showed non to significant variations at small scale (between land uses, soil classes) and large scale levels of resolution (between study areas). At small scale level, significant variations may be due to past and present grazing pressure as well as inherent diversity and nature of the soil. This study suggests in severely encroached area in particular that generalized statements about the relatively bad condition of any land use system and the permanence of any specific condition must be treated with caution.
PDF Abstract XML References
How to cite this article
Solomon Tefera, B.J. Dlamini and A.M. Dlamini, 2008. Dynamics of Savannas in Swaziland: Encroachment of Woody Plants in Relation to Land Use and Soil Classes and Indigenous Knowledge on Plants Utilization. Research Journal of Botany, 3: 49-64.
URL: https://scialert.net/abstract/?doi=rjb.2008.49.64
URL: https://scialert.net/abstract/?doi=rjb.2008.49.64
INTRODUCTION
The savannas of Africa support large, diverse and dynamic ecosystems abundant in game, livestock and other agriculture enterprises that constitute the biggest primary production sector for many countries in the continent. Nevertheless, the productivity and long-term economic viability as well as the ecological integrity of savannas is undermined by increasingly dense thickets of woody vegetation often termed as bush encroachment.
Bush encroachment has been documented in many semi-arid rangelands in Southern Africa (Roques et al., 2001; Francina and Smit, 2006; Wigley, 2006). The theory of bush encroachment in savannas was presented by Walker and Noy-Meir (1982) and Noy-Meir (1982). This model described savannas as a system of two functionally separate soil layers and postulated the utilization of soil moisture by woody vegetation and herbaceous layer from the two distinct soil profiles. It also stated that a healthy grass layer may out-compete woody species for water in the surface soil. Therefore, a decrease in the grass layer increases moisture available for the woody plants. Cattle are responsible for opening up the grass sward and so enabling woody plant species to have access to soil moisture that could otherwise have been utilized by the grasses (Walker and Noy-Meir, 1982; Bland, 1985). Limited evidence is available on whether or not game animals are also accountable for increase in the density of woody vegetation. Bush encroachment can alter soil moisture, nutrients and micro climatic condition of rangelands and this is often accompanied by decreases in herbaceous productivity (Richter et al., 2001; Roques et al., 2001). Many studies (Epstein et al., 2002; Hudak et al., 2003; Satti et al., 2003) reported that woody plants also affect the distribution of soil nutrients. Overall declining conditions are symptomatic of rangelands where the ecosystems are under ecological stress in the form of bush encroachment. Encroachment of woody plants is continuously threatening the pastoral production and challenging the sustainability of livestock and game farming (Francina and Smit, 2006; Solomon et al., 2007a). In East Africa, an increase in bush cover by 10% reduces grazing by 7% and grazing is eliminated completely by 90% bush cover (Van Wijingaarden, 1985). In the Kalahari of Namibia, increase in bush cover reduces the pastoral industry by 75% (Adams, 1996).
Given that the savannas of Southern Africa support large and expanding human population, a loss of rangeland productivity through bush encroachment is of great significant. Therefore, the question of what determines woody encroachment is of general interest. In addressing the problem in this region, recent studies have used series of aerial photographs on rangelands with known fire history, land use practices and precipitation records (Roques et al., 2001; Wigley, 2006). The use of aerial photograph can be exclusively advantageous in that it covers unreachable vast areas, but this method lacks objectivity in terms of determining density, identifying noxious invaders and of determining size class distribution. Understanding of the advancement of bush encroachment, its consequences and the planning of the control measures requires quantitative data on species composition, density and height distribution classes of woody plants (Abule et al., 2005). Attempts were not made in the past to investigate the density and structure of woody plants in the semi-arid rangeland of Swaziland in general and taking into account the different land use practices and soil classes in particular. In addition, despite the ecological problems of bush encroachment, several woody plants have been used by the rural people for various purposes. However, the Indigenous Knowledge (IK) associated with the rich biodiversity of woody plants have not been adequately documented.
The current study was therefore undertaken to understand the effect of land use systems (communal, ranch and game reserve) on the distribution and composition of woody plants, understand the effect of soil class (vertisol, lithosol and raw mineral) on the distribution and composition of woody plants and to assess the forage and traditional values of woody plants based on the perceptions of the communal people.
MATERIALS AND METHODS
Study Area
This study was conducted in the Lowveld savannas of Swaziland. Altitude in the Lowveld ranges from 200 m to 300 m above sea level. The area has more of a semi-arid climate with mean annual rainfall ranging from 400-600 mm, while the mean annual temperature varies between 18 and 26°C (Monadjem and David, 2005). The vegetation has been classified as lowveld savanna (Acocks, 1988). Three major vegetation types were described, namely broad leaved woodland savannas, rangelands dominated by Acacia and the allied genera and riverine forest along the rivers and major drainage lines (Sweet and Khumalo, 1994).
Site Selection and Layout
Three land use systems: communal grazing land, government ranch and game reserve located adjacent to each other were identified in two areas locally named as Bigbend and Simunye (Fig. 1). The two areas differ mainly in the distribution of dominant soil classes and landscape features and therefore, were not considered as spatial replicates. Bigbend area is dominated by vertsiol and lithosol classes whereas raw mineral soil and lithosol represent the dominate soil classes in Simunye area. Six sites were selected, one on each of the two common soil classes for each of the three land use systems in the two areas. Three hundred meter transects were laid out randomly on each site, giving a total of 36 transects in the two areas with each area having equal number of transects. Unfortunately, there is a lack of accurate information on the grazing history of the land uses in these areas. Besides, a sequence of historical aerial photographs to document the extent of change over time was not available. However, Sweet and Khumalo (1994) and Dlamini et al. (2000) estimated that the stocking rate in the communal grazing land was between 2 ha LSU-1 and 0.94 ha LSU-1 before the year 2000. The government ranches represent sites with a moderate to heavy grazing pressure some years ago and the average stocking rate in recent years was estimated to be 6 ha LSU-1. Similarly, the game reserves represent varied history of heavy grazing pressure with recent average stocking to be 4.4 ha LSU-1.
Survey of the Woody Vegetation Layer
Three 10x10 m2 plots (Solomon, 2003) were evenly laid out along each transect to record the variables of the woody vegetation layer. All rooted live woody plants were recorded and counted in each plot for the analysis of density, species composition and diversity. For multi-stemmed plants, an arbitrary decision was made to count the plant as a separate individual if the stems were at least 400 mm apart. Plants were recorded by species and divided into height classes of >0-0.5; >0.5-1; >1-1.5; >1.5-2; >2-3; >3-4; >4-5 and >5 m (Friedel, 1987). The plant data was then standardized to tree equivalent (1 TE = 1 shrub/ tree, 1.5 m high) for determining the total density per unit area. Indigenous knowledge and perceptions on the use of woody plants was collected from two groups of 20 communal people per group with open ended interviews. Data on vegetation and perceptions of the communal people were collected late in the growing season (March-April, 2006).
Statistical Analysis
To limit other influencing factors, the three land uses in the two study areas (Bigbend and Simunye) were chosen within a small area and this occurred only in one place in each study area. The closeness of the sites allows transects to constitute dependent representative samples of a specific soil type for the separate land uses. Therefore, field experiment in present study contained six treatment combinations with three replicates for each study area. Contrary to the design commonly applied in planned field experiment, the three transects for each treatment combination are, by necessity spatially clustered due to variations in soil and other factors over short distances.
In present study, the risk of pseudoreplication was expected because the sample area was smaller or more restricted to infer about the land use and soil effects. However, it was assumed that the errors due to pseudoreplication were lower than those associated with the non-representativeness of taking large area and those with time variables for collection of vegetation data over large sampling area. Pseudoreplication should not be considered as inappropriate as to sampling design and statistical analysis, or to the use of allowable generalization (Annika, 2000). It is often difficult to conduct ecological research at landscape level as controlled experiments and/or is costly and the problem as discussed above can seldom be avoided. Most ecological research at landscape or regional level is inferential and inductive, but conclusion drawn out need not be of less interest (Hargrove and Pickering, 1992).
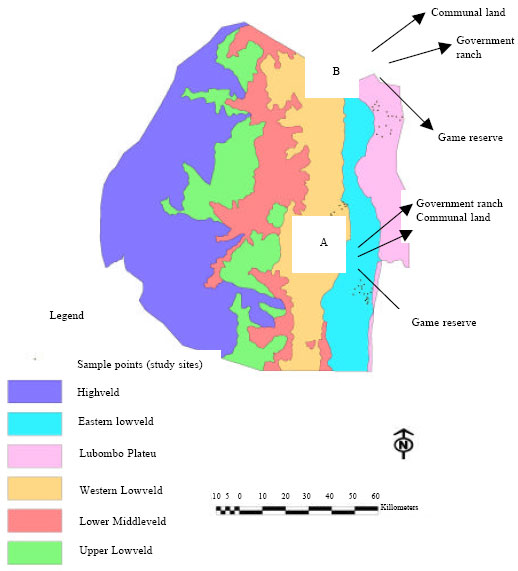 | |
| Fig. 1: | Map of the study sites in the lowveld of Swaziland: Bigbend (A) and Simunyae (B). Points show the soil classes and transects under each land use system |
Since data on height distribution, density and diversity of woody plants were not normally distributed and transects were few, the non-parametric Whitney U-test for two independent samples was used (Kent and Coker, 1992). The significance levels found were not compared to priori requirements, but instead used as semi qualitative ‘relevance indicators’ for individual differences. Shannon diversity index was employed to compute woody plant diversity for each site (Kent and Coker, 1992). For data that did not require analysis, simple descriptive statistics were employed where appropriate.
RESULTS
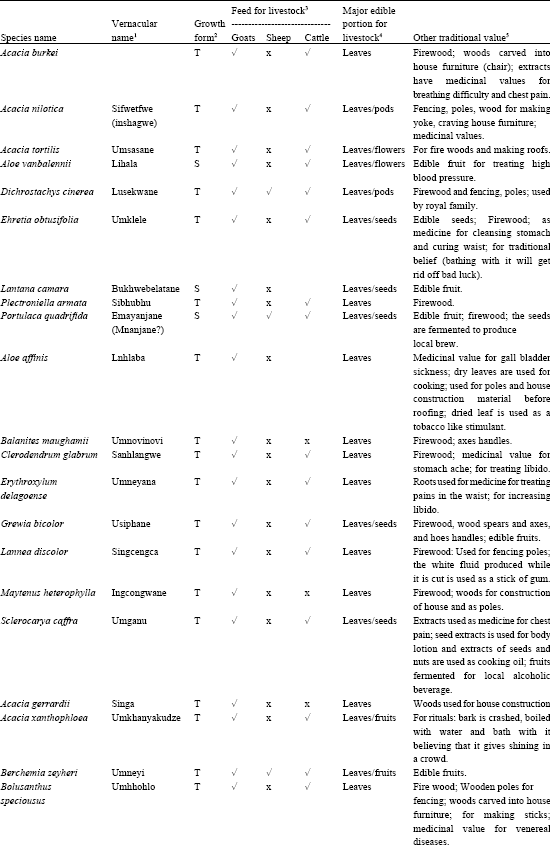
A total of 90 woody plant species were recorded for the two study areas of which seven were Acacia species as shown in Table 1.
| Table 1: | Scientific and vernacular name, growth form, forage and other traditional household uses of native woody plants identified in the lowveld of Swaziland |
 | |
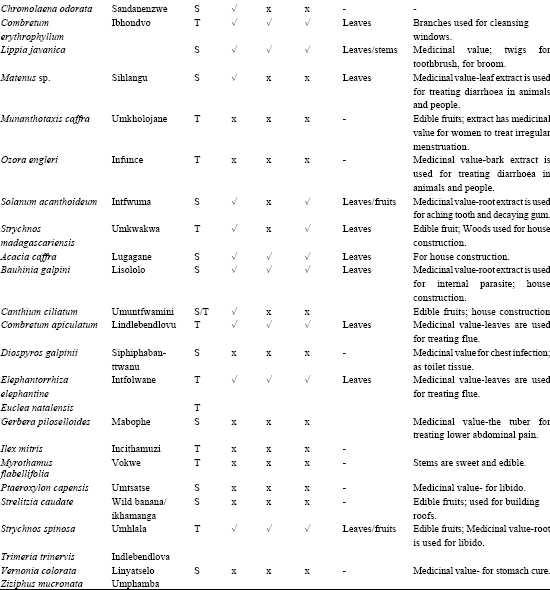 | |
| 1= Swazi names; 2= S-shrub, T-tree; 3,4,5 = Interview of herdsmen and communal people (n = 40) | |
Encroachment of woody plants in Bigbend area
Density of all woody plants combined, seedlings and small shrubs (>0-1 m) did not show differences among the land use systems (Table 2, 3). Shrubs in height class >1-1.5 m were more (p<0.05) common in the government ranch than game reserve, while the abundance on communal land was not apparently different from the ranch or the game reserve. Shrub density with range of >1.5-2 m height was similar to all land use systems. In contrast, large difference (p<0.05) was evident in height class >2-3 m, with the communal land having greatest density. Density of trees in height class >3-4 m was not affected by land use. Trees in height class >4-5 m were not recorded on communal land and in the ranch. Mature trees (>5 m) were less frequent on communal land than in the other land uses. As for soil classes, density of all woody plants combined, young shrubs (>1-1.5 m) and mature trees (>5 m) were significantly (p<0.05) lower on vertisol than lithosol, but other classes were remarkably similar (Table 2).
| Table 2: | Total density of woody plants (mean TE ha-1) under three land use systems and two soil classes in Bigbend and Simunye areas |
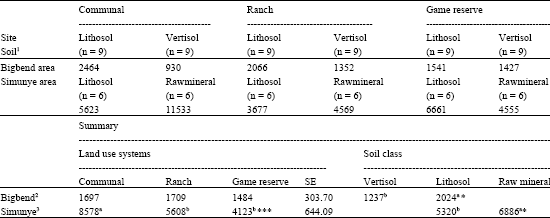 | |
| 1: n = 9 per soil class; 2: SE-land use = 303, soil class = 428 3: SE-land use = 644, soil class = 525.9. *: Significant (p<0.05); ***: Highly significant (p<0.001) Means in the same raw with different superscripts are significantly different. | |
| Table 3: | Total woody plant density (mean number of plants ha-1) by height class under the land use systems and soil classes in Big bend area |
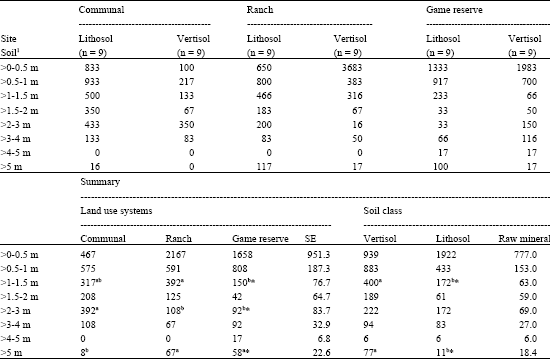 | |
| *: Significant (p<0.05), Means in the same raw with different superscripts are significantly different | |
In addition, soil classes within each land use system shows a unique pattern of density and size class distribution (Table 3).
Most Common Woody Plants and Diversity
The most important encroaching species identified in Bigbend area were in descending order: D. cinerea, A. burkei, A. tortilis, A. nilotica, E. obtisifolia and P. quadrifida. Dichrostachys cinerea was more common on communal land than in the ranch and game reserve. Acacia burkei and P. quadrifida were more abundant in the ranch (Table 4). For E. obtisifolia, a significantly lowest density was obtained on communal land.
| Table 4: | Density of common woody plant species (mean TE ha-1) in relation to the land use systems and soil classes in the two study areas |
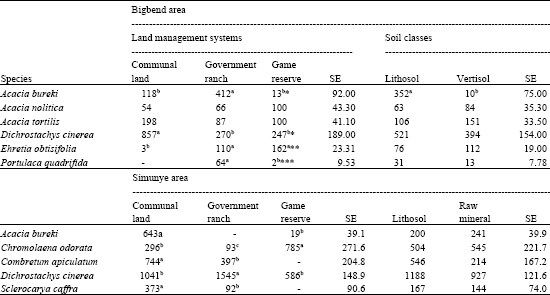 | |
| *= Significant (P < 0.05); **/*** = Highly significant (p<0.01/0.001) Means in the same raw with different superscripts are significantly different. | |
| Table 5: | Total woody plant density (mean number of plants ha-1) by height class under the land use systems and soil classes in Simunye area |
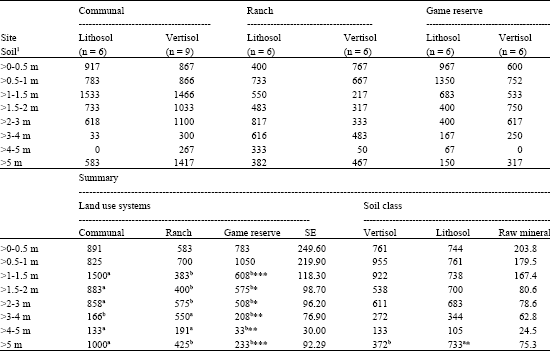 | |
| *: Significant at p<0.05; **/*** = Highly significant (p<0.01/0.001). Means in the same raw with different superscripts are significantly different | |
The density of A. tortilis and A. nilotica was not significantly affected by land use. As regards to soil classes, the most striking difference was obtained only for A. burkei.
Shannon’s diversity was significantly greater in the ranch than the other land uses. There were more diverse plants on lithosol than vertisol (Fig. 2). It was observed that 24% of the total woody plants were common to the three land use systems, 6% to the communal and the ranch, 18% to the communal and game, 15% to the ranch and game reserve.
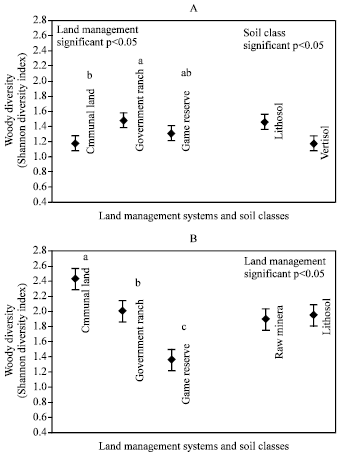 | |
| Fig. 2: | Woody plant diversity index in the three land management systems and two soil classes in Bigbend (A) and Simunyae (B) area |
The percentage of woody plants common to any of the land use system varies between 9 to 18%. As for the soil classes, 50% of the woody plants were common to the two soil classes and 26% to only the lithosol soil.
Encroachment of Woody Plants in Simunye Area
Total density for all woody plants combined differed significantly among the land use systems. This was greater on communal land, followed by government ranch (Table 2). The density of seedlings (>0-0.5 m) and young shrubs (>0.5-1 m) did not show marked difference among the land use systems. There were more shrubs (>1-2 m) and trees (>2-3 m) on communal land than in the other land use systems. For trees >3-4 m height, the communal land and game reserve had similar and significantly lower densities than the ranch. There were more trees (>4 m) on communal land than in the other land use systems. As regards to soil classes the density of all woody plant combined was greater (p<0.05) on raw mineral than lithosol. However, soil classes within each land use showed a different pattern of variation. Inconsistent results were found for size class distribution between soil classes within each land use, but in general, except for mature trees above 5 m, significant differences were not evident between lithosol and raw mineral soil (Table 5).
Most Common Woody Plants and Diversity
Among the common woody plants identified in Simunye area, the most important encroaching species in descending order were D. cinerea, C. odorata, C. apiculatum, A. burkei and S. caffra. Dichrostachys cinerea was more abundant in the ranch than on communal land and game reserve. Encroachment of C. odorata was more severe in the game reserve, followed by communal land (Table 4). Combretum apiculatum and S. caffra were not recorded in the game reserve, but the communal land had nearly two and three times the TE density in the ranch, respectively. Acacia burkei was more common on communal land than in the game reserve.
Shannon’s diversity index values revealed that communal land had the richest plant diversity where as the game reserve had the lowest. As for the soil classes, diversity on lithosol and raw mineral soil was similar (Fig. 2). Seven percent of the total wood plants identified were common to the three land use systems, 12% to communal and the ranch, 13% to communal and game, 4% to the ranch and game reserve. Thirty-three percent of the total woody plants was found only on communal land, 29% in the ranch and 8% in the game reserve. Results for the soil classes indicated that 30% of the total woody plants were common to the two soil classes while the lithosol had 36.
Common Use of Woody Plants
Opinions of the communal people indicated that 58% of the total woody plants identified has forage values to livestock. Twenty-one percent was used by the three livestock species, 39% by goats and cattle and 35% browsed by goats. Leaves are the major edible portion of most browse plants followed by the seeds. The role of woody plants in other traditional uses was also discussed (Table 1). The result showed that 87% of the total plants had traditional values in addition to the role as browse plants. These include fencing and firewood (24 species), house construction (11 species), making house utensils and furniture (12 species), traditional medicine for people and livestock (41 species), human food (17 species), rituals (5 species) and local beverages and body lotions (3 species).
DISCUSSION
Total Density and Diversity
The overall mean total woody plant density in the lowveld savanna of Swaziland ranged from 1352 TE ha-1 to 11533 TE ha-1 which indicates the existence of great variability at site and landscape level of resolution. Comparison of the two study areas explained that bush encroachment is a more serious problem in Simunye than Bigbend area. Woody plant density of 2400 ha-1 was regarded as borderline between non-encroached and encroached condition in African Savannas (Roques et al., 2001). A density of 2500 TE ha-1 was considered as highly encroached condition in Southern Africa (Richter et al., 2001). Based on these results the lowved of Swaziland is in a state of light to severely encroached condition. Advancement of bush encroachment has been reported in many semi-arid regions of Southern Africa. Hudak and Wessman (2001) reported a 30% increase in woody plant cover between 1955 and 1996 for Madikwe Game reserve, South Africa. Roques et al. (2001) investigated that woody plant encroachment increased from approximately 3% to over 40% in 50 year in South African communal rangelands.
In the less encroached area (Bigbend), difference in soil class was more important than land use to cause variation in total woody plant density. In severely encroached rangelands (Simunye area) however, total density differed significantly among the land use systems and between soil classes. The current findings agreed with Smet and Ward (2005), who reported that shrub encroachment on communal land was more severe than in the other land use systems in the North Cape of South Africa which is also supported by a study conducted in the North-district of Botswana (Annika, 2000). In contrast, substantial increase in bushes for the three land use systems were reported in Hluhluwe area in Kwazulu-Natal of South Africa, with the communal land-the least severely affected (Wigley, 2006). The study of Solomon (2003) in East Africa rangelands had indicated that the magnitude of bush encroachment was not significantly related to land use practices.
In the current study, the communal land in a severely encroached area is characterized by an increasingly consistent heavy grazing pressure. Sweet and Khumalo (1994) and Dlamini et al. (2000) estimated that the stocking rate in the lowveld was between 2 and 0.94 ha LSU-1 before the year 2000. Although an estimate of the recent stocking rate is unavailable, improvement is not expected because of increased external forces that could reduce available grazing area (Solomon et al., 2007a,b, unpublished). The government ranch represents an area with a history of heavy grazing pressure some years ago and hence severe bush encroachment occurred. Recently, the stocking rate at the ranch was significantly reduced (perhaps due to the invasion of the ranch by C. odorata). Similarly, the game reserve was observed as an area with a history of heavy grazing pressure. Prior to its establishment (before 1986), the game reserve was utilized as cattle ranches with average estimated stocking rate of 4.4 ha LSU-1, but has only been sparsely populated in recent time. Since differences in land use are often linked to differences in stocking rate/land management practices, it is difficult to separate the effect of one from the other (Parsons et al., 1997). The high cattle densities commonly practiced in the communal area of Southern Africa were reported to be the main cause of bush encroachment (Skarpe, 1986; Ringrose et al., 1996). High grazing pressure may be a key determinant of shrub encroachment through grass fuel removal and concomitant reduction in fire frequency. On the other hand, lower fire frequency promotes woody plant recruitment and in the long run, may lead to shrub encroachment (Van Auken, 2000). Results obtained in the southern rangelands of Ethiopia suggested that external factors such as cultivation and land alienation inappropriate to the communal grazing systems compressed the available grazing land and changed spatial pattern of land use (Oba et al., 2000; Solomon et al., 2007a). Consequently, this will increase the grazing pressure on the remaining grazing land which will contribute to the advancement of bush encroachment. Some works in other parts of Southern Africa do not support the theory that high cattle densities are the main driving factors (Abel, 1992).
It is important to note that problems of scale could emerge when trying to correlate bush encroachment with land use systems. Results from grass-tree plot scale experiments within a small patch of savannas may contradict results of broad scale studies. Gosz and Sharpe (1989) and Anthony et al. (2006) noted that large scale vegetation pattern could be correlate with climatic (mainly rainfall) and other geographical variables. In addition, variations at small scale can occur due to site-specific characteristics other than land use such as soil discontinuities and proportions. Belsky (1990) noted that patchness of rainfall at small scale could occur and this may lead to patchy woody vegetation patterns but within a range of intermediate long-term rainfall levels only.
When comparing the soil classes in present study, heavier encroachment was noted on lithosol and raw mineral soil than vertisol. The former soils are characterized by a shallow layer of sandier soil (5-35 cm) compared to the later (>60 cm). Therefore, rooting niche separation may not be a prerequisite for woody plant encroachment because encroachment may occur on soils too shallow to allow for root separation (Wiegand et al., 2006). In contrast, Walker and Noy-Meir (1982) and Noy-Meir (1982) explained that savanna is a two layered soil water system in which the roots of woody plants and grasses are confined to separate soil layers. Booth and Barker (1981) contended that susceptibility of soils to bush encroachment increases as the soil becomes sandier while the texture which determines the hydrological functioning in turn affects the redistributed water use by the plants.
Indeed, the overall increase in woody plants in the study areas could be due to the introduction of plants into new areas and/or the gradual thickening of the existing vegetation. Ruminants may act as a dispersing agent by browsing certain woody plants (Radford et al., 2001; Tews et al., 2004) and their fruits and later disperse the encroacher seeds via the dung (Solomon, 2003). Some invasive shrubs that produce copious number of viable seeds can be dispersed by other agents such as wind, birds and water.
Woody plant diversity in present study showed a clear but non-consistent difference in relation to land use systems, whereas the difference in relation to soil classes was not apparent. Furthermore, this study portrayed that the communal land had the most diverse and rich species in severely encroached area which is in contrast with the published data of Smet and Ward (2005) who reported that the commercial ranch had more diverse and richer species composition than the other land use systems.
Densities and Proportional Distribution of Height Classes
All the sites in the study areas were dominated by shrubs (>0.5 -2 m) with an average of 44%. However, overall patterns of height class distributions differ between land use systems and soil classes. Mature trees (> 5 m) were recorded from very few in low density area to very high in severely encroached area. The dominance of small shrubs in encroached rangelands was reported in many savanna rangelands in Africa (Van Vegeten, 1981, Friedel, 1985; Solomon et al., 2007a). Three theories can explain the dominance of small, low growing plants in the current study (1) could be a result of genotypes with stunted growth and short life span (Van Vegeten, 1981) as reflected in the total woody plants identified in the present study to be 26% being shrub growth forms; (2) the age of the plant and (3) the consequence of spatial and temporal pattern of competition for water and nutrients between closely spaced woody plants (Solomon, 2003). The higher densities of woody plants in the height class >0.5-2 m could be an indication of the potential for long-term increase. However, smaller size classes are effectively controlled by burning provided there is enough grass fuel load to burn the bushes (Solomon, 2003). Regardless of growth forms, height class represents the age of the plant and its pattern of distribution explains different stages of invasion. Therefore, control of bush encroachment may require a thorough understanding of the different growth stages and the life cycle of the plant.
Common Woody Plants
Many encroaching woody plants identified in present study showed variability in total density at landscape level of resolution. E. obtisifolia, P. quadrifida and C. odorata were not documented in the previous report (Dlamini and Rycroft, 1981). The population of C. odorata was restricted to study sites in Simunye area, but patches of C. odorata were visually observed outside the study areas. The invasion of savannas by C. odorata is a recent phenomenon with episodic potential threat to livestock, game and crop farming as well as the biodiversity conservation in the region.
The present study indicated that A. burkei and D. cinerea may grow on a variety of soil types, but showed preference to sandier soils especially on the raw mineral and lithosol soil. Many encroaching species showed significant differences between the land use systems though without overall consistent trend. Acacia burkei, C. apiculatum, S. caffra and D. cinerea showed the greatest density on communal land. The result implied that the four species are good indicators of disturbance gradient for rangelands subject to long-term over utilization. Although C. apiculatum and D. cinerea were reported to be the commonest encroaching species in Southern Africa rangelands (De Klerk (undated); Annika, 2000), their abundance was not significantly related to land use practices. In any case A. tortilis was shown as one of the encroaching species in east Africa rangelands (Solomon, 2003).
Common Use of Woody Plants
The current study which indicates, that forage utilization of woody plants accounted for the highest contribution agrees with Gemedo et al. (2005) and Solomon et al. (2007b). The woody plants of forage value and not mentioned in earlier study (Dlamini and Rycroft, 1981) were: E. obtusifolia, P. quadrifida, unidentified species locally named as Yaweyawe, Ingogola, Umlahlantfwana, Ndlebetendlovu and Sinukane. Woody plants browsed by three livestock species included D. cinerea, P. quadrifida, L. javanica, B. zeyheri, C. erythruphyllum, B. galpini, S. spinosa and C. apiculatum. The foliage of woody browse and leaf succulent plants appeared to be an important food resource for goats particularly where the availability of forbs is limited (Hendricks et al., 2002). Empirical evidence in Southern Africa suggested that, cattle diets showed significant fraction of browse with an annual mean of 20% (Fritz et al., 1996). The current study also ranked medicinal value of woody plants next to forage utilization. In contrast, medicinal value is the largest plant use for most of the rural people in South America (Coe and Anderson, 1999; Rossato et al., 1999). In East Africa, Solomon et al. (2007b) ranked medicinal value next to forage and food utilization. Therefore, plants used for this purpose may play critical role in the development of sustainable health practices.
Wild edible plants constitute 19% of the total species in the present study and were given a fourth rank. Ziziphys, Euclea, Lantana and Sclerocarya species were similarly reported as edible plants elsewhere in Africa (Gemedo-Dalle et al., 2005; Solomon et al., 2007b). Nutritional analysis of a few wild plants in the Sahel revealed protein, vitamin and mineral contents to be comparable to the WHO standards (FAO, 1986).
CONCLUSIONS
This study concludes that bush encroachment showed non-to-significant variations at small scale level between land use systems and soil classes. Great variability at large scale level was also evident. At small scale level significant differences between the land use systems occurred in severely encroached area and past as well as present grazing pressure could be the major attributing factors to promote bush encroachment and cause differences between the land uses. This is particularly the case where the communal grazing land had more encroachment and the other land use systems manifested lower densities but beyond a threshold condition. Differences between soil classes were due to their inherent diversity and nature. The present study suggests in particular that generalized statements about the relatively bad condition of any land use system and the permanence of any specific condition must be treated with caution. External factors such as expansion of cultivation and land alienation inappropriate to the communal grazing may be added factors to indirectly promote encroachment of woody plants, but scientific examination to validate this theory requires long-term investigation. Furthermore, the most abundant size classes of wood plants (>0.5-2 m) are within the range vulnerable to fire, but also a sign of lasting increase. In general, although bush encroachment is a slow process which limits the present study, future investigation must confirm the findings and be expanded to include various soil types, landscape and rainfall patterns. Woody plants identified as browse and medicinal require research attention as forage and pharmaceutical resources while nutritional assessment and other indigenous knowledge of the wild plants consumed by the rural people remain essentially imperative.
ACKNOWLEDGMENTS
The authors are grateful to the UNISWA Research Board of the University of Swaziland for funding the project. The Ministry of Agriculture and cooperation and the National Trust Commission of Swaziland are fully acknowledged for using their ranches and game reserves for the study. We also express our appreciation to all staff members of Nisella safaris for their assistance and hospitability as well as the communal people in Simunye area for their active participation.
REFERENCES
- Abule, E., G.N. Smit and H.A. Snyman, 2005. The influence of woody plants and livestock grazing on grass species composition, yield and soil nutrients in the Middle Awash Valley of Ethiopia. J. Arid Environ., 60: 343-358.
Direct Link - Annika, C.D., 2000. Vegetation density and change in relation to land use, soil and rainfall: A case study from North-East District, Botswana. J. Arid Environ., 44: 19-40.
Direct Link - Anthony, J.M., H. Kevin, M.S. Rogers and T.F. Witkowski, 2006. A framework for exploring the determinants of savanna and grassland distribution. BioScience, 56: 579-589.
Direct Link - Dlamini, B.J., G.Z. Khumalo and B.B. Xaba, 2000. Cattle performance and nutritive value of pastures on Swazi Nation Land. J. Agric. Sci. Technol., 3: 38-44.
Direct Link - Epstein, H.E., R.A. Gill, J.M. Paruelo, W.K. Lauenroth, G.J. Jia and I.C. Burke, 2002. The relative abundance of three plant functional types in temperate grasslands and shrublands of North and South America: Effects of projected climate change. J. Biol. Geogr., 29: 875-888.
Direct Link - Gemedo-Dalle, T., L.M. Brigitte and I. Johannes, 2005. Plant biodiversity and ethnobotany of Borana pastoralists in Southern Oromia, Ethiopia. Econ Bot., 59: 43-65.
Direct Link - Hudak, A.T. and C.A. Wessman, 2001. Textural analysis of high-resolution imagery to quantify bush encroachment in Madikwe Game Reserve, South Africa, 1955-1996. Int. J. Remote Sens., 22: 2731-2740.
Direct Link - Hudak, A.T., C.A. Wessman and T.R. Seastedt, 2003. Woody overstorey effects on soil carbon and nitrogen pools in South African savanna. Aust. Ecol., 28: 173-181.
Direct Link - Monadjem, A. and K.G. David, 2005. Nesting distribution of vultures in relation to land use in Swaziland. Biodegrad. Conservat., 14: 2079-2093.
CrossRefDirect Link - Oba, G., E. Post, P., P.O. Syvertsen and N.C. Stenseth, 2000. Bush cover and range condition assessments in relation to landscape and grazing in Southern Ethiopia. Landscape Ecol., 15: 535-546.
CrossRefDirect Link - Radford, I.J., D.M. Nicholas, J.R. Brown and D.J. Kriticos, 2001. Paddock-scale patterns of seed production and dispersal in the invasive shrub Acacia nilotica Mimosaceae in Northern Australian rangelands. Aust. Ecol., 26: 338-348.
CrossRefDirect Link - Richter, C.G.F., H.A. Snyman and G.N. Smit, 2001. The influence of tree density on the grass layer of three semi-arid savanna types of Southern Africa. Afr. J. Range Forage Sci., 18: 103-109.
Direct Link - Satti, P., M.J. Mazzarino, M. Obbi, F. Funes, L. Rosell and H. Fernandez, 2003. Soil N dynamics in relation to leaf litter quality and soil fertility in North-Western Patagonian forests. J. Ecol., 91: 173-181.
Direct Link - Smet, M. and D. Ward, 2005. A comparison of the effects of different rangeland management systems on plant species composition, diversity and vegetation structure in a semi-arid savanna. Afr. J. Range Forage Sci., 22: 59-71.
CrossRefDirect Link - Solomon, T., H.A. Snyman and G.N. Smit, 2007. Rangeland dynamics of Southern Ethiopia: (2) Assessment of woody vegetation structure in relation to land use and distance from water in semi-arid Borana rangelands. J. Environ. Manage., 85: 443-452.
Direct Link - Solomon, T.B., H.A. Snyman and G.N. Smit, 2007. Cattle-rangeland management practices and perceptions of pastoralists towards rangeland degradation in the Borana zone of Southern Ethiopia. J. Environ. Manage., 82: 481-494.
CrossRefDirect Link - Tews, J., F. Schur and F. Jeltsch, 2004. Seed dispersal by cattle may cause shrub encroachment of Grewia flava on Southern Kalahari rangelands. Applied Vegtation Sci., 7: 89-102.
Direct Link - Van Auken, O.W., 2000. Shrub invasion of semiarid grasslands. Ann. Rev. Ecol. Syst., 31: 197-215.
Direct Link - Wiegand, K., D. Salts and D. Ward, 2006. A patch-dynamics approach to savanna dynamics and woody plant encroachment-insights from an arid savanna. Perspect. Plant Econ. Evol. Syst., 7: 229-242.
Direct Link