
N. Nejat
Institute of Tropical Agriculture, Universiti Putra Malaysia, 43400, Malaysia
G. Vadamalai
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400, Malaysia
Plant Pathology Journal
Year: 2010 | Volume: 9 | Issue: 3 | Page No.: 112-121







ABSTRACT
Phytoplasmas are small bacteria with very small genomes which also have extremely low levels of the nucleotides guanine plus cytosine (G+C). They are associated with hundreds of plant diseases globally. The uneven distribution and low concentration of phytoplasmas in the phloem of infected plant, especially in woody hosts, and variations in titre according to season and plant organ are also important obstacles for efficient diagnosis. Polymerase Chain Reaction (PCR), nested PCR and real-time PCR have been employed for phytoplasma detection. PCR is the most versatile tool for detecting phytoplasmas in their plant and insect hosts. Nested PCR with a combination of different universal primers can improve the diagnosis of unknown phytoplasmas present with low titre in the symptomatic host. Universal ribosomal primers nested with group-specific primers are extremely useful when the phytoplasma to be diagnosed belongs to a well-defined taxonomic group. Real-time PCR has been shown to be an effective method of quantifying the titre of phytoplasmas within the plant. This paper also discuss phytoplasma diseases on coconut palm.
PDF Abstract XML References Citation
Received: May 10, 2010;
Accepted: June 03, 2010;
Published: September 02, 2010
How to cite this article
N. Nejat and G. Vadamalai, 2010. Phytoplasma Detection in Coconut Palm and Other Tropical Crops. Plant Pathology Journal, 9: 112-121.
DOI: 10.3923/ppj.2010.112.121
URL: https://scialert.net/abstract/?doi=ppj.2010.112.121
DOI: 10.3923/ppj.2010.112.121
URL: https://scialert.net/abstract/?doi=ppj.2010.112.121
INTRODUCTION
Phytoplasmas, formerly termed mycoplasma-like organisms (MLOs), were first discovered in the early 1900s (Kunkel, 1926). They are small bacteria with very small genomes which also have extremely low levels of the nucleotides guanine plus cytosine (G+C), sometimes as little as 23-29% which is thought to be the threshold for a viable genome and coding DNA that are associated with hundreds of plant diseases globally, affecting hundreds of plant species including many important food, vegetable and fruit crops; ornamental plants; timber and shade trees. Geographically, the occurrence of phytoplasmas is worldwide (Lee et al., 2000; Bertaccini, 2007; Dickinson, 2003; Hogenhout et al., 2008).
Phytoplasmas require a vector to be transmitted from plant to plant. They are phloem-limited bacterial pathogens that can cause devastating losses in crops and natural ecosystems worldwide and most prevalent in tropical and sub-tropical regions of the world (Lee et al., 2000; Bertaccini, 2007). Phytoplasmas are minute cell-wall-less prokaryotes with a diameter less than 1 micrometer ranging from 200 to 800 μm, or roughly the size of a plant cell’s chloroplasts, a rounded pleiomorphic or filamentous shape surrounded by a triple layered unit membrane and have cytoplasma, ribosome and strands of nuclear material, DNA is free in the cytoplasm (Bertamini et al., 2003). They arose from a gram-positive clostridiumlike bacterial ancestor of the lactobacillus lineage which appear to have suffered extreme genome reductions compared with their Gram-positive relatives.
The sizes of phytoplasma genomes vary considerably, ranging from 530 to 1350 kbp. The Bermudagrass white leaf phytoplasma has a genome size of just 530 kb, the smallest chromosome of any known living organism to date. The aster yellows phytoplasmas genomes, which are the largest (≤1185 kbp) among the phytoplasmas, are still much smaller than those of culturable relatives (Acholeplasma sp.) (Lee et al., 2000).
Phytoplasmas cause a wide variety of symptoms that ranges from mild yellowing to death of infected plants that suggest profound disturbances in the normal balance of plant hormones or growth regulators (Lee et al., 2000). A common symptom caused by phytoplasma infection include general yellowing or reddening of leaves, stunting, sterility of the flowers, bushy or witche's broom, proliferation of auxiliary shoots, reduced yields, increase in size of the internodes and necrosis of the phloem tissues. Some symptoms fairly unique to phytoplasma diseases such as phyllody, the production of leaf like structures in place of flowers and virescence (the development of green flowers and the loss of normal flower pigments) (Lee et al., 2000; Welliver, 1999).
Phytoplasma detection: Diagnosis of phytoplasmas is routinely done by PCR and can be divided into three phases: total DNA extraction from symptomatic tissue; PCR amplification of phytoplasma-specific DNA; characterization of the amplified DNA by sequencing, RFLP analysis or nested PCR with group-specific primers (Marzachi, 2004). The success of these protocols relies on obtaining nucleic acid preparations of good quality, if possible enriched in phytoplasma DNA. The titre of phytoplasma cells in the phloem of infected plants varies according to the season, plant organ and the plant species and it is often very low in woody hosts. For these reasons, although phytoplasmas seriously threaten the cultivation of some very important crop species, their diagnosis is not simple (Firrao et al., 2007). Thus several methods have been developed to extract total phytoplasma DNA for the detection of phytoplasmas, aiming to concentrate it while reducing plant phenolics and polysaccharides (Ahrens and Seemuller, 1992; Daire et al., 1997; Green et al., 1999; Marzachi et al., 2000; Prince et al., 1993; Zhang et al., 1998). The main goal of each protocol is to concentrate phytoplasma DNA whilst decreasing plant polysaccharide and polyphenolic molecules that can inhibit the Taq DNA polymerase used in the PCR assay. Amplification will not occur if DNA preparations contain PCR-inhibiting substances. Most of the protocols involve a phytoplasma enrichment step followed by total DNA extraction, but some researchers also suggest addition of a proteinase digestion step. The Cetyl Trimethyl Ammonium Bromide (CTAB) method of Doyle and Doyle (1990), phytoplasma enrichment method of Kirkpatrick et al. (1987) and small scale DNA extraction method of Zhang et al. (1998) have been used to extract total nucleic acids from infected coconut tissues (Harrison et al., 2002a; Llauger et al., 2002; Sharmila et al., 2004; Warokka et al., 2006).
DNA extraction methods of a modification of an MLO-enrichment procedure described by Kirkpatrick et al. (1987) and Ahrens and Seemuller (1992), the Cetyl Trimethyl Ammonium Bromide (CTAB) method of Doyle and Doyle (1990) and the small scale procedure (Zhang et al., 1998) with some modification provided a good template for DNA amplification of phytoplasma (Nejat et al., 2009a; Warokka et al., 2006). On the other hand, the use of the small scale method is much faster and more convenient than the MLO-enrichment procedure for DNA extraction from plant tissue and insect vectors. This method also significantly decreased the amount of the tubes, reagents and chemicals and is relatively inexpensive compared to the MLO-enrichment procedure and commercially available nucleic acid extraction kits (Zhang et al., 1998).
The amount of phytoplasma DNA recovered from total DNA was significantly affected by the plant part used for DNA extraction (Kollar et al., 1990). Furthermore, the known low concentrations and uneven distribution of the phytoplasmas in the phloem vessels of infected plants increases the dependence of detection on the appropriately chosen material (Davis and Lee, 1993; Jones et al., 1999; Seemüller et al., 1998). Detection of LY phytoplasma by PCR has indicated highest sensitivity when the DNA sample has been extracted from immature palm leaf bases (heart tissues) rich in functional phloem (Harrison et al., 1994). Detection of LY by PCR employing ribosomal or non-ribosomal primer pairs has also been obtained with the DNA sample that has been extracted from leaves or inflorescence, trunk tissues and roots of coconut palm (Harrison et al., 1999). Such studies have also revealed that the LY phytoplasmas are more readily detected in immature than mature tissues (Escamilla et al., 1995; Harrison et al., 1995). Mpunami (1997) reported that phytoplasma DNA was detected in all meristematic tissues sampled, including the area below the growing point, the spear leaf, the petioles of very young leaves, inflorescences and root tips. This is in agreement with Warokka (1999) who showed that the highest concentrations were found in the area below the growing point, the petioles of young unopened leaves and the root tips. Therefore, the inability to locate the pathogen in mature leaves reflects perhaps an irregular distribution of phytoplasma rather than its absence from these organs (Harrison et al., 1994). The Coconut Yellow Decline (CYD) phytoplasma detection was consistent and reproducible for the affected coconut palm ecotypes when template DNAs for PCR were derived from either spear leaves, inflorescences or trunk tissues (Nejat et al., 2009b).
Since, phytoplasmas occur in low titre in the phloem tissues of their host-plants and their concentration may be subject to seasonal fluctuations. In woody plants, it is known that the persistence of phytoplasma in the aerial parts depends on the seasonal state of the phloem (Braun and Sinclair, 1976). The phytoplasmas overwinter in the root system and recolonize the stem in spring. In pears and apples, phytoplasmas disappeared from the above-ground parts of the trees due to cold temperatures during the winter months and that they survived in the roots of the scion to recolonize the stem and branches in the following spring (Seemuller et al., 1984). The reason behind this behaviour is thought to be the degeneration and complete inactivation of sieve tubes in the above ground parts of trees in winter (Evert, 1960; Jarausch et al., 1999; Schaper and Seemüller, 1982). Seemuller et al. (1988) reported that the highest level of phytoplasma population in proliferation-diseased apple trees was reached in September or October in the young shoots and trunks, using fluorescence microscopy using DAPI.
The titre of phytoplasma in coconuts also depends on environmental and climatic conditions in Malaysia. The lower temperature and higher humidity in the wet season might allow circulation through phloem into the aerial parts of the tree to continue for longer. It may also reflect an increase in the concentration and the even distribution of phytoplasma in phloem tissue. As a consequence, CYD could continue to be distributed through the tree, thus enhancing the possibility of detection in December, without necessarily indicating an increase in CYD concentration.
Detection and identification of phytoplasmas is necessary for accurate disease diagnosis. Molecular diagnostic techniques for the detection of phytoplasma introduced during the last two decades have proven to be more accurate and reliable than biological criteria long used for phytoplasma identification (Lee et al., 2000). Polymerase Chain Reaction (PCR) is the most versatile tool for detecting phytoplasmas in their plant and insect hosts (Smart et al., 1996). Phytoplasma diagnostics has been routinely based on phytoplasma-specific universal (generic) (Table 1) or phytoplasma group specific Polymerase Chain Reaction (PCR) primers designed on the basis of the highly conserved 16S ribosomal RNA (rRNA) gene sequences (Ahrens and Seeuller, 1992; Davis and Lee, 1993; Deng and Hiruki, 1991a, b; Harrison et al., 1996; Jomantiene et al., 1998; Schaff et al., 1992; Smart et al., 1996). Phytoplasma group-specific primers have also been designed on ribosomal protein, SecA, SecY and elongation factor genes sequences (Table 2) (Gundersen et al., 1996; Hodgetts et al., 2008; Lee et al., 2006; Marcone et al., 2000; Schneider et al., 1997) and alternative diagnostic methods based on other gene sequences have been established, such as Heteroduplex Mobility Assays (HMAs) (Wang and Hiruki, 2005), Single Stand Conformation Polymorphisms (SSCP) (Music et al., 2008), Terminal Restriction Fragment Length Polymorphism (T-RFLP) (Hodgetts et al., 2007) and real-time PCR (Baric and Dalla-Via, 2004; Hren et al., 2007; Wei et al., 2004).
For sensitive detection of phytoplasmas in plants with very low levels of inoculum, specific primers based on Mollicutes 16S rRNA genes have been used to selectively amplify phytoplasma DNA from mixtures with host DNA (Ahrens and Seemüller, 1992; Davis et al., 1992; Deng and Hiruki, 1990, 1991a, b; Lee et al., 1993; Namba et al., 1993). Proof of phytoplasma association can be obtained by several DNA-based techniques including PCR, nested PCR and recently, real-time PCR for routine disease diagnosis and epidemiological studies. Generally, the 16S rDNA is amplified by universal primer pair P1/P7 followed by nested PCR with R16F2n/R16R2 primer pair (Deng and Hiruki, 1991b; Gundersen and Lee, 1996; Lee et al., 1993; Schneider et al., 1995). The choice of primer sets for phytoplasma diagnosis by nested PCR mostly depends on the phytoplasma we are looking for. Nested PCR with a combination of different universal primers can improve the diagnosis of unknown phytoplasmas present with low titre in the symptomatic host. Universal ribosomal primers nested with group-specific primers are extremely useful when the phytoplasma to be diagnosed belongs to a well-defined taxonomic group (Marzachi, 2004).
Unfortunately, some primers can induce dimers or unspecific bands. They also have sequence homology in the 16S-spacer region to chloroplasts and plastids increasing the risk of false positives (Heinrich et al., 2001). The universal phytoplasma primer pair, R16F2n/R16R2 amplified the 16S rRNA gene from coconut chloroplast derived from spear leaf and inflorescence samples of some asymptomatic coconut palms. These results indicate that R16F2n/R16R2 can amplify plant DNA. Secondly, Bacillus sp., were amplified with these primers from the inflorescences of some asymptomatic coconut palms.
| Table 1: | PCR general primers commonly used for the detection of phytoplasma |
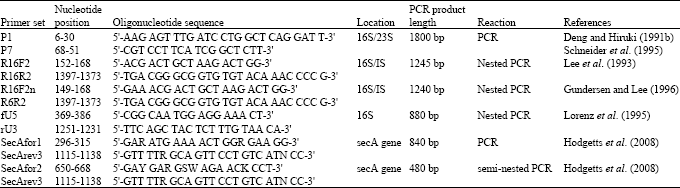 | |
| Table 2: | Several group specific primers used for phytoplasma detection |
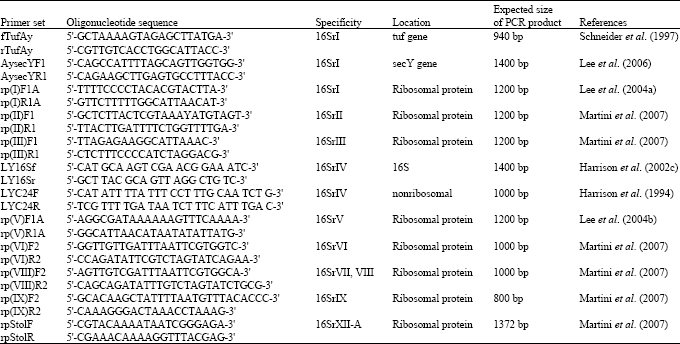 | |
| Table 3: | Oligonucleotide primers and TaqMan probes sequences used for phytoplasma detection by real-time PCR (Galetto and Marzachi, 2010) |
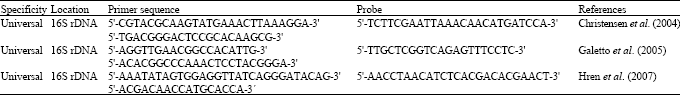 | |
Bacillus megaterium has previously been isolated from the trunk samples of date palms (Phoenix canariensis Chabaud) affected by the lethal decline phytoplasma using universal phytoplasma primer pair (P1/P7) in Texas (Harrison et al., 2002c) more specific universal phytoplasma primers are currently being developed (Hodgetts et al., 2007, 2008; Martini et al., 2007) and it may be that these will be more suitable for diagnostics from samples such as coconuts which appear to harbor other Gram-positive bacteria in significant populations particularly on the spear leaves and inflorescences. Primers fU5/rU3 were used in nested PCR more reliable to detect from the spear leaves and inflorescences of palms (Nejat et al., 2009c).
The use of nested PCR has been reported for diagnostic purposes particularly in trees and insect vectors when phytoplasmas occur in low titre in the phloem vessels of their host-plants and their concentration may be subjected to seasonal fluctuation (Gundersen and Lee, 1996; Jacobs et al., 2003; Leyva-Lopez et al., 2002; Marzachi, 2004).
The PCR program using parameters as described by Lee et al. (1993), using 1 min (2 min for an initial denaturation) at 94°C, 2 min at 55°C and 3 min at 72°C for 35 cycles and a final extension at 72°C for 10 min is time-comsuming require about 9 h for nest PCR. The following PCR program is shorter: 35 cycles of 1 min at 94°C, 1 min at 57°C and 90 sec at 72°C with a final elongation of 10 min at 72°C.
Real-time PCR method has many advantages over the conventional PCR in terms of accuracy, dynamic range, short analysis time, high automation capability, high-throughput capacity and absence of post-PCR manipulations that prevents carryover contamination (Higuchi et al., 1993; Schaad and Fredrick, 2002). The application of this method to plant pathogens is increasing and in the case of phytoplasmas real-time PCR has been up to now applied for both phytoplasmas detection and quantification. Real-time PCR has been shown to be effective methods of quantifying the titre of phytoplasmas within the plant.
Recently three protocols for the universal diagnosis of phytoplasmas using direct real-time PCR amplification of the 16S rDNA gene have been developed (Christensen et al., 2004; Galetto et al., 2005; Hren et al., 2007). Real-time PCR have been developed for diagnosis Flavescence Dorée (FD) and Bois Noir (BN) phytoplasmas infecting grapevines, Columbia Basin potato purple top and CYD novel phytoplasma infecting Malayan yellow and red dwarf ecotypes (Aldaghi et al., 2007; Baric and Dalla-Via, 2004; Baric et al., 2006; Crosslin et al., 2006; Jarausch et al., 2004; Nejat et al., 2010).
Identification: The highly conserved 16S rRNA gene sequence has been widely used as the very useful primary molecular tool for preliminary classification of phytoplasmas. A total of 19 distinct groups, termed 16S rRNA groups (16Sr groups), based on actual RFLP analysis of PCR-amplified 16S rDNA sequences or 29 groups based on in silico RFLP analysis have been identified (Lee et al., 1998, Wei et al., 2007).
Phylogeny based on 16S ribosomal DNA (16Sr) sequences divides the phytoplasmas into three distinct clusters (Hogenhout et al., 2008). The first cluster (Cluster I) contains the Aster yellows (AY) 16SrI group and the stolbur (STOL) 16SrXII group phytoplasmas. These two groups have diverged but are clearly more closely related to each other than the other phytoplasma groups (Hogenhout et al., 2008). The second cluster (Cluster II) contains the Apple Proliferation (AP) 16SrX group phytoplasmas and the third cluster (Cluster III) contains the largest number of phytoplasma groups, including Western X (WX, 16SrIII), Palm Lethal Yellowing (LY, 16SrIV) and Elm Yellows (EY, 16SrV) (Hogenhout et al., 2008). To date, phytoplasma diseases belonging to 16SrI, 16SrXIV and novel phytoplasma have been identified in Malaysia (Nejat et al., 2009c).
Coconut palm phytoplasma diseases: The coconut palm is a versatile plant with a variety of uses in the world. Every part of it is useful to mankind in form or the other it supplies food, drink, oil and shelter and also raw materials for a number of industries for generations. Coconut cultivation faces a strong phytopathological constraint caused by the lethal yellowing (16SrIV) group phytoplasmas. Lethal yellowing is a highly destructive, fast spreading disease of coconut and at least 35 other palm species. Lethal yellowing in the USA (Harrison et al., 1994), Lethal Disease (LD) in Tanzania, East Africa (Schuiling et al., 1981), Cape St. Paul Wilt (CSPW) in Ghana, West Africa (Johnson and Harries, 1976; Nipah et al., 2007), Kaincope disease in Togo (Nienhaus and Steiner, 1976), Kribi disease in Cameroon (Dollet et al., 1977) and Awka or bronze leaf wilt (LDN) in Nigeria (Ekpo and Ojomo, 1990) are all classified as 16SrIV phytoplasmas. Subgroup 16SrIV-A phytoplasmas are associated with Lethal Yellowing (LY) of coconut and other palm species in the Americas (Harrison et al., 2002a) whereas, phytoplasmas that induce symptoms similar to LY on coconut in Africa are referred to by other names to reflect strain differences that have previously been identified through 16S rRNA gene analysis. Recently, Wei et al. (2007) have allocated the Nigerian coconut lethal decline group (LDN) to a distinct 16Sr group, 16SrXXII-A and this has been confirmed by the work of Hodgetts et al. (2008), who showed a high degree of divergence between the different coconut phytoplasmas based on the secA gene which supported their separation into at least three distinct Ca. Phytoplasma species that reflect the geographical origins of the strains.
Weligama wilt disease of coconut in Sri Lanka is in the 16SrXI Ca. Phytoplasma oryzae group based on its 16S rRNA sequence (ACC. No. EU635503), a group of phytoplasmas commonly found in sugar cane, whilst Kalimantan Wilt (KW) disease of coconuts in Indonesia which belongs to the two phylogenetic groups, 16SrXI Ca. Phytoplasma oryzae and 16SrXIII Mexican periwinkle virescence (Warokka, 2005).
Kerala Wilt Disease (KWD) of coconuts in India which as a different 16S rRNA phytoplasma group phylogenetically (16SrIV-C) (Edwin and Mohankumara, 2007; Sharmila et al., 2004). In Malaysia, a 16SrXIV Ca. Phytoplasma cynodontis phytoplasma was found associated with coconut yellow decline in Malayan Tall and Malayan Red Dwarf ecotypes and a novel phytoplasma was identified in Malayan Yellow dwarf ecotype (Table 4).
Cynodon dactylon L. Pers is commonly called Bermudagrass in many areas of the world. Bermudagrass white leaf (BGWL) is a destructive, phytoplasmal disease of Bermudagrass has been detected in Thaiwan, Pakistan, China, Italy, Iran, Thailand, India and Malaysia.
| Table 4: | Coconut palm phytoplasma diseases |
 | |
| Table 5: | Phytoplasmas related to the Candidatus Phytoplasma cynodontis (16SrXIV) group |
 | |
The causal agent of BGWL is Candidatus Phytoplasma cynodontis (Marcone et al., 2004). In Northern Sudan, a 16SrXIV Ca. Phytoplasma cynodontis phytoplasma was found associated with slow decline and white tip die-back in date palm (Cronje et al., 2000a) (Table 5).
Control of phytoplasma diseases: In controlling phytoplasmal diseases, the primary concern is often prevention rather than treatment. The most promising strategy for avoiding phytoplasma disease is the use of disease resistance varieties of crops. Phytoplasmas are normally controlled by planting disease-free stocks (Propagate from seed, or from phytoplasma-free plants), control of insect vectors and weed plant hosts act as sources of inoculums, Rogue out and destroy symptomatic plants and avoid planting susceptible crops next to crops harboring the phytoplasma (Lee et al., 2000; Welliver, 1999).
REFERENCES
- Aldaghi, M., S. Massart, S. Roussel and M.H. Jijakli, 2007. Development of a new probe for specific and sensitive detection of Candidatus Phytoplasma mali in inoculated apple trees. Ann. Applied Biol., 151: 251-258.
Direct Link - Baric, S. and J. Dalla-Via, 2004. A new approach to apple proliferation detection: A highly sensitive real-time PCR assay. J. Microbiol. Meth., 57: 135-145.
Direct Link - Baric, S., C. Kerschbamer and J. Dalla-Via, 2006. TaqMan real-time PCR versus four conventional PCR assays for detection of apple proliferation phytoplasma. Plant Mol. Biol. Rep., 24: 169-184.
Direct Link - Bertaccini, A., 2007. Phytoplasmas: Diversity, taxonomy and epidemiology. Front Biosci., 12: 673-689.
PubMedDirect Link - Bertamini, M., M.S. Grando and N. Nedunchezhian, 2003. Effects of phytoplasma infection on pigments, chlorophyll-protein complex and photosynthetic activities in field grown apple leaves. Biol. Plant., 47: 237-242.
Direct Link - Braun, E.J. and W.A. Sinclair, 1976. Histopathology of phloem necrosis in Ulnus americana. Phytopathology, 66: 598-607.
Direct Link - Christensen, N.M., M. Nicolaisen, M. Hansen and A. Schulz, 2004. Distribution of phytoplasmas in infected plants as revealed by real-time PCR and bioimaging. Mol. Plant Microbe Interact., 17: 1175-1184.
Direct Link - Cronje, P., A.J. Dabek, P. Jones and A.M. Tymon, 2000. First report of a phytoplasma associated with a disease of date palms in North Africa. Plant Pathol., 49: 801-801.
Direct Link - Cronje, P., A.J. Dabek, P. Jones and A.M. Tymon, 2000. Slow decline: A new disease of mature date palms in North Africa associated with a phytoplasma. Plant Pathol., 49: 804-804.
CrossRef - Crosslin, J.M., G.J. Vandemark and J.E. Munyaneza, 2006. Development of a real-time, quantitative PCR for detection of the columbia basin potato purple top phytoplasma in plants and beet leafhoppers. Plant Dis., 90: 663-667.
CrossRef - Dafalla, G.A. and M.T. Cousin, 1988. Fluorescence and electron microscopy of Cynodon dactylon affected with a white leaf disease in Sudan. J. Phytopathol., 122: 25-34.
Direct Link - Daire, X., D. Clair, W. Reinert and E. Boudon-Padieu, 1997. Detection and differentiation of grapevine yellows phytoplasma belonging to the elm yellows group and to the stolbur subgroup by PCR amplification of non-ribosomal DNA. Eur. J. Plant Pathol., 103: 507-514.
Direct Link - Davis, R.E. and I.M. Lee, 1993. Cluster-specific polymerase chain reaction amplification of 16s rDNA sequences for detection and identification of mycoplasma-like organisms. Phytopathology, 83: 1008-1011.
Direct Link - Deng, S. and C. Hiruki, 1990. Enhanced detection of a plant pathogenic mycoplasma-like organism by polymerase chain reaction. Proc. Jpn. Acad. Ser. B: Phys. Biol. Sci., 66: 140-144.
Direct Link - Deng, S. and C. Hiruki, 1991. Genetic relatedness between two nonculturable mycoplasma-like organisms revealed by nucleic acid hybridization and polymerase chain reaction. Phytopathology, 81: 1475-1479.
Direct Link - Deng, S. and C. Hiruki, 1991. Amplification of 16S rRNA genes from culturable and nonculturable mollicutes. J. Microbiol. Methods, 14: 53-61.
CrossRefDirect Link - Dollet, M., J. Giannotti, J.L. Renard and S.K. Ghosh, 1977. Study of a lethal yellowing of coconuts in cameroon: Kribi Disease. Observation of mycoplasma-like organisms. Oleagineux, 32: 317-322.
Direct Link - Edwin, B.T. and C. Mohankumara, 2007. Kerala wilt disease phytoplasma: Phylogenetic analysis and identification of a vector, Proutista moesta. Physiol. Mol. Plant Pathol., 71: 41-47.
CrossRef - Escamilla, J.A., N.A. Harrison, H. Nunez, L. Alpizar, I. Cordova, I. Islas-Flores and C. Oropeza, 1995. Practical use of DNA Probes for the Detection of Lethal Yellowing of Cocos nucifera L. in Mexico. In: Lethal Yellowing: Research and Practical Aspects, Oropeza, C., F.W. Howard and G.R. Ashburner (Eds.). Kluwer Academic Publishers, Dordecht, Netherlands, pp: 93-99.
- Evert, R.F., 1960. Phloem structure in Pyrus communis L. and its seasonal changes. Univ. Calif. Publ. Bot., 32: 127-194.
Direct Link - Firrao, G., M. Garcia-Chapa and C. Marzachi, 2007. Phytoplasmas: Genetics, diagnosis and relationships with the plant and insect host. Front Biosci., 12: 1353-1375.
PubMedDirect Link - Galetto, L., D. Bosco and C. Marzachi, 2005. Universal and group-specific real-time PCR diagnosis of flavescence doree (16Sr-V), bois noir (16Sr-XII) and apple proliferation (16Sr-X) phytoplasmas from field-collected plant hosts and insect vectors. Ann. Applied Biol., 147: 191-201.
Direct Link - Green, M.J., D.A. Thompson and D.J. MacKenzie, 1999. Easy and efficient DNA extraction from woody plants for the detection of phytoplasmas by polymerase chain reaction. Plant Dis., 83: 482-485.
Direct Link - Gundersen, D.E., I.M. Lee, D.A. Schaff, N.A. Harrison, C.J. Chang, R.E. Davis and D.T. Kingsbury, 1996. Genomic diversity and differentiation among phytoplasma strains in the 16S rRNA groups I (aster yellows and related phytoplasmas) and III (X-disease and related phytoplasmas). Int. J. Syst. Bacteriol., 46: 64-75.
Direct Link - Harrison, N.A., I. Cordova, P.A. Richardson and R. Dibonito, 1999. Detection and Diagnosis of Lethal Yellowing. In: Current Advances in Coconut Biotechnology, Oropeza, C., C.L. Verdeil, G.R. Ashburner, R. Cardena and J.M. Santamaria (Eds.). Kluwer Academic Publishers, Dordrecht, Netherlands, pp: 183-196.
- Harrison, N.A., P.A. Richardson and J.H. Tsai, 1995. Detection and Diagnosis of Lethal Yellowing: Conventional Methods and Molecular Techniques. In: Lethal Yellowing: Research and Practical Aspects, Oropeza, C., F.W. Howard and G.R. Ashburner (Eds.). Kluwer Academic Publishers, Dordrecht, Netherlands, pp: 79-91.
- Harrison, N.A., P.A. Richardson and J.H. Tsai, 1996. PCR assay for detection of the phytoplasma associated with maize bushy stunt disease. Plant Dis., 80: 263-269.
Direct Link - Harrison, N.A., M. Womack and M.L. Carpio, 2002. Detection and characterization of a lethal yellowing (16SrIV) group phytoplasma in Canary Island date palms affected by lethal decline in Texas. Plant Dis., 86: 676-681.
Direct Link - Harrison, N.A., W. Myrie, P. Jones, M.L. Carpio and C. Oropeza, 2002. 16S rRNA interoperon sequence heterogeneity distinguishes strain populations of the palm lethal yellowing phytoplasma in the Caribbean region. Ann. Applied Biol., 141: 183-193.
Direct Link - Heinrich, M., S. Botti, L. Caprara, W. Arthofer and S. Strommer et al., 2001. Improved detection methods for fruit tree phytoplasmas. Plant Mol. Biol. Rep., 19: 169-179.
Direct Link - Higuchi, R., C. Fockler, G. Dollinger and R. Watson, 1993. Kinetic PCR analysis: Real-time monitoring of DNA amplification reactions. Nat. Biotechnol., 11: 1026-1030.
Direct Link - Hodgetts, J., T. Ball, N. Boonham, R. Mumford and M. Dickinson, 2007. Use of terminal restriction fragment length polymorphism (T-RFLP) for identification of phytoplasmas in plants. Plant Pathol., 56: 357-365.
Direct Link - Hodgetts, J., N. Boonham, R. Mumford, N. Harrison and M. Dickinson, 2008. Phytoplasma phylogenetics based on analysis of secA and 23S rRNA gene sequences for improved resolution of candidate species of Candidatus phytoplasma. Int. J. Syst. Evol. Microbiol., 58: 1826-1837.
CrossRefDirect Link - Hogenhout, S.A., K. Oshima, E. Ammar, S. Kakizawa, H.N. Kingdom and S. Namba, 2008. Phytoplasmas: Bacteria that manipulate plants and insects. Mol. Plant Pathol., 9: 403-423.
CrossRefDirect Link - Hren, M., J. Boben, A. Rotter, P. Kralj, K. Gruden and M. Ravnikar, 2007. Real-time PCR detection systems for Flavescence doree and Bois noir phytoplasmas in grapevine: comparison with conventional PCR detection and application in diagnostics. Plant Pathol., 56: 785-796.
Direct Link - Jacobs, K.A., I.M. Lee, H.M. Griffiths, F.D. Miller and K.D. Bottner, 2003. A new member of the clover proliferation phytoplasma group (16SrVI) associated with elm yellows in Illinois. Plant Dis., 87: 241-246.
Direct Link - Jarausch, W., T. Peccerella, B. Schwind, B. Jarausch and G. Krczal, 2004. Establishment of a quantitative real-time PCR assay for the quantification of apple proliferation phytoplasmas in plants and insects. Acta Hort., 657: 415-420.
Direct Link - Jomantiene, R., R.E. Davis, J. Maas and E.L. Dally, 1998. Classification of new phytoplasmas associated with diseases of strawberry in Florida, based on analysis of 16S rRNA and ribosomal protein gene operon sequences. Int. J. Syst. Bacteriol., 48: 269-277.
Direct Link - Kirkpatrick, B.C., D.C. Stenger, T.J. Morris and A.H. Purcell, 1987. Cloning and detection of DNA from a nonculturable plant pathogenic mycoplasma-like organism. Science, 238: 197-200.
CrossRef - Kollar, A., E. Seemuller, F. Bonnet, C. Saillard and J.M. Bove, 1990. Isolation of the DNA of various plant pathogenic mycoplasma-like organisms from infected plants. Phytopathology, 80: 233-237.
Direct Link - Lee, I.M., R.E. Davis and D.E. Gundersen-Rindal, 2000. Phytoplasma: Phytopathogenic mollicutes. Ann. Rev. Microbiol., 54: 221-555.
CrossRefPubMedDirect Link - Lee, I.M., D.E. Gundersen-Rindal, R.E. Davis and I.M. Bartoszyk, 1998. Revised classification scheme of phytoplasma based on RFLP analyses of 16S rRNA and ribosomal protein gene sequences. Int. J. Syst. Bacteriol., 48: 1153-1169.
CrossRef - Lee, I.M., R.W. Hammond, R.E. Davis and D.E. Gundersen, 1993. Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasma-like organisms. Phytopathology, 83: 834-842.
Direct Link - Lee, I.M., D.E. Gundersen-Rindal, R.E. Davis, K.D. Bottner, C. Marcone and E. Seemuller, 2004. Candidatus Phytoplasma asteris, a novel phytoplasma taxon associated with aster yellows and related diseases. Int. J. Syst. Evol. Microbiol., 54: 1037-1048.
CrossRef - Lee, I.M., M. Martini, C. Marcone and S.F. Zhu, 2004. Classification of phytoplasma strains in the elm yellows group (16SrV) and proposal of Candidatus Phytoplasma ulmi for the phytoplasma associated with elm yellows. Int. J. Syst. Evol. Microbiol., 54: 337-347.
Direct Link - Lee, I.M., Y. Zhao and K.D. Bottner, 2006. SecY gene sequence analysis for finer differentiation of diverse strains in the aster yellows phytoplasma group. Mol. Cell. Prob., 20: 87-91.
CrossRef - Llauger, R., D. Becker, J. Cueto, E. Peralta, V. Gonzalez, M. Rodriguez and W. Rohde, 2002. Detection and molecular characterization of phytoplasma associated with Lethal yellowing disease of coconut palms in Cuba. J. Phytopathol., 150: 390-395.
Direct Link - Lorenz, K.H., B. Schneider, U. Ahrens and E. Seemuller, 1995. Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and nonribosomal DNA. Phytopathology, 85: 771-776.
Direct Link - Marcone, C., B. Schneider and E. Seemuller, 2004. Candidatus Phytoplasma cynodontis, the phytoplasma associated with Bermuda grass white leaf disease. Int. J. Syst. Evol. Microbiol., 54: 1077-1082.
CrossRef - Marcone, C., I.M. Lee, R.E. Davis, A. Ragozzino and E. Seemuller, 2000. Classification of aster yellows-group phytoplasmas based on combined analyses of rRNA and tuf gene sequences. Int. J. Syst. Evol. Microbiol., 50: 1703-1713.
Direct Link - Marcone, C., A. Ragozzino and E. Seemuller, 1997. Detection of Bermuda grass white leaf disease in Italy and characterization of the associated phytoplasma by RFLP analysis. Plant Dis., 81: 862-866.
CrossRef - Martini, M., I.M. Lee, K.D. Bottner, Y. Zhao and S. Botti et al., 2007. Ribosomal protein gene-based phylogeny for finer differentiation and classification of phytoplasmas. Int. J. Syst. Evol. Microbiol., 57: 2037-2051.
CrossRef - Marzachi, C., 2004. Molecular diagnosis of phytoplasmas. Phytopathol. Mediterr., 43: 228-231.
Direct Link - Marzachi, C., F. Veratti, M. d'Aquilio, A. Vischi, M. Conti and G. Boccardo, 2000. Molecular hybridisation and PCR amplification of non-ribosomal DNA to detect and differentiate stolbur phytoplasma isolates from Italy. J. Plant Pathol., 82: 201-212.
Direct Link - Music, M.S., M. Krajacic and D. Skoric, 2008. The use of SSCP analysis in the assessment of phytoplasma gene variability. J. Microbiol. Methods, 73: 69-72.
CrossRef - Namba, S., S. Kato, S. Iwanami, H. Oyaizu, H. Shiozawa and T. Tsuchizaki, 1993. Detection and differentiation of plant-pathogenic mycoplasma-like organisms using polymerase chain reaction. Phytopathology, 83: 786-791.
CrossRef - Nejat, N., K. Sijam, S.N.A. Abdullah, G. Vadamalai and M. Dickinson, 2009. First report of a 16SrXIV, Candidatus phytoplasma cynodontis group phytoplasma associated with coconut yellow decline in Malaysia. Plant Pathol., 58: 389-389.
CrossRef - Nejat, N., K. Sijam, S.N.A. Abdullah, G. Vadamalai and M. Dickinson, 2009. Molecular characterization of a phytoplasma associated with coconut yellow decline (CYD) in Malaysia. Am. J. Applied Sci., 6: 1331-1340.
Direct Link - Nejat, N., K. Sijam, S.N.A. Abdullah, G. Vadamalai and M. Dickinson, 2009. Phytoplasmas associated with disease of coconut in Malaysia are from two different phylogenetic groups and occur in other plant species. Plant Pathol., 58: 1152-1160.
Direct Link - Nipah, J.O., P. Jones and M.J. Dickinson, 2007. Detection of lethal yellowing phytoplasma in embryos from coconut palms infected with Cape St Paul wilt disease in Ghana. Plant Pathol., 56: 777-784.
Direct Link - Prince, J.P., R.E. Davis, T.K. Wolf, I.M. Lee and B. Mogen et al., 1993. Molecular detection of diverse Mycoplasma-Like Organisms (MLOs) associated with grapevine yellows and their classification with aster yellows, X-disease and elm yellows MLOs. Phytopathology, 83: 1130-1137.
Direct Link - Schaff, D., I.M. Lee and R.E. Davis, 1992. Sensitive detection and identification of mycoplasma-like organisms by polymerase chain reactions. Biochem. Biophys. Res. Commun., 186: 1503-1509.
PubMed - Schaper, U. and E. Seemuller, 1982. Conditions of the phloem and the persistence of mycoplasma-like organisms associated with apple proliferation and pear decline. Phytopathology, 72: 736-742.
Direct Link - Schneider, B., A. Padovan, S. De La Rue, R. Eichner, R. Davis, A. Bernuetz and K. Gibb, 1999. Detection and differentiation of phytoplasmas in Australia: An update. Aust. J. Agric. Res., 50: 333-342.
Direct Link - Schneider, B. and K.S. Gibb, 1997. Sequence and RFLP analysis of the elongation factor Tu gene used in differentiation and classification of phytoplasmas. Microbiology, 143: 3381-3389.
CrossRefDirect Link - Spoodee, R., B.L. Schneider, A.C. Padovan and K.S. Gibb, 1999. Detection and genetic relatedness of phytoplasmas associated with plant diseases in Thailand. J. Biochem. Mol. Biol. Biophys., 3: 133-140.
Direct Link - Seemuller, E.C., U. Marcone, A. Lauer, A. Ragozzino and M. Goschl, 1998. Current status of molecular classification of the phytoplasma. J. Plant Pathol., 80: 3-26.
CrossRefDirect Link - Sharmila, L.B., S. Bhasker, M.T. Thelly, T.E. Boby and C. Mohankumar, 2004. Cloning and sequencing of phytoplasma ribosomal DNA (rDNA) associated with Kerala wilt disease of coconut palms. J. Plant Biochem. Biotechnol., 13: 1-5.
Direct Link - Smart, C.D., B. Schneider, C.L. Blomquist, L.J. Guerra and N.A. Harrison et al., 1996. Phytoplasma-specific PCR primers based on sequences of the 16S-23S rRNA spacer region. Phytopathol., 62: 2988-2993.
PubMedDirect Link - Snehi, S.K., M.S. Khan, S.K. Raj, S. Mall, M. Singh and G.P. Rao, 2008. Molecular identification of Candidatus phytoplasma cynodontis associated with bermuda grass white leaf disease in India. Plant Pathol., 57: 770-770.
CrossRef - Tymon, A.M., P. Jones and N.A. Harrison, 1998. Phylogenetic relationships of coconut phytoplasmas and the development of specific oligonucleotide PCR primers. Ann. Applied Biol., 132: 437-452.
Direct Link - Wang, K. and C. Hiruki, 2005. Distinctions between phytoplasmas at the subgroup level detected by heteroduplex mobility assay. Plant Pathol., 54: 625-633.
Direct Link - Wei, W., R.E. Davis, I.M. Lee and Y. Zhao, 2007. Computer-simulated RFLP analysis of 16S rRNA genes: Identification of ten new phytoplasma groups. Int. J. Syst. Evol. Microbiol., 57: 1855-1867.
CrossRefPubMedDirect Link - Wei, W., S. Kakizawa, S. Suzuki, H.Y. Jung and H. Nishigawa et al., 2004. In planta dynamic analysis of onion yellows phytoplasma using localized inoculation by insect transmission. Phytopathology, 94: 244-250.
CrossRef - Zahoor, A., M. Bashir, K. Nakashima, T. Mitsueda and N. Murata, 1995. Bermuda grass white leaf caused by phytoplasmas in Pakistan. Pak. J. Bot., 27: 251-252.
Direct Link - Zhang, Y., J.K. Uyemoto and B.C. Kirkpatrick, 1998. A small-scale procedure for extracting nucleic acids from woody plants infected with various phytopathogens for PCR assay. J. Virol. Methods, 71: 45-50.
CrossRefDirect Link