
Hadi Ahmadi
Agricultural Faculty, University of Lorestan, P.O. Box 465, Khoramabad, Iran
K. Moore
Department of Agricultural and Environment Science, The University,
Newcastle Upon Tyne NEl 7RU, UK
Plant Pathology Journal
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 164-168







ABSTRACT
Powdery mildew of wheat is caused by Erysiphe graminis f. sp. tritici and it is a major disease of wheat (Triticum aestivum L.). Therefore there is a need for more research to find new genes for resistance. To study the inheritance of gene/s for resistance to powdery mildew three susceptible T. durum varieties were crossed with three resistant T. dicoccoides accessions. The segregation results in F1, F2, F3 and reciprocal BC1 progenies confirmed that T. dicoccoides accessions TA1055 and TA1150 possess one similar dominant gene for resistance to Erysiphe graminis f. sp. tritici To determine chromosomal location of gene for resistance to powdery mildew 14 monosomic lines of Chinese Spring were used (1A-7A and 1B-7B) to cross as female with the resistant accessions TA1055 and TA1150. The F1 hybrid seeds were germinated to obtain F2 seeds. Analysis of obtained data revealed that one major dominant gene conferring resistance is located on chromosome 2A of T. dicoccoides accession TA1055 which is different from 33 current known genes for resistance to wheat powdery mildew and should be designated Pmtd1055.
PDF Abstract XML References
How to cite this article
Hadi Ahmadi and K. Moore, 2007. Inheritance and Chromosomal Location of Powdery Mildew Resistance Gene in Wild Wheat Triticum turgidum Var. dicoccoides. Plant Pathology Journal, 6: 164-168.
DOI: 10.3923/ppj.2007.164.168
URL: https://scialert.net/abstract/?doi=ppj.2007.164.168
DOI: 10.3923/ppj.2007.164.168
URL: https://scialert.net/abstract/?doi=ppj.2007.164.168
INTRODUCTION
Powdery mildew is caused by Erysiphe graminis f. sp. tritici and it is one of the most important disease of wheat in arias with maritime or semicontinental climate (Bennett, 1984). Up to now, 51 Pm alleles at 33 loci have been identified for wheat powdery mildew resistance (McIntosh et al., 2005). Thirty alleles at 24 loci from Pm1 to Pm25, their locations on chromosomes and their sources have been reviewed, other Pm genes, Pm26, Pm27 and Pm28, Pm29, Pm30, Pm31 and Pm32 have also been reported (Shi et al., 1998; Jarve et al., 2000; Peusha et al., 2000; Rong et al., 2000; Zeller et al., 2002; Liu et al., 2002; Xie et al., 2003; Hsam et al., 2003).
Pm1, Pm2, Pm3 (a-f), Pm9, Pm18, Pm22, Pm28 and Pm29 were found in the hexaploid common wheat (Triticum aestivum). Pm19 was derived from Triticum tauschii Coss (2n = 14, DD); Pm4a and Pm5 from T. dicoccum (2n = 28,AABB); Pm4b from T. carthlicum (2n = 28, AABB); Pm6 and Pm27 from T. timopheevii (2n = 28, AAGG); Pm7, Pm8, Pm17 and Pm20 from Secale cereale (2n = 14, RR), Pm12 and Pm32 from T. speltoides (2n = 14, SbSb); Pm13 from T. longissimum (2n = 14, SpSp). Pm16 Pm26 and Pm31 from T. turgidum var. dicoccoides; Pm21 from Dasypyrum villosum (2n = 14, VV) and Pm25 from T. monococcum ssp. aegilopoides (2n = 14, AA). Pm10 and Pm11 were detected in T. spelta ssp. duhamelianum, Pm14 in Norin 10 and Pm15 in T. macha ssp. (McIntosh et al., 1998).
The most resistance genes/alleles are currently in use in Europe (Zeller et al., 1993; Peusha et al., 1996) however, in recent years most of the established genes for resistance have been ineffective (Bennett, 1984; Brown et al., 1990). Therefore additional effective resistance genes or alleles need to be identified to enable the continued diversification of gene combinations in breeding new cultivars. In the present study a new gene for resistance to mildew is described in the wild emmer T. dicoccoides accessions TA1055 and TA1150.
MATERIALS AND METHODS
Plant materials and isolate of Erysiphe graminis f. sp. tritici: Three T. durum varieties (IR4 = Tokhmi Siah; IR5 = Siah Daas and IR10 Chahar Tokhmi) from Iran. Three resistant accessions of T .dicoccoides TA64, TA1055 and TA1150 (Ahmadi firouzabad, 2001) were obtained from Kansas State University.
The Erysiphe graminis f. sp. tritici isolate Nor2 used for the inheritance and location of gene for resistance to powdery mildew was provided as described by Ahmadi firouzabad (2001). This isolate contains genes for virulence against Pm1, Pm2, Pm3a, Pm3c, Pm4a, Pm4b, Pm5, Pm9, Pm11, Pm15, Mld and Mlk .
Resistance analysis: The susceptible T. durum varieties were crossed with three resistant T. dicoccoides accessions in 2000 at department of field station, University of Newcastle Upon Tyne. The F1 hybrids were grown to produce F2 populations. Backcrosses to IR10 and crosses between TA1055 X TA1150 were made to confirm the number of genes for resistance present in these accessions. The methods of inoculation and conditions of incubation and disease assessment were used according to Hsam and Zeller (1997).
Chromosomal location: In this study the source of resistance T. dicoccoides is a tetraploid wheat, and thus 14 monosomic lines of Chinese Spring were used (1A-7A and 1B-7B) to cross as female with the resistant accessions TA1055 and TA1150. The F1 monopentaploid lines were grown to produce F2 populations. The F2 populations were inoculated with isolate Nor2. Twenty four hours prior to inoculation the isolate was tapped to dislodge the old conidia, and the two-week-old seedlings of parents and F2 plants were inoculated with the Nor2 isolate. After two weeks the numbers of plants with either resistant or susceptible reactions to mildew were recorded.
Cytological analysis: Meiosis in F1 hybrid plants was studied to identify monopentaploid hybrids. For the meiotic study, spikes of the F1 plants were fixed in acetic acid: ethanol (1:3) solution and anthers were selected for meiotic metaphase (MI) by carmine staining and stored in 70% alcohol at 4°C.
In a F1 monopentaploid, it is expected that 13 or 14 bivalents (genomes A and B) and 7 or 8 univalent (7 from genome D and one from genome A or B) are observed at metaphase I. In the case of F1 plants (2n = 34 or 2n = 35) at metaphase I, the number of univalents will be odd or even. The even ones are monopentaploid because 7 univalents belong to the D genome and one univalent is the homologous T. dicoccoides chromosome of the monosomic A or B genome chromosomes.
At early anaphase I the number of chromosomes were counted and plants with 2n = 34 and 2n = 35 chromosomes were identified.
RESULTS AND DISCUSSION
Inheritance of gene for resistance to powdery mildew: The F1 hybrids from all crosses were resistant when tested at the seedling stage and exhibited slight symptoms on their leaves, with scattered colonies beginning to necrose before much sporulation had occurred. By the second tiller stage, symptoms of infection had progressed to a characteristic chlorosis spreading from the tip of the oldest leaves (infection type 1-2). The leaf blades and spikes of all of the F1 plants were resistant at the heading stage and as at the seedling stage a few necrotic flecks were observed on leaf blades. However, the F1 plants displayed more obvious symptoms of susceptibility with moderate, sometimes heavy infection on leaf sheaths (IT 3-4), some sporulation and no apparent necrosis around colonies.
In the F2 generations, at the seedling stage, the F2 plants segregated in a ratio 1:2:1 with two groups resistant (IT 0-1) and (IT 2) and one susceptible (IT 3-4) (Table 1). This segregation can be explained in terms of one partially dominant gene present in the T. dicoccoides accessions.
At heading stage, the F2 plants were classified into 3 classes: ‘Class A’ in which the leaf blade (IT 0-1), leaf sheath (IT 0-1) and spikes were resistant; ‘Class B’ in which the leaf blades (IT 2) and spikes were resistant, but the leaf sheath was susceptible (IT 3); and ‘Class C’ in which the leaf blade (IT 3-4), leaf sheath (IT 3) and spikes were susceptible. The F2 segregations of crosses of IR10 with both T. dicoccoides accessions fit a 1 class A : 2 class B : 1 class C ratio (Table 2). In order to confirm the F2 segregation results, twenty F3 selfed seed was germinated from each F2 plant (5 to 7 F2 plants from each of the classes A, B and C) from the crosses IR10 x TA1055 and IR10 x TA1150. The results are summarized in Table 3. The offspring of class C F2 plants were all susceptible (IT 3-4) like their parents, which were obviously homozygous recessive for the gene for resistance to powdery mildew. The F2 resistant plants could be divided into segregating F3 families (class B) and families in which all individuals were fully resistant (class A). The latter class A F2 plants must have been homozygous dominant because all of the plants had a reaction IT 0-1 and the segregating F3 families (class B) obviously originated from F2 plants heterozygous for the gene for resistance since this group segregated plants with IT 0-1, IT 2 and IT 3-4. The ratio between F2 plants with segregating offspring (12 individuals) and with only resistant offspring (9 individuals) was expected to be 2:1. Despite the low numbers a chi-square value was calculated, indicating a good fit to the expectation (x2 = 0.86, P = 0.30-0.50). Thus it was concluded that resistance in T. dicoccoides accessions TA64, TA1055 and TA1150 is controlled by a single partially dominant gene.
The BC1 segregation of crosses of IR10 with both T. dicoccoides TA1055 and TA1150 fit a 1:1 ratio (Table 2).
| Table 1: | Segregation for resistance to Erysiphe graminis f. sp. tritici in three F2, reciprocal BC1 populations and their parents at the seedling stage |
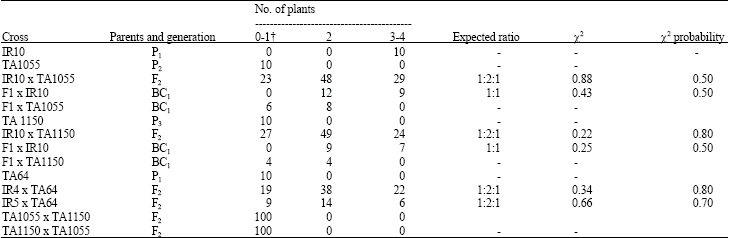 | |
| †Infection Type: 0-1 = Resistant, 2 = Moderately resistant, 3-4 = Susceptible | |
| Table 2: | Segregation for resistance to Erysiphe graminis f. sp. tritici in three F2, reciprocal BC1 populations and their parents at heading stage |
 | |
| Table 3: | Analysis of F3 families from F2 plants with resistant and susceptible reactions, from crosses between T. durum and two T. dicoccoides genotypes at the seedling stage |
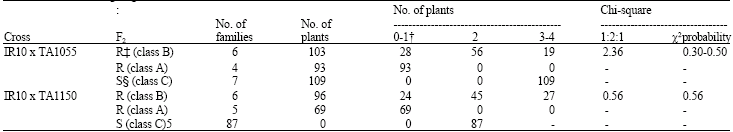 | |
| †Infection types: 0-1 = Resistant, 2 = Moderately resistant, 3-4 = Susceptible, ‡ = Resistant, § = Suseptible | |
This lead to confirm one dominant gene for resistance to powdery mildew is present in each T. dicoccoides accession. When the T. dicoccoides accessions were used as the recurrent parent, all of the BC1 progenies were resistant (Table 2). The F2 populations of reciprocal crosses between resistant TA1055 and TA1150 after inoculation with Nor2 isolate were all resistant and did not segregated (Table 2). This indicated that both T. dicoccoides accessions carry the same gene for resistance to powdery. The segregation results in F1, F2, F3 and reciprocal BC1 progenies confirmed that T. dicoccoides accessions TA1055 and TA1150 possess one similar dominant gene for resistance to Erysiphe graminis f.sp. tritici isolate Nor2.
Many sources of resistance to Erysiphe gramins f.sp. tritici have been identified in T. dicoccoides (Moseman et al., 1984). Thus T. dicoccoides can be a valuable genetic resource to broaden the genetic base of both cultivated species. Wild emmer population contain high frequencies of disease resistant genes. Gerechter-Amitai and Stubbs (1970) studied T. dicoccoides accessions collected from 32 sites, and found seedlings from 17 of the 55 accessions were resistant to Puccinia striiformis West. in the first leaf stage. Gerechter-Amitai and Grama (1974) showed that the resistance of selection G-25 of T. dicoccoides had one dominant gene conditioning its resistance to P. striiformis.
Moseman et al. (1984) tested the reaction of 233 T. dicoccoides accessions to infection with cultures of E. graminis f. sp. tritici. The reactions indicated that one hundred and fourteen or 49% of the accessions were resistant and 23 of the accessions were moderately resistant to infection with four cultures of E. graminis f. sp. tritici which possessed the virulence genes (Pm1, Pm2, Pm3a, Pm3c, Pm4).
Self seed-set: The emergence rate of F1 monopentaploid hybrid seed was 97.5 and 98.2% for crosses CS/TA1055 and CS/TA1150, respectively. This indicates that the majority of the monopentaploid seeds were viable. No selfed seed was produced by the F1 hybrid plants from crosses CS/TA1150. The F1 plantsof monopentaploid families had a range of 1.9 to 9.2 mean percent seed-set per plant.
| Table 4: | Segregation for seedling reaction to mildew isolate Nor2 in 14 monopentaploid F2 populations from crosses of 14 Chinese Spring monosomics with T. diccoccoides accession TA1055 |
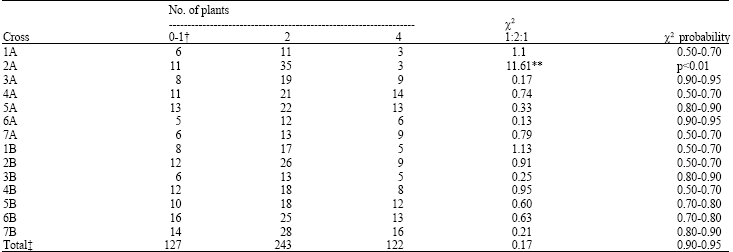 | |
| † Infection Types: 0-1 = no symptoms or very slight chlorotic flecking on the leaves; IT 2 = small pustules surrounded by necrosis or chlorosis, no sporulation; IT 4 = large pustules with massive sporulation, no chlorossis ‡ Excluding mono-2A, **p<0.01 | |
Eight monopentaploid F1 hybrid lines, 1A, 3A, 4A, 6A, 7A, 3B, 4B and 5B, showed a significant reduction in self seed per plant, indicating that at least eight chromosomes may have factors which influence self seed-set.
Chromosomal location: The F2 progeny from crosses of the 14 Chinese Spring (CS) monosomic lines with TA1055 were inoculated with the Nor.2 isolate. Thirteen of the F2 progeny representing each hybrid combination and the pooled data (excluding mono-2A hybrid) segregated in a ratio of 3:1 (Table 4) for the three infection types. The more excess of type 2 of F2 plants in mono-2A population can be explained as follows. When a monosomic plant is allowed to produce selfed-seed, it is expected that the majority of the F2 plants to be monosomic, because the frequency of monosomic plants in a F2 population has been predicted about 73%. In this study the frequency of plants with type 2 which can be monosomic for chromosome 2A was 72.9% which is most closely to the expected frequency. This indicated that the gene for resistance to wheat powdery mildew is present and expressed in monopentaploid hybrids. The F2 segregation from the monopentaploid hybrid CS mono-2A x TA1055 deviated significantly (p<0.01) from the expected 1:2:1 ratio. The results revealed that the gene conferring resistance to powdery mildew is located on chromosome 2A.
The reaction of T. dicoccoides accessions was different from the wheat mildew resistance genes Pm4a and Pm4b which are known to be located on chromosome 2A (McIntosh et al., 1998). Because Nor 2 was virulent to Pm4b, Pm4a and avirulent on the T. dicoccoides accessions. Resistance to powdery mildew in Td1055 and Td1150 is controlled by a gene other than Pm16 and Pm26 already transferred to common wheat from T. dicoccoides. The gene Pm16 is located on chromosome 4A (Reader and Miller, 1991) and the gene Pm26 is located on chromosome 2B (Rong et al., 2000) but the gene for resistance present in Td1055 is located on chromosome 2A. It is proposed that this new resistance gene in TA1055 which is different from 33 current known genes for resistance to wheat powdery mildew, should be temporarily designated Mltd1055.
REFERENCES
- Bennett, F.G.A., 1984. Resistance to powdery mildew in wheat: A review of its use in agriculture and breeding programmes. Plant Pathol., 33: 279-300.
CrossRefDirect Link - Gerechter-Amitai, Z.K. and R.W. Stubbs, 1970. A valuable source of yellow rust resistance in Israel populations of wild emmer, Triticum dicoccoides Koern. Euphytica, 19: 12-21.
CrossRefDirect Link - Gerechter-Amital, Z.K. and A. Grama, 1974. Inheritance of resistance to stripe rust (Puccinia striformis) in crosses between wild emmer (Triticum dicoccoides) and cultivated tetraploid and hexaploid wheats. I. Triticum durum. Euphytica, 23: 387-392.
CrossRefDirect Link - Hsam, S.L.K. and F.J. Zeller, 1997. Evidence of allelism between genes Pm8 and Pm17 and chromosomal location of powdery mildew and leaf rust resistance genes in the common wheat cultivar Amigo. Plant Breed., 116: 119-122.
Direct Link - Hsam, S.L.K., I.F. Lapochkina and F.J. Zeller, 2003. Chromosomal location of genes for powdery mildew resistance in common wheat (Triticum aestivum L. em Thell.). 8. Gene Pm32 in a wheat-Aegilops speltoides translocation line. Euphytica, 133: 367-370.
Direct Link - Jarve, K., H.O. Peusha, J. Tsymbalova, S. Tamm, K. Devos and T. Enno, 2000. Chromosomal location of a Triticum timopheevii derived powdery mildew resistance gene transferred to common wheat. Genome, 43: 377-381.
Direct Link - Liu, Z.Y., Q.X. Sun, Z.F. Ni, E. Nevo and T.M. Yang, 2002. Molecular characterization of a novel powdery mildew resistance gene Pm30 in wheat originating from wild emmer. Euphytica, 123: 21-29.
CrossRefDirect Link - Moseman, J.G., E. Nevo, M.A. Elmorshidy and D. Zohary, 1984. Resistance of Triticum dicoccoides to infection with Erysiphe graminis f. sp. tritici. Euphytica, 33: 41-47.
CrossRef - Peusha, H., S.L.K. Hsam and F.J. Zeller, 1996. Chromosomal location of powdery mildew resistance genes in common wheat (Triticum aestivum L. em. Thell.) 3. Gene Pm22 in cultivar Virest. Euphytica, 91: 149-152.
Direct Link - Peusha, H., T. Enno and O. Priilinn, 2000. Chromosomal location of powdery mildew resistance genes and cytogenetic analysis of meiosis in common wheat cultivar Meri. Hereditas, 132: 29-34.
Direct Link - Reader, S.M. and T.E. Miller, 1991. The introduction into bread wheat of a major gene for resistance to powdery mildew from wild emmer wheat. Euphytica, 53: 57-60.
Direct Link - Rong, J.K., E. Millet, J. Manisterski and M. Feldman, 2000. A new powdery mildew resistance gene: Introgression from wild emmer into common wheat and RFLP-based mapping. Euphytica, 115: 121-126.
Direct Link - Shi, A.N., S. Leath and J.P. Murphy, 1998. A major gene for powdery mildew resistance transferred to common wheat from wild einkorn wheat. Phytopathology, 88: 144-147.
Direct Link - Xie, C., Q. Sun, Z. Ni, T. Yang, E. Nevo and T. Fahima, 2003. Chromosomal location of a Triticum dicoccoides-derived powdery mildew resistance gene in common wheat by using microsatellite markers. Theor. Appl. Genet., 106: 341-345.
CrossRefDirect Link - Zeller, F.J., L. Kong, L. Hartl, V. Mohler and S.L.K. Hsam, 2002. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.) 7. Gene Pm29 in line Pova. Euphytica, 123: 187-194.
Direct Link