
Fakher Ayed
Horticultural High School of Chott-Mariem, 4042 Sousse, Tunisia
Mejda Daami-Remadi
National Institute of Agronomic Research, Tunisia
(PRRDA-CE Chott-Mariem, 4042 Sousse- Tunisia)
Hayfa Jabnoun-Khiareddine
Horticultural High School of Chott-Mariem, 4042 Sousse, Tunisia
Khaled Hibar
Horticultural High School of Chott-Mariem, 4042 Sousse, Tunisia
Mohamed El Mahjoub
Horticultural High School of Chott-Mariem, 4042 Sousse, Tunisia
Plant Pathology Journal
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 239-243







ABSTRACT
Four fungicides, which active ingredients are hymexazol, azoxystrobin, fludioxonil and quinoline, are tested in vitro and in vivo against five isolates of Fusarium oxysporum f. sp. tuberosi, causal agent of potato Fusarium wilt. Pathogen radial growth noted on PDA amended with fungicides after incubation for four days at 25°C varied upon pathogen isolates. Mycelial growth of all pathogen isolates was significantly inhibited by fungicide PDA amendment; inhibition obtained by hymexazol treatment reached 77% comparatively to untreated control. All chemical fungicides reduced disease incidence compared to the untreated control. Hymexazol and azoxystrobin are the most active during all the bioassay period. Whereas, fludioxonil and quinoline showed a limited effect in controlling Fusarium wilt development.
PDF Abstract XML References Citation
How to cite this article
Fakher Ayed, Mejda Daami-Remadi, Hayfa Jabnoun-Khiareddine, Khaled Hibar and Mohamed El Mahjoub, 2006. Evaluation of Fungicides for Control of Fusarium Wilt of Potato. Plant Pathology Journal, 5: 239-243.
DOI: 10.3923/ppj.2006.239.243
URL: https://scialert.net/abstract/?doi=ppj.2006.239.243
DOI: 10.3923/ppj.2006.239.243
URL: https://scialert.net/abstract/?doi=ppj.2006.239.243
INTRODUCTION
Fusarium oxysporum (Schlecht.) Snyd. et Hans. is frequent in warm areas. It causes vascular wilt in several plants (Theron, 1991; Meulemans, 1996; Norguès et al., 2002). In Tunisia, Fusarium oxysporum f. sp. tuberosi causes dry rot of tubers and vascular wilts of potatoe plants (Daami-Remadi and El Mahjoub, 2004).
This soilborne fungus infected plants through roots and colonized xylem vessels of stems. Potato plants infected by Fusarium oxysporum f. sp. tuberosi showed a characteristic discoloration in their vascular tissue due to obstruction of the water and nutrient-conducting tissue of the plant resulting in stunting, chlorosis, unilateral wilt, and eventual plant death (Hwang and Evans, 1985; Kucharek et al., 2000). This pathogen caused economic losses estimated of 10 to 53% of potato yield (Thanassoulopoulos and Kitsos, 1985).
As it is a soil-borne pathogen, control of F. oxysporum f. sp. tuberosi has generally been restricted to the use of long-term rotations (3-5 years) and solarization. However, in Tunisia these techniques are not generalised due to limited surfaces and high cost of solarization (Katan, 1980; Monnet, 2001; Triki et al., 2001). While in several other countries, fumigation with methyl bromide, which was effective in reducing soilborne inoculum of numerous Fusarium, will be totally removed from the agricultural markets, because of its ozone-depleting effect (Watson et al., 1992). As alternatives to these control measures, several reports demonstrated successful use of biological control agents (mostly bacteria and fungi) and fungicides for control of this disease (Larkin et al., 1998; Reid et al., 2002; Elmer and McGovern, 2004). In fact, some fungicides are reported to be rated with reduced risk toxicology (Errampalli, 2004).
Since Fusarium wilt is a serious threat a strategic crop in Tunisia, control measures integrating with cropping practices are searched. The present research was conducted in order to evaluate the effectiveness, in vitro and in vivo, of four fungicides in controlling F. oxysporum f. sp. tuberosi.
MATERIALS AND METHODS
Fungal cultures and fungicides: Isolates of Fusarium oxysporum f. sp. tuberosi used in this study (Fot1, Fot2, Fot3, Fot4 and Fot5) were obtained from potato tubers showing dry rot symptoms collected from traditional potato-stores (Hammam Ghezaz, Hawaria and Korba).
| Table 1: | Fungicides tested against F. oxysporum f. sp. tuberosi development |
 | |
| Table 2: | Fungicide relative rates used in in vitro experiments against F. oxysporum f. sp. tuberosi (Fot) |
 | |
They were cultured on Potato Dextrose Agar (PDA) and incubated at 25°C in the dark. Single spore cultures were maintained on glycerol at -80°C for long-term storage.
For plant inoculation, mycelium taken from the edge colony of each isolate was transferred to 150 mL of Potato Dextrose Broth (PDL) and incubated at 25°C for 5 days in a rotary incubator (120 rpm). The liquid culture was filtered and the conidial suspension was adjusted to 107 spores.mL-1 by means of a Malassez cystometer.
The characteristics of the four fungicides tested were listed in Table 1.
In vitro experiments: Five isolates of F. oxysporum f. sp. tuberosi was grown on Potato Dextrose Agar (PDA) at 25°C for 7 days. Agar disks (6 mm) bearing the fungus were taken from freshly growing colony and transferred on PDA amended with various concentrations of fungicides (Table 2). The fungal radial growth of colonies was recorded after incubation for four days at 25°C. Eight replicates were used per elementary treatment.
Data are arranged as a complete randomized factorial design where treatments and isolates are both fixed factors. They were analyzed using SPSS and subjected to analysis of variance and Fisher’s least significant difference test LSD (at p<5%).
In vivo experiments: An isolate of F. oxysporum f. sp. tuberosi (Fot3), being the most aggressive following pathogenicity tests (Ayed et al., 2006), was used for plant inoculation. Potato tubers, cv. Spunta, were planted in plastic pots (6.74 l) containing an autoclaved mixture of perlite and peat (1:3) and kept at 8-32°C (minimum and maximum temperatures, respectively). Two weeks after their emergence, plants were inoculated by irrigation with 100 mL-1 of conidial suspension (107 spores mL-1). Ten non-inoculated control plants were treated similarly with 100 mL-1 of sterile distilled water. Ten days after inoculation, fungicides are applied at their relative rates (Table 3) using a drip spray method.
| Table 3: | Fungicide rates used in in vivo experiments for suppression of potato Fusarium wilt |
 | |
A second fungicide treatment was realized 20 days after. Potato plants were irrigated regularly and fertilized with a nutrient solution (20 N:20 K2O:20 P2O5) following Manici and Cerato (1994) method.
Effect of the tested fungicides on Fusarium wilt development was assessed via disease severity index. A scale of 0-4 was used to assess disease severity: 0 = asymptomatic leaf, 1 = leaf wilted, 2 = Leaf with hemiplegic yellowing, 3 = leaf with necrosis, 4 = dead leaf. Incidence of F. oxysporum f. sp. tuberosi were estimated weekly via an Index of Leaf Damage (ILD) calculated per potato plant following formula (Béye and Lafay, 1985) where:
| ILD | : | Index of Leaf Damage. |
| Σ notes | : | Total notes. |
| Max | : | 4 times of developed-leaves number. |
Data are arranged by completely randomized design where treatments (plants treated by each of five fungicides, inoculated and non-inoculated) are the only fixed factor. Ten plants per treatment were assessed. Mean comparisons were done following the LSD method (p<5%).
RESULTS
In vitro experiments: Colony diameters of F. oxysporum f. sp. tuberosi on PDA, amended with relative fungicide concentrations, noted after four days of incubation at 25°C, varied among fungicides and pathogen isolates. An interaction was observed between both fixed factors (Table 4). All fungicides reduced significantly mycelial growth of F. oxysporum f. sp. tuberosi isolates (Fig. 1 and Table 4).
Hymexazol showed the highest efficiency in reducing mycelial growth for more than 77%, compared to pathogen growth on unamended PDA. At 2 ppm, fludioxonil significantly limited mycelial development by 73% for Fot3. Azoxystrobin also showed fungicidal activity against F. oxysporum f. sp. tuberosi isolates. Quinoline showed lesser efficiency; mycelial growth was reduced by 30 to 43% for all tested isolates.
| Table 4: | Mean diameters of F. oxysporum f. sp. tuberosi colonies noted in presence of several fungicide treatments comparatively with the untreated control (PDA, after six days of incubation at 25°C) |
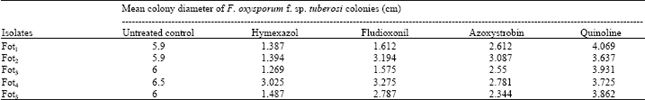 | |
| Fot1, Fot2, Fot3, Fot4 and Fot5: Isolates of F. oxysporum f. sp. tuberosi LSD at p<5% (Treatments x Isolates) = 0.273 cm | |
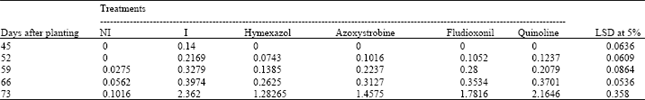
| Table 5: | Weekly evolution of mean Index of Leaf Damage (ILD) of potato plants, cv. Spunta, inoculated by F. oxysporum f. sp. tuberosi and treated by four fungicides comparatively with healthy (NI) and untreated-inoculated (I) plants |
 | |
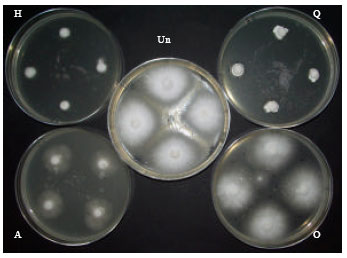 | |
| Fig. 1: | Mycelial growth of a F. oxysporum f. sp. tuberosi isolate (Fot3) observed in presence of several treatments (PDA, after 4 days of incubation at 25°C). (Un: Untreated; H: Hymexazol; A: Azoxystrobin; F: Fludioxonil; Q: Quinoline) |
 | |
| Fig. 2: | Comparison between Non-inoculated (NI) and inoculated (I+U) potato plants and inoculated plants treated 10 and 30 days after inoculation by four fungicides (H: Hymexazol; A: Azoxystrobin; F: Fludioxonil; Q: Quinoline). (cv. Spunta, 73 days after inoculation) |
Fot4 had the least important inhibition, whereas Fot3 was the most sensitive isolate to most tested fungicides.
In vivo experiments: Weekly evolution of Leaf Damage Index (ILD) of potato plants, cv. Spunta, inoculated by a virulent F. oxysporum f. sp. tuberosi isolate and treated by several fungicides was assessed as soon as the first typical Fusarium wilt symptom appeared (30 days after inoculation).
All tested fungicides tested significantly reduced the Fusarium wilt incidence compared to untreated-inoculated plants during all this bioassay (Table 5 and Fig. 2).
The ILD of potato plants treated with hymexazol didn’t exceed 1.28 at 73 days after planting in comparison to 2.36 for untreated-inoculated plants. Moreover, Azoxystrobin showed a remarkable efficiency in reducing severity of this disease. Fludioxonil reduced significantly Fusarium wilt but its efficiency was lesser than both previous fungicides. Quinoline was effective during the first three weeks after first symptom observation where the ILD increased from 0.37 to 2.16 at the end of the bioassay (Table 5). Furthermore, disease incidence increased suddenly in the last week of the bioassay especially for treated and untreated-inoculated plants.
DISCUSSION
Vascular infections by F. oxysporum f. sp. tuberosi and pathogen soilborne origin make disease control difficult. Chemical compounds tested in the present study (hymexazol, azoxystrobin, fludioxonil and quinoline) showed in vitro fungicidal effects on PDA of five isolates of F. oxysporum f. sp. tuberosi. Moreover, the in vivo experiment revealed their efficacy in controlling Fusarium wilt development. Hymexazol showed, in vitro as in vivo, the highest activity against this pathogen. Similar results were obtained with the same dosage and application procedures in reducing Fusarium wilt of tomato caused by F. oxysporum f. sp. radicis-lycopersici (Hibar, 2002). Daami-Remadi (2001) signalled its effectiveness in inhibiting by 43 to 50% potato leak syndrome caused by Pythium aphanidermatum. Moreover, azoxystrobin provided efficacy in the control of potato Fusarium wilt, as reported in carnation-F. oxysporum f. sp. dianthi pathosystem (Gullino et al., 2000). However, Elmer and McGoven (2004) reported that this strobilurin has poor curative properties in reducing Fusarium wilt of Cyclamen caused by F. oxysporum f. sp. cyclaminis and suggested their preventive application for this disease suppression. Bertelsen et al. (2001) explained fungicidal action of azoxystrobin by blocking electron transport in the fungal mitochondrial respiratory chain. Fludioxonil and quinoleine showed limited efficiency against F. oxysporum f. sp. tuberosi in in vivo experiments, in spite of their good activity against Botrytis cinerea, Monilinia spp. and Sclerotinia spp. reported by Gullino et al. (2000). In the same way, Reid et al. (2002) found that fludioxonil limited plant death caused by F. oxysporum f. sp. asparagi and F. proliferatum at high inoculum level. However, Elmer and McGovern (2004) confirmed that fludioxonil has poor curative properties against Fusarium wilt of Cyclamen. Therefore, an increase in dose and/or soil incorporation should be tested especially where Errampalli (2004) signalled that any chemicals that significantly inhibit germination and mycelial growth should reduce the ability of pathogen to cause disease. Furthermore, Bains et al. (2001) found that mixture of fludioxonil with other compounds such as mancozeb or difenoconazole provided tuber protection from seed piece decay caused by F. sambucinum. The number, the frequency and timing of applications also play an important role in efficacy of fungicides in different pathosystems (Bubici et al., 2005).
Suddenly increase of disease incidence at the end of the bioassay can be caused by obstruction of the water and nutrient-conducting tissue of inoculated plants as a result of inoculum level increase as by loss of activity of fungicides (Kucharek et al., 2000).
The present study showed that all fungicides limited F. oxysporum f. sp. tuberosi in vitro, but hymexazol and azoxystrobin are the most efficient in the in vivo experiment probably due to rates used in plant treatments. Integrating chemicals with biological agents proven effective and consisted alternative in several pathosystems and it has not been completely explored for potato Fusarium wilt.
ACKNOWLEDGMENTS
Authors thank High School of Horticulture and Breeding of Chott-Mariem, Technical Potato Centre of Tunisia and Interprofessional Groupment of Legumes for their financial contribution. Many thanks for Aymen Youssef for the excellent technical assistance.
REFERENCES
- Ayed, F., M. Daami-Remadi, H. Jabnoun-Khiareddine and M. El Mahjoub, 2006. Potato vascular fusarium wilt in Tunisia: Incidence and biocontrol by Trichoderma sp. Plant Pathol. J., 5: 92-98.
CrossRef - Elmer, W.H. and R.J. McGovern, 2004. Efficacy of integrating biologicals with fungicides for the suppression of Fusarium wilt of Cyclamen. Crop Prot., 23: 909-914.
Direct Link - Errampalli, D., 2004. Effect of fludioxonil on germination and growth of Penicillium expansum and decay in apple cvs. Empire and Gala. Crop Prot., 23: 811-817.
CrossRefDirect Link - Gullino, M.L., P. Leroux and C.M. Smith, 2000. Uses and challenges of novel compounds for plant disease control. Crop Prot., 19: 1-11.
CrossRefDirect Link - Larkin, R.P. and D.R. Fravel, 1998. Efficacy of various fungal and bacterial biocontrol organisms for control of Fusarium wilt of tomato. Plant Dis., 82: 1022-1028.
CrossRefDirect Link - Manici, L.M. and C. Cerato, 1994. Pathogenicity of Fusarium oxysporum f. sp. tuberosi isolates from tubers and potato plants. Potato Res., 37: 129-134.
Direct Link - Reid, T.C., M.K. Hausbeck and K. Kizilkaya, 2002. Use of fungicides and biological controls in the suppression of Fusarium crown and root rot of asparagus under greenhouse and growth chamber conditions. Plant Dis., 86: 493-498.
Direct Link - Theron, D.J. and G. Holz, 1991. Prediction of potato dry rot based on the presence of Fusarium in soil adhering to tubers at harvest. Plant Dis., 75: 126-130.
Direct Link - Triki, M.A., S. Priou and M. El Mahjoub, 2001. Effects of soil solarization on soil-borne populations of Pythium aphanidermatum and Fusarium solani and on the potato crop in Tunisia. Potato Res., 44: 271-279.
Direct Link - Bubici, G., M. Amenduni, C. Colella, M.D'Amico and M. Cirulli, 2006. Efficacy of acibenzolar-S-methyl and two strobilurins, azoxystrobin and trifloxystrobin, for the control of corky root of tomato and verticillium wilt of eggplants. Crop Prot., 25: 814-820.
CrossRefDirect Link