
Igboh M. Ngozi
Department of Biochemistry, Faculty of Science, Abia State University, P.M.B. 2000, Uturu, Nigeria
Ikewuchi C. Jude
Department of Biochemistry, Faculty of Science, University of Port Harcourt, P.M.B. 5323, Port Harcourt, Nigeria
C. Catherine
Department of Biochemistry, Faculty of Science, University of Port Harcourt, P.M.B. 5323, Port Harcourt, Nigeria
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 5 | Page No.: 521-524







ABSTRACT
The proximate, amino acid and phytochemical composition of Chromolaena odorata was investigated. A high total carbohydrate (20.58% WW and 50.82% DW), crude fibre (10.76% WW and 26.57% DW) and protein (6.56% WW and 16.20% DW) content was observed. The protein is rich in the essential amino acids (with histidine and phenylalanine being very high) and has a protein score of 88.24% with methionine as the limiting amino acid. The phytochemical screening revealed the presence of alkaloids, cyanogenic glycosides, flavonoids (aurone, chalcone, flavone and flavonol), phytates saponins and tannins. The anti-nutrients composition includes cyanogenic glycosides (0.05% WW and 0.13% DW), phytates (0.22% WW and 0.54% DW), saponins (0.80% WW and 1.98% DW) and tannins (0.15% WW and 0.37% DW). Our result suggests that C. odorata is a source of high quality protein which could serve as a potential source of protein supplement.
PDF Abstract XML References Citation
How to cite this article
Igboh M. Ngozi, Ikewuchi C. Jude and C. Catherine, 2009. Chemical Profile of Chromolaena odorata L. (King and Robinson) Leaves. Pakistan Journal of Nutrition, 8: 521-524.
DOI: 10.3923/pjn.2009.521.524
URL: https://scialert.net/abstract/?doi=pjn.2009.521.524
DOI: 10.3923/pjn.2009.521.524
URL: https://scialert.net/abstract/?doi=pjn.2009.521.524
INTRODUCTION
Chromolaena odorata (L) King and Robinson (also Osmia odorata L. (Schultz-Bip), or Euphatorium odoratum L.), belongs to the family Asteraceae. Its common names include “Awolowo”, “Independence weed”, siam weed, triffid weed, bitter bush or jack in the bush (Okon and Amalu, 2003). Although, native to South and Central America it is has spread throughout the tropics, Nigeria inclusive (http://www.ehs.cdu.edu.au/ chromolaena/pubs/friend.html). In traditional medicine, it is used as an antispasmodic, antiprotozoal, antitrypa-nosomal, antibacterial, antifungal, antihypertensive, anti-inflammatory, astringent, diuretic and hepatotropic agent (Iwu, 1993; Phan et al., 2001; Akinmoladun et al., 2007). The present study is designed to assess the proximate, phytochemical and amino acid composition of C. odorata.
MATERIALS AND METHODS
Samples of fresh Chromolaena odorata plants were collected from within the Abuja Campus of University of Port Harcourt, Port Harcourt, Nigeria. After due identification at the University of Port Harcourt Herbarium, Port Harcourt, Nigeria, they were rid of dirt, before collecting and storing their leaves, for subsequent use. A portion was immediately subjected to proximate analysis. The moisture, crude protein, fat, ash, fiber and total carbohydrate contents were determined in triplicates, according to standard methods (AOAC, 2006). The energy value was calculated using the Atwater factors of 4, 9 and 4 for protein, fat and carbohydrate, respectively (FAO/WHO/UNU, 1991; Chaney, 2006). The phytochemical screening of the sample was carried out as described by Sofowora (1980) and Harbone (1973). They were screened for alkaloids, cyanogenic glycosides, flavonoids (aurone, chalcone, flavone and flavonol), phytates, saponins and tannins. Quantitative determination of phytates, saponins and tannins were carried out in triplicates, using the method of AOAC (2006).
The amino acid profile of the sample was determined using methods described by Spackman et al. (1985). The sample was dried to a constant weight, defatted, hydrolyzed, evaporated in a rotary evaporator and loaded into a Technicon Sequential Multi-sample Amino Acid Analyzer (TSM).
RESULTS AND DISCUSSION
The proximate composition of C. odorata is given in Table 1. Its moisture content is less than those of Amarantus hybridus, Telferia occidentalis, Talinum triangulare (Oguntona, 1998) and Pennisetum purpureum (Okaraonye and Ikewuchi, 2009). The moisture content of any food is an index of its water activity (Olutiola et al., 1991) and is used as a measure of stability and the susceptibility to microbial contamination (Uraih and Izuagbe, 1990). This implies that:
| • | C. odorata may have a short shelf-life due to its high moisture content. |
| • | Dehydration would increase the relative concentrations of the other food nutrients and improve the shelf-life/preservation of the C. odorata |
| Table 1: | Proximate composition of Chromolaena odorata |
 | |
| Values are means of triplicate determinations | |
| Table 2: | Amino acid profile of protein from Chromolaena odorata |
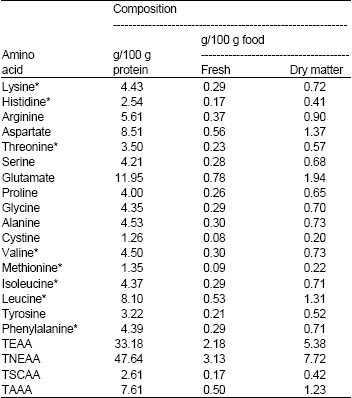 | |
| *Essential amino acids. TEAA = Total Essential Amino Acid. TNAA = Total Nonessential Amino Acid. TSCAA = Total Sulphur-Containing Amino Acid. TAAA = Total Aromatic Amino Acid | |
We observed a higher protein content than those reported for A. hybridus, T. occidentalis, T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009); although this is lower than the value earlier reported for the same plant by Apori et al. (2000). The total fat content of C. odorata is less than those found in A. hybridus, T. occidentalis, T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009). Its total carbohydrate content is greater than those of A. hybridus, T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009). Crude fibre content recorded in this study is greater than those reported for A. hybridus, T. occidentalis, T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009). Some epidemiological evidences suggest that increased fibre consumption may contribute to a reduction in the incidence of certain diseases including colon cancer, coronary heart disease, diabetes, high blood pressure, obesity and various digestive disorders (Walker, 1978; FAO, 1990; Eriyamremu and Adamson, 1994; SACN, 2008). They increase fecal bulk and rate of intestinal transit and have prebiotic effects. We observed a lower ash content than was reported for A. hybridus and T. occidentalis, although greater than those reported for T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009). The total metabolizable energy in C. odorata is greater than those of A. hybridus, T. triangulare (Oguntona, 1998) and P. purpureum (Okaraonye and Ikewuchi, 2009).
The amino acid profile and chemical scores of C. odorata leaf protein are given in Table 2 and 3, respectively. It is rich in the essential amino acids (especially histidine and phenylalanine) and can meet the minimum daily requirements (FAO/WHO/UNU, 1991; McGilvery and Goldstein, 1983) for the essential amino acids, except methionine, valine and lysine. Relative to the WHO reference protein pattern (FAO/WHO/UNU, 1991; McGilvery and Goldstein, 1983), its limiting amino acid is methionine and its protein score is 88.24%. This score is higher than the reported values for soy bean, maize, peanuts, polished rice and whole wheat and lower than those of human milk, egg, cow milk and beef (McGilvery and Goldstein, 1983; FAO, 1981). Every 100 g of this protein contains 33.18 g of essential amino acids, 2.61 g of sulphur-containing amino acids and 7.61 g of aromatic amino acids (Table 2). So, in the light of the earlier report of its high degradability of about 953 g/kg DM by Apori et al. (2000), this protein can be an invaluable means of protein supplementation.
The phytochemical screening revealed that C. odorata is highly rich in saponins, moderately rich in phytates and tannins, with little content of alkaloids, flavonoids (aurone, chalcone, flavone and flavonol) and cyanogenic glycosides (Table 4). The antinutrient composition of C. odorata is shown in Table 5. The phytate content of C. odorata is greater than those of T. triangulare, Piper guineense, T. occidentalis (Oguntona, 1998), Vigna unguiculata (Osagie, 1998), Trichosanthes anguina fruits (Ojiako and Igwe, 2008) and P. purpureum (Okaraonye and Ikewuchi, 2009). Phytic acid binds calcium, zinc, iron and other minerals, thereby reducing their availability in the body (FAO, 1990). It also inhibits protein digestion by forming complexes with them (Singh and Krikorian, 1982). However, the phytate content of foods can be reduced by a number of processing methods (FAO, 1990). Very high saponin content was observed in this study. It is higher than that reported for Vernonia amygdalina (Ejoh et al., 2007), but less than what we reported for P. purpureum (Okaraonye and Ikewuchi, 2009). Saponins reduce the uptake of certain nutrients including glucose and cholesterol at the gut through intra-lumenal physicochemical interaction and have been reported to have hypocholesterolemic effects (Price et al., 1987).
| Table 3: | Comparison of protein from Chromolaena odorata, with WHO reference protein pattern (McGilvery and Goldstein, 1983; FAO/WHO/UNU, 1991) |
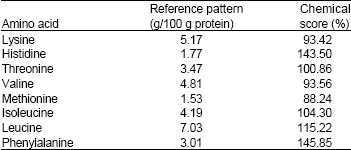 | |
| Table 4: | Phytochemical profile of Chromolaena odorata |
 | |
| Key: + = slightly present; ++ = moderately present; +++ = highly present | |
| Table 5: | Anti-nutrient composition of Chromolaena odorata |
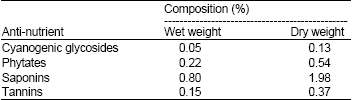 | |
| Values are Means ± SD of triplicate determinations | |
Thus when consumed, they may aid in lessening the metabolic burden that would have been placed on the liver. Moderately high tannin content was recorded in this study. It is higher than that reported for T. anguina fruits (Ojiako and Igwe, 2008) and what was earlier reported for C. odorata by Apori et al. (2000). This high tannin content can probably be lowered by processing methods such as soaking, boiling and fermentation (Esenwah and Ikenebomeh, 2008).
Finally, our results suggest that C. odorata is rich in high quality protein. The implication of this in nutritional protein supplementation can not be overemphasized.
REFERENCES
- Akinmoladun, A.C., E.O. Ibukun and I.A. Dan-Ologe, 2007. Phytochemical constituents and antioxidant properties of extracts from the leaves of Chromolaena odorata. Sci. Res. Essays, 2: 191-194.
Direct Link - Ejoh, R.A., D.V. Nkonga, G. Inocent and M.C. Moses, 2007. Nutritional components of some non-conventional leafy vegetables consumed in Cameroon. Pak. J. Nutr., 6: 712-717.
CrossRefDirect Link - Eriyamremu, G.E. and I. Adamson, 1994. Early changes in energy metabolism in rats exposed to an acute level of deoxycholate and fed on a Nigerian-like diet. Ann. Nutr. Metab., 38: 174-183.
Direct Link - Esenwah, C.N. and M.J. Ikenebomeh, 2008. Processing effects on the nuttritional and anti-nutritional contents of African locust bean (Parkia biglobosa Benth) seed. Pak. J. Nutr., 7: 214-217.
CrossRefDirect Link - Harbone, J.B., 1973. Phytochemical Methods: A Guide to Modern Technique of Plant Analysis. 2nd Edn., Chapman Hall, New York, Pages: 278.
Direct Link - Ojiako, O.A. and C.U. Igwe, 2008. The nutritive and hepatotoxic properties of Trichosanthes anguina (snake tomato) fruits from Nigeria. Pak. J. Nutr., 7: 85-89.
CrossRefDirect Link - Okaraonye, C.C. and J.C. Ikewuchi, 2009. Nutritional and antinutritional components of Pennisetum purpureum (Schumach). Pak. J. Nutr., 8: 32-34.
CrossRefDirect Link - Siddhuraju, P., V. Vijayakumari and K. Janardhanan, 1995. Studies on the unexploited legumes, Indigofera linifolia and Sesbania bispinosa Nutrient composition and antinutritional factors. Int. J. Food Sci. Nutr., 46: 195-203.
CrossRefDirect Link - Phan, T.T., L. Wang, P. See, R.J. Grayer, S.Y. Chan and S.T. Lee, 2001. Phenolic compounds of Chromolaena odorata protect cultured skin cells from oxidative damage: Implication for cutaneous wound healing. Biol. Pharm. Bull., 24: 1373-1379.
CrossRefDirect Link - Price, K.R., I.T. Johnson, G.R. Fenwick and M.R. Malinow, 1987. The chemistry and biological significance of saponins in foods and feedingstuffs. Crit. Rev. Food Sci. Nutr., 26: 27-135.
CrossRefPubMedDirect Link - Singh, M. and A.D. Krikorian, 1982. Inhibition of trypsin activity in vitro by phytate. J. Agric. Food Chem., 30: 799-800.
CrossRefDirect Link - Spackman, D.H., W.H. Stein and S. Moore, 1958. Automatic recording apparatus for use in chromatography of amino acids. Anal. Chem., 30: 1190-1206.
CrossRefDirect Link