
H. Silini-Cherif
Department of Microbiology, Laboratory of Applied Microbiology, Faculty of Natural and Life Sciences, University of Ferhat-Abbas, Setif, Algeria
A. Silini
Department of Microbiology, Laboratory of Applied Microbiology, Faculty of Natural and Life Sciences, University of Ferhat-Abbas, Setif, Algeria
M. Ghoul
Department of Microbiology, Laboratory of Applied Microbiology, Faculty of Natural and Life Sciences, University of Ferhat-Abbas, Setif, Algeria
S. Yadav
Research Scholar School of Life Sciences, Hyderabad Central University, Hyderabad, India
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 6 | Page No.: 267-276







ABSTRACT
The use of microbial technology in agriculture is expanding quickly with the identification of new bacterial strains which are more effective in promoting the growth of plants. The rhizobacteria that promote the growth of plants can have a positive effect on the productivity of crops especially when subjected to salt stress. A nitrogen-fixing bacterium was isolated from the wheat rhizosphere of an arid region. The strain was identified on the basis of tests API20E and 16S rRNA sequencing, as Pantoea agglomerans lma2. This strain degraded several carbon sources: sugars (fructose, ribose, dextrin, salicin…), lipids (lecithin, tributyrin and tween 80), proteins (gelatin, casein), grew on KCN and could grow from pH 4 to 8 and had an optimum at pH 7. The growth temperature showed a maximum at 30°C and the bacteria could tolerate from 4 to 41°C and the growth rate was higher when the NaCl concentration was between 100 and 300 mM. The performance of activities enhancing the growth of plants of P. agglomerans lma2 was significantly better in the presence of salt. Rates of Indole Acetic Acid (IAA), siderophores production and solubilization of phosphate increased between 100 and 400 mM NaCl compared to the control without salt. The maximum values were saved to 300 mM for the production of siderophores (18.32%) and solubilization of phosphate (1061.49 μg mL-1) and 100 mM for the production of IAA (161 μg mL-1). A significant correlation existed between these three activities. These results showed that P. agglomerans lma2 with its Plant Growth Promoting Rhizobacteria (PGPR) and halophilic properties could constitute a good fertilizer in arid and saline zone.
PDF Abstract XML References Citation
Received: February 25, 2012;
Accepted: June 18, 2012;
Published: July 12, 2012
How to cite this article
H. Silini-Cherif, A. Silini, M. Ghoul and S. Yadav, 2012. Isolation and Characterization of Plant Growth Promoting Traits of a Rhizobacteria: Pantoea agglomerans lma2. Pakistan Journal of Biological Sciences, 15: 267-276.
DOI: 10.3923/pjbs.2012.267.276
URL: https://scialert.net/abstract/?doi=pjbs.2012.267.276
DOI: 10.3923/pjbs.2012.267.276
URL: https://scialert.net/abstract/?doi=pjbs.2012.267.276
INTRODUCTION
Plant growth is strongly influenced by many biotic and abiotic factors. Salinity is one of the major factors limiting plant productivity and therefore agricultural production. In worldwide, 340 million ha of agricultural land are affected by salinity among them 23% of cultivated land (Cheverry, 1995) of which 3.2 million ha in Algeria (Hamdy, 1999). This salinization is mainly found in arid and semi arid lands of the country. It leads to the depletion of soil organic matter and accumulation of toxic ions.
Research on the use of rhizobacteria to promote plant growth has increased dramatically over the last few years due to potential benefits observed in the use of PGPR in field conditions especially in saline soils. However, inoculation of stressed plants by PGPR strains alleviates salt stress (Ashraf et al., 2008; Saharan and Nehra, 2011). The growth of halotolerant microorganisms associated with plant roots can lead to improve fertility in saline soils (Hallmann et al., 1997; Alizadeh et al., 2012). As a result, the rhizobacteria of saline soils are able to grow at different salinity levels between 0 and 5% NaCl (Tripathi et al., 1998).
The mechanisms by which these rhizobacteria enhance plant growth are numerous, which include production of plant growth-regulating substances, phytohormones, suppression of plant pathogens through antibiosis, siderophore production, nitrogen fixation, mineralization of organic phosphorus, etc. (Kloepper and Beauchamp, 1992; Glick, 1995; Verma et al., 2010). The PGPR have usually more than two or three activities which act in a synergistic manner (Joseph et al., 2007; Yasmin et al., 2007).
Actually, a diverse array of bacteria are identified as PGPR (Kloepper and Beauchamp, 1992).
This diversity is due to the numerous studies to understanding different mechanisms of action of these PGPR. The bacterial species including Pseudomonas, Azospirillum, Azotobacter, Klebsiella, Enterobacter, Alcaligenes, Arthrobacter, Burkholderia, Pantoea, Serratia and Bacillus have shown a capacity to enhance plant growth (Glick, 1995; Nezarat and Gholami, 2009).
The PGPR used as biofertilizer and/or antagonist against plant pathogens, are a promising alternative to chemical fertilizers and pesticides. The plant-microbe interactions depend upon plant nutrient status in soil, soil environment, plant defence mechanism and the type of microorganism proliferating in the rhizosphere zone. However, the bacteria must be able to colonize roots and survive in the soil (Normander and Prosser, 2000; Woyessa and Assefa, 2011). Consequently, the selection and use of PGPR should consider adaptation of the inoculant to plant and to a particular ecosystem. In addition, the selection of effective PGPR strains is related to the characterization of properties of plant growth (Cattelan et al., 1999).
The objective of this study was to do a biochemical and physiological characterization of a rhizobacteria isolated from an arid soil and to determine its capabilities to improve plant growth under salt stress.
MATERIALS AND METHODS
Isolation of bacterial strain: The sample was collected from the rhizosphere of wheat in an arid soil located in the region of Bou-Saada, 250 km south of Algiers (Algeria). One gram of soil strongly adhering to the roots was extracted from the sample, added to 10 mL of sterile distilled water and shaken for 30 min. To select the nitrogen-fixing bacteria, the isolation was carried out on nitrogen-free medium: the Winogradsky Salt (WS) medium, incubated at 30°C/48 h. Typical colonies were subcultured several times on nutrient agar to obtain pure cultures. To select rhizobacteria tolerant high salt concentrations and able to producing PGP traits, the bacterial strain studied was chosen based on its best growth in presence of salt.
Identification of the bacterial strain: The bacterium was identified according to macroscopic appearance (appearance of the colony on solid medium, form, texture and pigmentation), Gram staining, mobility, oxidase, catalase and nitrate reductase tests. These tests were followed by identification using a biochemical API20E system (BioMerieux).
16S rRNA gene sequence analysis: PCR fragments obtained by the amplification of a DNA fragment corresponding to a region of the 16S rDNA gene of the isolate were sequenced using the automatic sequencer at DNA Vision Company (http:/www.dnavision.com). The sequence was submitted to the GenBank, and accession number was assigned GQ 478021. The partial 16S rDNA sequence of the isolate strain was compared with those available in the databases. The phylogenetic tree was constructed on the aligned datasets using the Neighbor-Joining method (Saitou and Nei, 1987). Phylogenetic analyses were conducted in MEGA4 (Tamura et al., 2007).
Biochemical characterization: For phenotypic screening for the bacterial strain, several carbon sources were used. Monosaccharides (D-glucose, D-galactose, D-fructose, D-xylose, D-ribose, raffinose, D-mannose, fucose and L-sorbose), disaccharides (lactose, maltose, trehalose, levulose and D-cellobiose) and derivatives sugar (glycerol, D-mannitol, dulcitol, adonitol, D-sorbitol, salicin and dextrin, citrate) were evaluated on minimal salt medium (Brown and Dilworth, 1975). Polysaccharides (starch and esculin), lipids (lecithin, tributyrin, Tween 20 and 80), proteins (casein, gelatin) and organic acids (L(-)malate, L(+)lactate, L(+)tartrate, succinate and maleate) were evaluated on corresponding media using standard methods (Cappuccino and Sherman, 1992). Growth on KCN was determined on medium containing 0.1 mL of potassium cyanide at 5%.
Effect of salt, pH and temperature: Salt tolerance was evaluated on nutrient broth in the presence of increasing concentrations ranging from 0 mM of NaCl to 1 M. The effect of pH was tested on nutrient broth at different pH values 4, 5, 6, 7, 8, 9 and 10. The media were inoculated with 100 μL of culture and incubated at 30°C/48 h. The experiments were performed in triplicate. The Optical Density (OD) was measured at 600 nm.
The ability of the strain to grow at different temperatures was tested on nutrient broth inoculated with 100 μL of culture and incubated at 4, 30, 37, 41 and 44°C/48 h. Growth was measured using a spectrophotometer at 600 nm. A triplicate was performed for each temperature of incubation.
Measurement of PGPR activities
Production of hydrogen cyanide (HCN): The ability of the isolated strain to produce HCN was measured using the method of Lorck (1948) on a HCN medium (nutrient agar supplemented with 4.4 g of glycine L-1) inoculated with the bacterial culture. Whatman paper 9 cm in diameter impregnated with a solution of sodium picrate (5% picric acid and 2% sodium carbonate) was deposited in the lid of the Petri dish which sealed with parafilm and incubated at 30°C/4 h. The development of color orange to red indicated the production of HCN.
Production of NH3: The production of NH3 was evaluated on peptone water inoculated with 100 μL of the bacterial culture (Cappuccino and Sherman, 1992) incubated at 30°C/48 h. The addition of 0.5 mL of Nessler's reagent which gives a color yellow to brown indicated the production of NH3.
Solubilization of phosphates: The ability of the strain to solubilize phosphates was tested on Pikovskaya (PVK) medium containing tricalcium phosphate (Ca3HPO4) as the sole source of phosphate. A volume of 10 μL of the bacterial culture was deposited as a spot on the surface of the PVK agar as described by Gaur (1990). After incubation at 30°C/7 days, the diameter of the halo around the colony was measured. Quantitative analysis of tricalcium phosphate solubilization in liquid medium was carried out on PVK liquid inoculated with 100 μL of culture and incubated at 30°C/4 h. The cultures were then centrifuged at 3000 rpm/15 min. The amount of soluble phosphate was measured by the colorimetric method of Olsen (Olsen and Sommers, 1982). The concentration of phosphate was determined by the absorbance of the color blue at 610 nm. A standard calibration curve was performed with a solution of KH2PO4. Three repetitions were performed.
Production of indole acetic acid (IAA): The production of IAA was tested on Winogradsky broth supplemented with 2 g L-1 tryptophan. The different media were inoculated with 100 μL of bacterial culture and incubated at 30°C/48 h. The colorimetric assay was performed using the method of Loper and Scroth (1986). The cultures were centrifuged at 3000 rpm/20 min. Two milliliter of the supernatant were mixed with 4 mL of Salkowski reagent (50 mL of perchloric acid and 1 mL of 35c/o FeCl3 0.5 M). The OD was measured at 530 nm. Concentrations of IAA were determined using a calibration curve of a solution of IAA obtained in the range 0 to 10-3 M. A triplicate was performed.
Production of siderophores: The production of siderophores was tested in medium Chrome Azurol S (CAS) (Schwyn and Neilands, 1987). The King B medium, given its composition free of iron, was used to demonstrate the production of siderophores. The King B solid medium was inoculated with 10 μL of bacterial culture and incubated at 30°C/24 h. After growth, 15 mL of CAS agar were poured on the bacterial culture. Contact after a few hours, a change of color from blue to orange, appeared around the colony producing siderophores. The diameter of orange halo was determined by subtracting the diameter of the colony of the total diameter (halo+colony). Quantitative analysis was performed on King B liquid medium inoculated with 100 μL of culture, incubated at 30°C/48 h. Cultures were centrifuged at 3000 rpm/30 min and 500 μL of the supernatant were mixed with 500 μL of CAS solution. The color changed from blue to orange at the rate of production of siderophores. The OD was measured by a spectrophotometer at 630 nm after 20 min of incubation. The experiment was performed in triplicate.
The percentage of siderophores was calculated using the following formula (Gokarn, 2010):
St = Se/St x 100
Where:
| St | = | CAS solution of color intense blue (control) |
| Se | = | Solution of the sample of less blue to orange depending on the intensity of production |
Effect of salt on the activities PGPR: The strain was tested for its ability to solubilize phosphates and to produce siderophores and IAA under salt stress. Each corresponding medium described above was supplemented with increasing concentrations of NaCl (from 0 to 1000 mM), inoculated with 100 μL of culture and incubated at 30°C/48 h. The quantitative estimate for each activity was determined by the methods described previously.
Statistical analysis: Each data was the mean of three replicates. All data were subjected by one-way analysis of variance and the mean differences were compared by Lowest Standard Deviations (LSD) test. Comparisons with p<0.05 were considered significantly different. All figures the spread values were shown as error bars representing standard errors of the means. The relationship between the three activities under salt stress conditions (IAA and, siderophore productions and phosphates solubilization) was examined using regression analysis.
RESULTS
Identification of the strain: The isolated strain grew on nutrient agar (GN) after 24 h at 30°C. The colonies obtained had the following macroscopic characteristics: round, smooth, regular edge, more or less flat, less than 1 mm diameter and produce a yellow pigment. Microscopic examination revealed rights bacilli, Gram-negative with rounded ends 3 to 5 μm in length and 0.5 to 1 μm wide. They were presented in isolation or in pairs. The strain was mobile, asporulιe, catalase+, oxidase-, nitrate reductase+(state N2), facultative anaerobic, fermented glucose without gas production (Table 1).
| Table 1: | Biochemical characterization of P. agglomerans lma2 |
 | |
| +: Positive test, -: Negative test | |
These characters mentioned, oriented us toward the family of Enterobacteriaceae. The API20E system confirmed biochemical identification and the result have determined the specie: Pantoea agglomerans.
The 16S rDNA sequencing and phylogenetic analysis According to the sequence analysis of the 16S rDNA partial sequences of the strain Pantoea agglomerans lma2 (accession number GQ 478021) was placed within the Pantoea cluster with 97% 16S rDNA similarity (Fig. 1).
Biochemical characterization: The results of various biochemical tests were listed in Table 1. The strain P. agglomerans lma2 degraded several sources of carbon sugars (D-fructose, D-galactose, D-xylose, D-ribose, D-mannose, maltose, trehalose, levulose, cellobiose, glycerol, D-mannitol, dulcitol, adonitol, salicin, dextrin, citrate and esculin), lipids (lecithin, tributyrin and Tween 80) proteins (gelatin, casein) and organic acids (malate, lactate and maleate) and grew on KCN (Table 1).
Physiological characterization
Effect of pH: Growth of P. agglomerans lma2 revealed that it grew over a wide pH range from pH 4 to pH 8. The inhibition of growth was visible to alkaline pH (pH 9 and 10). There was a maximum growth at pH 7 (Fig. 2).
Effect of temperature: The measure of turbidity indicated a growth of P. agglomerans lma2 at different temperatures. However, this rate varied according to temperature. It is significantly higher at T = 30°C but reduced to 37°C. At extreme temperatures (4, 41 and 44°C), the strain retained a significant but low growth ability (Fig. 3).
Effect of salt: The growth of the strain on nutrient broth with salt concentrations ranging from 0 to 1000 mM showed a good capacity for salt tolerance. Growth was observed up to 1 M with high rates at 100, 200 and 300 mM NaCl (Fig. 4).
Activities promoting growth plant
Nitrogen fixation, production of NH3 and HCN: Given its ability to grow on nitrogen-free medium like the WS medium, P. agglomerans lma2 was nitrogen fixing. This strain produced a yellow-brown color after addition of Nessler's reagent after 4 days of incubation in peptone water, indicating the production of NH3. The cyanogen activity was absent in P. agglomerans lma2. After 4 days at 30°C, the color of the filter paper remained unchanged (Table 2).
Solubilization of phosphate: P. agglomerans lma2, tested for its ability to solubilize Ca3HPO4 on PVK solid medium, produced a clear zone around the colony of 10 mm in diameter (Fig. 5). The quantitative estimation of soluble phosphate on PVK liquid medium was 809.19 μg mL-1 (Table 2).
Production of siderophores: The production of siderophores by Pantoea agglomerans lma2 on CAS solid medium after 24 h at 30°C was characterized by an orange halo around the colony.
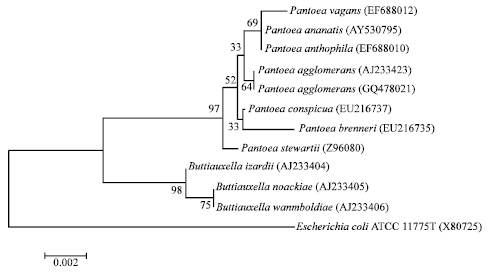 | |
| Fig. 1: | Unrooted phylogenetic tree based on a comparison of the 16S ribosomal DNA sequence of Pantoea agglomerans lma2 (GQ478021) and some of their closest phylogenetic relatives (Validly published strains), The tree was created by the neighbor-joining method, The numbers on the tree indicates the percentages of bootstrap sampling derived from 1,000 replications, Bar inferred nucleotide substitutions per nucleotides |
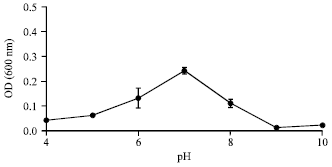 | |
| Fig. 2: | Effect of pH on growth of P. agglomerans lma2, OD: Optical density at 600 nm |
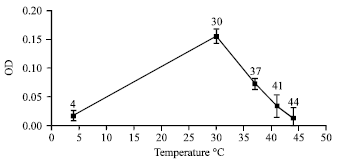 | |
| Fig. 3: | Effect of temperature on growth of P. agglomerans lma2, OD: Optical density at 600 nm |
The diameter of the halo was 5 cm (Fig. 6) (Table 2). The percentage of production of siderophores in CAS liquid medium was calculated by the difference in OD at 630 nm between the sample and the control. The color will turn from blue (control) to orange more or less intense depending on the rate of production. This strain produced 4,195% of siderophores (Table 2).
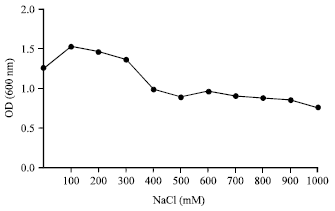 | |
| Fig. 4: | Effect of salt on growth of P. agglomerans lma2 |
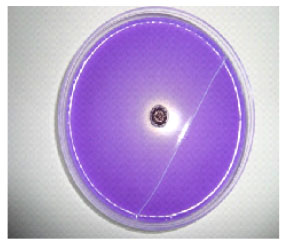 | |
| Fig. 5: | Clearing zone around the colony of P. agglomerans lma2 indicating the ability of the isolate to solubilize phosphate in PVK agar medium |
 | |
| Fig. 6: | Orange halo around the colony of P. agglomerans lma2 indicating the ability of this isolate to excrete siderophores |
| Table 2: | PGP activities of P. agglomerans lma2 |
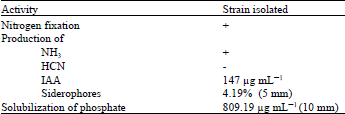 | |
| +: Positive test, -: Negative test, Parentheses values are diameter of halo on solid medium | |
Production of indole acetic acid (IAA): The IAA was produced from a precursor the tryptophan. The production of IAA by the isolated strain was measured after 4 days of incubation. The production rate was 147 μg mL-1 (Table 2).
Effect of salt on the activities of P. agglomerans lma2 In the presence of increasing concentrations of NaCl (from 0 to 1 M), the amounts of P solubilized by P. agglomerans lma2 varied from 809.19 to 1061.49 μg mL-1. It showed a higher solubility in the presence of salt from 100 to 800 mM compared to the control without salt, the maximum rate of soluble phosphate was attained in presence of 300 mM at 1061.49 μg mL-1. The rate of soluble phosphate decreased but remained significant at 900 and 1000 mM NaCl measuring 721.6 μg mL-1 and 583.90 μg mL-1, respectively (Fig. 7).
The production of siderophores was increased in the presence of salt compared to the control without salt (4.195%). The production was 12.01% at 100 to 900 mM to 8.16%, the maximum rate was 18.32% at 300 mM. At 1 M of NaCl the percentage was decreased and was 2.165% (Fig. 8).
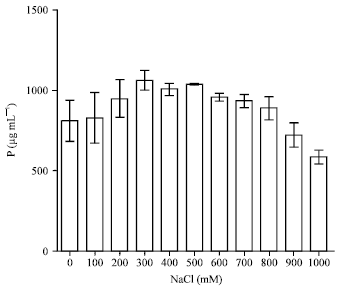 | |
| Fig. 7: | Phosphate solubilization by P. agglomerans lma2 under NaCl stress |
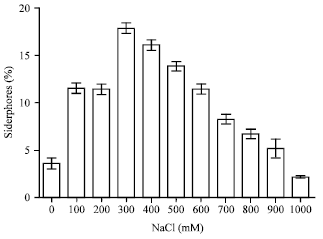 | |
| Fig. 8: | Siderophores production by P. agglomerans lma2 under NaCl stress |
The production capacity of the IAA by P. agglomerans lma2 was also enhanced in the presence of NaCl. The maximum was observed at 200 mM of NaCl (161 μg mL-1). The production decreased at 400 mM. At high concentrations 700, 800, 900 and 1000 mM NaCl, IAA production was very low (Fig. 9, 10).
Correlation between the activities of P. agglomerans lma2: The analysis of the correlation between the different activities of P. agglomerans lma2 showed a significant relationship at p<0.01 between production of siderophores and the solubilization of phosphates.
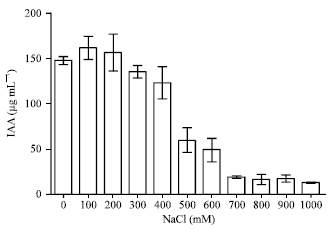 | |
| Fig. 9: | IAA production by P. agglomerans lma2 under NaCl stress |
 | |
| Fig. 10: | Pink color of the solutions indicating the IAA production by P. agglomerans lma2 under NaCl stress |
| Table 3: | Correlation between activities of P. agglomerans lma2 |
 | |
| **Significant at 1% probability level at p<0.01, *Significant at 5% probability level at 0.01≤p<0.05 | |
The correlation between the production of IAA and siderophores and between phosphate solubilization and the production of IAA was significant at p <0.05 (Table 3).
DISCUSSION
Selected strain was Gram-negative rod, facultative anaerobic, catalase positive, oxidase negative, mobile, fermenting glucose without gas production. The identification of this strain by the API20E system gave the specie P. agglomerans. The analysis of the 16S rDNA gene sequence confirmed this identification. On the basis of biochemical tests and by comparing the isolated strain P. agglomerans lma2 to biochemical characteristics of type strain P. agglomerans ATCC 27155 (Gavini et al., 1989), two differences revealed: the dulcitol use and reduction of nitrate in the state of molecular nitrogen. However, the sequence showed a very significant degree of homology (97%) with related species.
Reinhardt et al. (2008) suggested that several genera of Enterobacteriaceae such as P. agglomerans were beneficial to plants. These bacteria were able to use a wide variety of carbon sources as nutrients in t he rhizosphere (Misko and Germida, 2002). On the other hand, metabolic profile of strains allowed us to understand their competitive ability and survival under various environmental conditions (Francis et al., 2000; Giongo et al., 2010).
In this study, the optimum growth temperature was 30°C, indicating that Pantoea agglomerans lma2 was a mesophilic bacterium. In comparison with the type-strain, the strain grew equally well at 4°C after 4 days (Gavini et al., 1989). However, growth at 41°C, in contrast to the type strain, could explain the presence of these organisms in arid regions. Temperature was one of the most important factors governing the physiology and growth of microorganisms as reported by Rahman et al. (2006).
The effect of pH on the growth of P. agglomerans lma2 showed an optimum growth at pH 7. However, growth at pH slightly alkaline (pH 8) and acidic (pH 4-5) was noted. This explained the ability of the strain to grow on a wide pH range. This result was consistent with that of P. agglomerans CPA-2 that could grow at pH 5 to pH 8.6 (Costa et al., 2002).
According to Borneman et al. (1996), soil salinity played a prominent role in microbial process selection. Several studies indicated that bacteria isolated from saline or arid environments were more able to survive inhibitory salt concentrations compared to those isolated from non-saline habitats (Tripathi et al., 1998; Jat and Sharma, 2006). This was the case of P. agglomerans lma2 which our results showed that, this strain was halotolerant, it had a high growth in the presence of NaCl concentrations between 100 and 400 mM.
Rhizobacteria affected plant growth by improving their mineral supply phosphorus (P). The ability of some microorganisms to convert insoluble phosphorus in a soluble form was a significant beneficial effect for increasing yields of plants. Microorganisms solubilize P by the production of organic acids and/or secretion of H+. Therefore, P can be released by the substitution of protons or chelation with Ca2+ (Illmer and Schinner, 1995). The solubilization of P was a very common character in P. agglomerans lma2 . The amount of P solubilized on PVK medium supplemented with Ca3 (PO4)2 was 809.19 μg L-1. According Sulbaran et al. (2008), the amount of P solubilized by P. agglomerans MMB051 varied with the source of P present in the medium. It was high with the Ca3 (PO4)2 (95.75±2.94 μg L-1) but low with FePO4: (0.97±0.06 μg L-1), AlPO4: (3, 11±0.24 μg L-1) and KH2PO4 (6.69±0.45 μg L-1).
The strain P. agglomerans lma2 tested in the presence of salt showed a high solubilization capacity. This indicated that this strain had a character of resistance to salinity. These results were in agreement with those obtained by Son et al. (2006), indicating that P. agglomerans R-42 produced 900 μg L-1 of soluble P in optimum conditions. Moreover, this strain showed resistance against different environmental stresses such as salt concentration (1-5%) and solubilized P at higher levels CaHPO4 (1367 μg L-1), hydroxy apatite (1357 μg L-1), Ca3 (PO4) 2 (1312 μg L-1).
Auxin was the most efficient plant growth hormone, and among them the indole acetic acid was the most common. About 80% of rhizosphere bacteria were capable of producing indole 3-acetic acid. L-tryptophan was considered the precursor of IAA, because its addition was necessary for the production of IAA (Dastager et al., 2010; Keyeo et al., 2011). Strains that produce large amounts of auxin in the soil caused a maximum increase of the growth and crop yield (Khalid et al., 2004; Sarkar et al., 2002; Sudha et al., 2012). According Barazani and Friedmann (1999) bacteria, able to secrete a higher rate to 13.5 μg L-1 of indole compounds were considered as PGPR.
Production of IAA by P. agglosmerans lma2 was high and increased with salt. The maximum rate of product was 161 μg mL-1 at 300 mM NaCl. This behavior seemed to be due to the ability of the strain to resist osmotic stress. Indeed, the stressful environment fostered the development of bacteria showing the best activity (Banerjee et al., 2010). The comparison between the above results it was concluded that the strain isolated in this study was very effective in the production of IAA.
The volatile compounds were involved in the suppression of various pathogens (Howell et al., 1988). These authors noted that the ammonia produced as an intermediate in the catabolism of amino acids, root exudates and assimilated by the bacteria was an inhibitor of plant pathogens (Yasari and Patwardhan, 2007). Another secondary metabolite produced by certain rhizobacteria was hydrogen cyanide. Although, this compound was a general metabolic inhibitor. It was synthesized and secreted by certain bacteria as a means to avoid predation or competition (Heydari et al., 2008). However, the isolated strain P. agglomerans lma2 did not have the ability to produce HCN This could be explained by the non-existence of genes (hcn) responsible for the production of this metabolite (Voisard et al., 1989; Laville et al., 1998). However, the NH3 was easily produced by P. agglomerans lma2 as for the majority of rhizobacteria as reported by Ahmad et al. (2008).
Another activity of PGPR was the production of siderophores that can influence plant growth by binding iron in its available form: Fe3+. Through this process, iron was made unavailable to plant pathogens. Therefore, siderophores protect the health of plants of several fungal or bacterial diseases (Siddiqui, 2005; Sahu and Sindhu, 2011). P. agglomerans lma2 was characterized by an important production of siderophores (large halo around the colony). P. agglomerans lma2 was very effective in siderophore production under salt stress and had substantially the same growth characteristics and adaptation to salinity than other activities.
The analysis showed that there was a significant correlation between the three activities of P. agglomerans lma2 tested in the presence of salt. Several studies on the performance of PGPR showed that rhizobacteria could have several activities that act synergistically to improve plant growth (Ahmad et al., 2008).
In addition, it should be noted the behavior of the isolated strain against salt, indeed, the increase of each activity improving plant growth was between 100 and 400 mM of NaCl concentration and indicated clearly its resistance to salinity and halophilic character.
Performance of activities of P. agglomerans lma2 was significantly better in the presence of salt.
These findings suggested that the strain P. agglomerans lma2 was halophilic and could be a good biofertilizer for plant growth in arid regions and regions with saline soils.
REFERENCES
- Ahmad, F., I. Ahmad and M.S. Khan, 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res., 168: 173-181.
CrossRefDirect Link - Alizadeh, O., S. Sharafzadeh and A.H. Firoozabadi, 2012. The effect of plant growth promoting rhizobacteria in saline condition. Asian J. Plant Sci., 11: 1-8.
CrossRef - Ashraf, M., H.R. Athar, P.J.C. Harris and T.R. Kwon, 2008. Some prospective strategies for improving crop salt tolerance. Adv. Agron., 97: 45-110.
CrossRefDirect Link - Banerjee, S., R. Palit, C. Sengupta and D. Stranding, 2010. Stress induced phosphate solubilization by Arthrobacter sp. And Bacillus sp. Isolated from tomato rhizosphere. Aust. J. Crop Sci., 4: 378-383.
Direct Link - Barazani, O. and J. Friedman, 1999. Is IAA the major root growth factor secreted from plant-growth-mediating bacteria. J. Chem. Ecol., 25: 2397-2406.
CrossRefDirect Link - Borneman, J., P.W. Skroch, K.M. O'Sullivan, J.A. Palus and N.G. Rumjanek et al., 1996. Molecular microbial diversity of an agricultural soil in Wisconsin. Applied Environ. Microbiol., 62: 1935-1943.
PubMedDirect Link - Cattelan, A.J., P.G. Hartel and J.J. Fuhrmann, 1999. Screening for plant growth-promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J., 63: 1670-1680.
CrossRefDirect Link - Costa E., J. Usall, N. Teixido, J. Delgado and I. Vinas, 2002. Water activity, temperature and pH effects on growth of the biocontrol agent Pantoea agglomerans CPA-2. Can. J. Microbiol., 48: 1082-1088.
CrossRef - Francis, C.A., A.Y. Obraztson and B.M. Tebo, 2000. Dissimilatory metal reduction by the facultative anaerobe Pantoea agglomerans SPI. Applied Environ. Microbiol., 66: 543-548.
Direct Link - Gavini, F., J. Mergaert, A. Beji, C. Mielcarek, D. Izard, K. Kersters and J. De Ley, 1989. Transfer of Enterobacter agglomerans (Beijerinck 1888), Ewing and Fife 1972 to Pantoea gen. nov. as Pantoea agglomerans comb. nov. and description of Pantoea dispersa sp. nov. Int. J. Syst. Evol. Bacteriol., 39: 337-345.
CrossRef - Giongo, A., A. Beneduzi, A. Ambrosini, L.K. Vargas and M.R. Stroschein et al., 2010. Isolation and characterization of two plant growth-promoting from the rhizoplane of a legume (Lupines albescens) in sandy soil. Rev. Bras. Cienc. Solo, 34: 361-369.
CrossRef - Glick, B.R., 1995. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol., 41: 109-117.
CrossRefDirect Link - Gokarn, K., 2010. Siderophores and Pathogenecity of Microorganisms. J. Biosci Tech., 1: 127-134.
Direct Link - Howell, C.R., R.C. Beier and R.D. Stipanovic, 1988. Production of ammonia by Enterobacter cloacae and its possible role in the biological control of Pythium pre-emergence damping-off by the bacterium. Phytopathology, 78: 1075-1078.
CrossRefDirect Link - Illmer, P. and F. Schinner, 1995. Solubilization of inorganic calcium phosphates-solublization mechanisms. Soil Biol. Biochem., 27: 257-263.
CrossRefDirect Link - Jat, N.K. and V. Sharma, 2006. The interactive effect of salinity and pgr on certain bio-chemical parameters in wheat seedlings. Am. J. Plant Physiol., 1: 132-141.
CrossRefDirect Link - Joseph, B., R.R. Patra and R. Lawrence, 2007. Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). Int. J. Plant Prod., 1: 141-152.
CrossRefDirect Link - Keyeo, F., O.N. Ai'shah and H.G. Amir, 2011. The effects of nitrogen fixation activity and phytohormone production of diazotroph in promoting growth of rice seedlings. Biotechnol., 10: 267-273.
CrossRefDirect Link - Khalid, A., M. Arshad and Z.A. Zahir, 2004. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol., 96: 473-480.
CrossRefDirect Link - Kloepper, J.W. and C.J. Beauchamp, 1992. A review of issues related to measuring colonization of plant roots by bacteria. Can. J. Microbiol., 38: 1219-1232.
CrossRefDirect Link - Nezarat, S. and A. Gholami, 2009. Screening plant growth promoting rhizobacteria for improving seed germination, seedling growth and yield of maize. Pak. J. Biol. Sci., 12: 26-32.
CrossRefPubMedDirect Link - Normander, B. and J.I. Prosser, 2000. Bacterial origin and community composition in the barley phytosphere as a function of habitat and presowing conditions. Applied Environ. Microbiol., 66: 4372-4377.
CrossRef - Rahman, M., S. Mubassara, S. Hoque and Z. Khan, 2006. Effect of some environmental factors on the growth of azospirillum species isolated from saline soils of satkhira district, Bangladesh. Bangladesh J. Microbiol., 23: 145-148.
CrossRefDirect Link - Reinhardt, E.L., P.L. Ramos, G.P. Manfio, H.R. Barbosa, C. Pavan and C.A. Moreira-Filho, 2008. Molecular characterization of nitrogen-fixing bacteria isolated from Brazilian agricultural plant in Sao Paulo State. Braz. J. Microbiol., 39: 414-422.
CrossRef - Saharan, B.S. and V. Nehra, 2011. Plant Growth promoting rhizobacteria: A critical review. Life Sci. Med. Res.
Direct Link - Sahu, G.K. and S.S. Sindhu, 2011. Disease control and plant growth promotion of green gram by siderophore producing Pseudomonas sp. Res. J. Microbiol., 6: 735-749.
CrossRefDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Sarkar, P.K., M.S. Haque and M.A. Karim, 2002. Effects of GA3 and IAA and their frequency of application on morphology, yield contributing characters and yield of soybean. J. Agron., 1: 119-122.
CrossRefDirect Link - Schwyn, B. and J.B. Neilands, 1987. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem., 160: 47-56.
CrossRefPubMedDirect Link - Siddiqui, Z.A., 2005. PGPR: Prospective Biocontrol Agents of Plant Pathogens. In: PGPR: Biocontrol and Biofertilization, Siddiqui, Z.A. (Ed.), Springer, Dordrecht, Netherlands, ISBN: 978-1-4020-4002-3, pp: 111-142.
CrossRefDirect Link - Sudha, M., R.S. Gowri, P. Prabhavathi, P. Astapriya, S.Y. Devi and A. Saranya, 2012. Production and optimization of indole acetic acid by indigenous micro flora using agro waste as substrate. Pak. J. Biol. Sci., 15: 39-43.
CrossRef - Sulbaran, M., E. Perez, M. Ball, A. Bahsas and L.A. Yarzabal, 2008. Characterization of the Mineral Phosphate Solubilizing Activity of Pantoea agglomerans MMB051 Isolated from an Iron-Rich Soil in Southeastern Venezuela (Bolivar State). Curr. Microbiol., 58: 378-383.
CrossRef - Tripathi, A.K., B.M. Mishra and P. Tripathi, 1998. Salinity stress responses in the plant growth promoting rhizobacteria, Azospirillum sp. J. Biosci., 23: 463-471.
CrossRef - Verma, J.P., J. Yadav, K.N. Tiwari, Lavakush and V. Singh, 2010. Impact of plant growth promoting rhizobacteria on crop production. Int. J. Agric. Res., 5: 954-983.
CrossRefDirect Link - Yasari, E. and A.M. Patwardhan, 2007. Effects of (Azotobacter and Azosprillium) inoculants and chemical fertilizers on growth and productivity of canola (Brassica napus L.). Asian J. Plant Sci., 6: 77-82.
CrossRefDirect Link - Yasmin, F., O. Radziah, S. Mohd. Said and S. Kamaruzaman, 2007. Screening for beneficial properties of rhizobacteria isolated from sweetpotato rhizosphere. Biotechnology, 6: 49-52.
CrossRefDirect Link - Voisard, C., C. Keel, D. Haas and G. Defago, 1989. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. Eur. Mol. Biol. Organiz. J., 8: 351-358.
Direct Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Laville, J., C. Blumer, C. von Schroetter, V. Gaia, G. Defago, C. Keel and D. Haas, 1998. Characterization of the hcnABC gene cluster encoding hydrogen cyanide synthase and anaerobic regulation by ANR in the strictly aerobic biocontrol of Pseudomonas fluorescens. CHA0. J. Bacteriol., 180: 3187-3196.
PubMedDirect Link - Hallmann, J., A. Quadt-Hallmann, W.F. Mahaffee and J.W. Kloepper, 1997. Bacterial endophytes in agricultural crops. Can. J. Microbiol., 43: 895-914.
CrossRefDirect Link - Woyessa, D. and F. Assefa, 2011. Effect of plant growth promoting rhizobacteria on growth and yield of Tef (Eragrostis tef Zucc. Trotter) under greenhouse condition. Res. J. Microbiol., 6: 343-355.
CrossRef - Son, H.J., G.T. Park, M.S. Cha and M.S. Heo, 2006. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-42 isolated from soybean rhizosphere. Bioresour. Technol., 97: 204-210.
CrossRefDirect Link - Loper, J.E. and M.N. Schroth, 1986. Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology, 76: 386-389.
CrossRefDirect Link