
Mariam A. Abu- Al-Basal
Department of Biological Sciences, Faculty of Science, Al-al-Bayt University, P.O. Box 130040, Mafraq 25113, Jordan
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 23 | Page No.: 1038-1046







ABSTRACT
Nigella sativa has been used for a long time in Jordanian folk medicine to treat skin diseases like microbial infections and inflammation. Therefore, the present study was conducted to assess the healing efficacy of petroleum ether extract of Nigella sativa seeds (fixed oil) on staphylococcal-infected skin. Male BALB/c mice were infected with 100 μL of Staphylococcus aureus (ATCC 6538) suspension at a dose of 108 colony forming unit/mouse into shaved mild dorsal skin. Application of treatments for each group (100 μL sterile saline, 100 μL chloramphenicol (10 μg/mouse) and Nigella sativa fixed oil at a dose of 50, 100 or 150 μL/mouse) was performed at the site of infection twice a day for two consecutive days after 3 h of infection. At day 3 and 5 after infection, total White Blood Cells (WBCs) count; differential and absolute differential WBC counts and the number of viable bacteria present in the skin area were measured. At day 5 after infection, the animals were sacrificed and the histology of skin was examined. Results indicated that fixed oil of Nigella sativa seeds enhance healing of staphylococcal-infected skin by reducing total and absolute differential WBC counts, local infection and inflammation, bacterial expansion and tissue impairment. These effects provide scientific basis for the use of Nigella sativa in traditional medicine to treat skin infections and inflammations.
PDF Abstract XML References Citation
Received: July 01, 2011;
Accepted: October 14, 2011;
Published: December 02, 2011
How to cite this article
Mariam A. Abu- Al-Basal, 2011. Influence of Nigella sativa Fixed Oil on Some Blood Parameters and Histopathology of Skin in Staphylococcal-Infected BALB/c Mice. Pakistan Journal of Biological Sciences, 14: 1038-1046.
DOI: 10.3923/pjbs.2011.1038.1046
URL: https://scialert.net/abstract/?doi=pjbs.2011.1038.1046
DOI: 10.3923/pjbs.2011.1038.1046
URL: https://scialert.net/abstract/?doi=pjbs.2011.1038.1046
INTRODUCTION
Nigella sativa L. (Family Ranunculaceae), commonly known as black seed, is an annual herbaceous plant cultivated in different parts of the world, mainly in countries bordering the Mediterranean Sea (Bourgou et al., 2010). Seeds of Nigella sativa are frequently added to bread and pickles as a flavouring agent. They have been used in Jordan and Palestine, as a natural remedy, for health promotion and also to treat many diseases and ailments. For example, fever, allergies, arthritis, hypercholesterolemia, lung diseases, common cold, high blood pressure, immune disorders and skin diseases including psoriasis, eczema, hair loss, microbial infections and inflammations (Oran and Al-Eisawi, 1998; Lev and Amar, 2002; Said et al., 2002; Abu-Irmaileh and Afifi, 2003). Previous studies confirmed that the seed extract of Nigella sativa possesses a remarkable effect in the treatment of many health problems. These include immune stimulation (Salem, 2005), anti-inflammatory (Ali and Blunden, 2003; El Gazzar et al., 2006), anti-oxidant (Burits and Bucar, 2000), anti-microbial (Mashhadian and Rakhshandeh, 2005), anti-tumor (Amara et al., 2008) and wound healing stimulatory effects (Abu-Al-Basal, 2001).
Staphylococcus aureus is one of the most important human and veterinary pathogens that cause infections ranging from benign to life threatening diseases (Francois et al., 2010). The high incidence of methicillin resistance in hospitals and the emergence of vancomycin-intermediate Staphylococcus aureus complicated the prevention and treatment of serious infections due to staphylococci (Griffith et al., 2003). Skin and soft tissue abscesses and infected cysts are among the most common infections caused by these organisms that may lead to serious local and systemic complications (Brook, 2010). A number of experimental models of staphylococcal skin infections have been previously described and these showed that bacteria can readily invade the epidermis and dermis to produce localized infection and cause a variety of pathologic changes in the skin, including impetigo, furuncles, subcutaneous abscesses, scalded skin syndrome and necrotizing fasciitis (Hahn et al., 2009). In fact, infections due to multi-resistant Gram-positive organisms are increasing in frequency and hence their early recognition, proper medical treatment and surgical management are of primary importance and urgently required. Consequently, new treatment strategies is crucial to deal with this emerging issue involving the use of medicinal plants as a source for new therapeutic agents aimed at the management and treatment of skin infections.
The oil and active ingredients of Nigella sativa seeds have been proved to exert promising inhibitory effects against many strains of bacteria, including those known to be highly resistance to drugs. (Hanafy and Hatem, 1991; Salman et al., 2008). In a recent study, organic solvents were used to extract fixed oil constituents from seeds of Nigella sativa including petroleum ether. That had the most powerful significant inhibitory effects both in vitro and in vivo against clinical isolates from patients with skin wound infections (Abu-Al-Basal, 2009). As a matter of fact, in vivo studies on strains of pathogenic microbes are scarce and fixed oil extracted by petroleum ether was never used before at least with the seeds of Nigella sativa. Therefore, in a continuation with efforts of previous study, the present research was followed for investigating the effect of petroleum ether extract of Nigella sativa seeds on some blood parameters and histopathology of skin in Staphylococcus aureus infected BALB/c mice.
MATERIALS AND METHODS
Plant selection: Seeds of Nigella sativa were purchased in March, 2011 from a herbal shop in Irbid, Jordan. The seeds were authenticated at the Herbarium of the Department of Biological Sciences, University of Jordan, Amman, Jordan. A voucher specimen (NO.NSJD-3-011) has been deposited in the Department of Biological Sciences, Faculty of Science, Al-al-Bayt University, Mafraq, Jordan for future reference.
Plant extract: Seeds of Nigella sativa were ground into slightly coarse powder using electric blender. Fixed oil was prepared by soaking 100 g of the dried powder in 500 mL of petroleum ether (40-60°C) using a conical flask plugged with cotton wool. The mixture was kept at room temperature for 48 h under continuous shaking (memmert shaker SV 1422, Germany) and filtered through Whatman filter paper No. 2 under vacuum. The filtrate was evaporated to dryness by Rota vapor (Ruchl R-114, Switzerland), where the rotary water bath (Ruchl B-480) was adjusted to 55°C. The extract was kept overnight under vacuum fume hood to obtain a constant dry weight and later stored in closed vessel at 4°C in a refrigerator for further use.
Animals: Male BALB/c mice six week of age (18-20 g) were obtained from the animal house of the Department of Biological Sciences, Yarmouk University, Irbid, Jordan. Mice were kept under specific pathogenic-free conditions, housed, fed and treated in accordance with the international guidelines principles of laboratory animal use and care (Hedrich and Bullock, 2006). They were maintained on standard pellet diet and water ad libitum for two weeks to be acclimatized prior to the investigation.
Bacterial strain: Staphylococcus aureus (ATCC 6538) was kindly obtained from Dar Al Dawa, Na our, Jordan. Bacterial culture was routinely grown in nutrient broth at 37°C for 24 h. Active culture was harvested by centrifugation, washed twice and resuspended in sterile saline. The bacterial suspension was then diluted with sterile saline to 1x109 colony forming unit (CFU) mL-1 according to method of Kuo et al. (2005).
Infection of animals: Infection of BALB/c mice with 100 mL of Staphylococcus aureus (ATCC 6538) suspension at a dose of 1 x 108 CFU/mouse was conducted into shaved mid dorsal skin as described by Godin et al. (2005).
Mice grouping and treatment: Infected mice were randomly divided into control and experimental groups of twelve mice each. Various doses of N. sativa fixed oil were prepared and preliminary tested for their tolerance in BALB/c mice to select the optimum dose intended for the treatment of experimental animals. Application of treatments for control or experimental groups was performed at the site of primary infection by cutaneous injection twice a day for two consecutive days using insulin syringes, after 3 h of infection. Group I: uninfected mice left without treatment, as normal control. Group II: infected mice treated with vehicle (100 μL sterile saline), as negative control. Group III: infected mice treated with 100 μL chloramphenicol antibiotic solution (10 μg/mouse dissolved in sterile saline), as positive control. Group IV: infected mice treated with 50 μL pure fixed oil. Group V: infected mice treated with 100 μL pure fixed oil. Group VI: infected mice treated with 150 μL pure fixed oil. Mice were housed and maintained on normal food and water ad libitum during the whole period of experiment.
Blood sampling and hematological methods: All blood samples were withdrawn via cardiac puncture at day 3 and 5 after infection and analyzed for total, differential and absolute differential White Blood Cell (WBC) counts by manual standard hematological methods as described by Doeing et al. (2003) and McGarry et al. (2010).
Bacterial counts: Counts of viable bacteria in mice infected skin at day 3 and 5 after infection was performed as previously described by Kuo et al. (2005) and Godin et al. (2005).
Behavioral responses of mice: Behavioral responses of mice were followed as proposed by Kugelberg et al. (2005). The mice were monitored at least twice a day for signs of fatigue, stress or aggressiveness and the weight was recorded before and after each experiment.
Histology: Skin samples from each group were obtained at day 5 after infection. Samples were dissected, fixed in 10% neutral formalin, dehydrated in ascending grades of alcohol and imbedded in paraffin wax. Five-micrometer sections were stained with hematoxylin and eosin for histological evaluation and Gram's crystal violet solution for the identification of bacteria under light microscope. Semi quantitative scoring system was carried out to characterize the size and density of bacteria presence, the inflammatory response and infiltrating cells in different layers of staphylococcal-infected skin (epidermis, dermis, subcutis and muscular tissue) in BALB/c mice as described previously by Kugelberg et al. (2005).
Statistical analysis: Results are expressed as Means± SEM (Standard Error of the Mean). Comparisons between groups were performed by using paired student's t-test on a statistical software package (SPSS). Differences were considered significant, if p value is less than 0.05.
RESULTS
Blood parameters: Staphylococcal-infected mice treated with Nigella sativa fixed oil exhibited a dose-dependent significant decrease (p<0.05) in total and absolute differential White Blood Cell (WBC) counts compared to those of control groups (Tables 1, 2). Remarkable reduction in total WBC counts was observed at a dose of 150 μL of Nigella sativa fixed oil/mouse (Group VI: 10.12±0.38x103 cells μL-1), when compared to vehicle-treated control (Group II: 15.48±0.21x103 cells μL-1), at day 3 after infection (Table 1). The same oil-treated group showed further significant decrease in total WBC counts (Group VI: 09.20±0.31x103 cells μL-1) which was very close to the level of normal control (Group I: 08.90±0.21x103 cells μL-1) but slightly higher than that of chloramphenicol-treated mice (Group III: 08.20±0.21x103 cells μL-1), at day 5 after infection (Table 2). Similar significant effect was noticed at the same dose of oil in absolute differential WBC counts of treated mice at day 3 and 5 after infection (Tables 1, 2). This was highly illuminated by considerable decrease in absolute differential counts of basophil to the level of normal control (Group I: 00.04±0.01x103 cells μL-1). Though, it was significantly much lower than those of chloramphenicol-treated mice (Group III: 00.07±0.02x103 cells μL-1) and vehicle-treated mice (Group II: 00.09±0.03x103 cells μL-1), at day 5 after infection (Table 2).
In vivo anti-bacterial effect: The effect of Nigella sativa fixed oil on Staphylococcus aureus (ATCC 6538) growth in the skin of mice at day 3 and 5 after infection was shown in Fig. 1. Counts of viable bacteria decreased considerably to a significant level (p<0.05) at a dose-dependent manner in Nigella sativa fixed oil-treated groups. The most significant anti-bacterial effect was observed at a dose of 150 μL fixed oil/mouse (bars labeled 5), when compared to control groups.
| Table 1: | Influence of Nigella sativa fixed oil on some blood parameters of staphylococcal-infected BALB/c mice at day 3 after cutaneous infection |
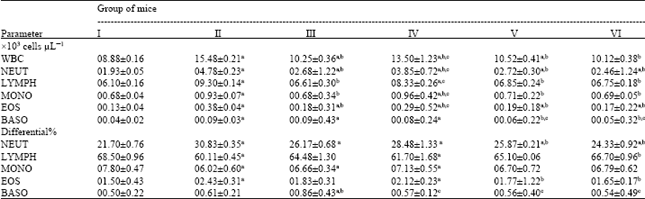 | |
| Data are expressed as Means±SEM for six mice in each group. I: Uninfected normal control; II: Infected vehicle-treated group (100 μL of sterile saline/mouse); III: infected chloramphenicol-treated group (100 μL of antibiotic solution at a dose of 10 μg/mouse); IV: Infected fixed oil-treated group (50 μL of Nigella sativa petroleum ether extract/mouse); V: Infected fixed oil-treated group (100 μL of Nigella sativa petroleum ether extract /mouse); VI: Infected fixed oil-treated group (150 μL of Nigella sativa petroleum ether extract/mouse). aStatistically significant when compared to uninfected normal control group (I) at p<0.05. b Statistically significant when compared to infected vehicle-treated group (II) at p<0.05. cStatistically significant when compared to infected chloramphenicol-treated group (III) at p<0.05 | |
| Table 2: | Influence of Nigella sativa fixed oil on some blood parameters of staphylococcal-infected BALB/c mice at day 5 after cutaneous infection |
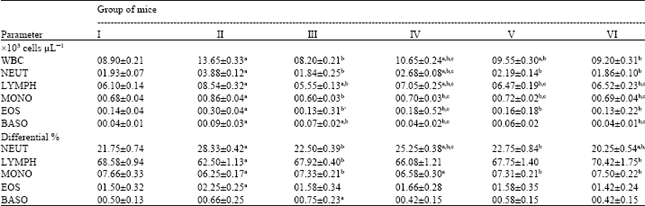 | |
| Data are expressed as Means±SEM for six mice in each group. I: Uninfected normal control; II: Infected vehicle-treated group (100 μL of sterile saline/mouse); III: infected chloramphenicol-treated group (100 μL of antibiotic solution at a dose of 10 μg/mouse); IV: Infected fixed oil-treated group (50 μL of Nigella sativa petroleum ether extract/mouse); V: Infected fixed oil-treated group (100 μL of Nigella sativa petroleum ether extract /mouse); VI: Infected fixed oil-treated group (150 μL of Nigella sativa petroleum ether extract /mouse). aStatistically significant when compared to uninfected normal control group (I) at p<0.05. bStatistically significant when compared to infected vehicle-treated group (II) at p<0.05. c Statistically significant when compared to infected chloramphenicol-treated group (III) at p<0.05 | |
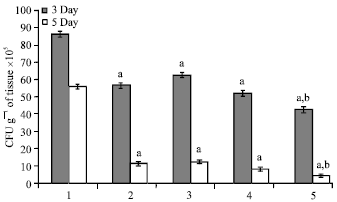 | |
| Fig. 1: | Counts of viable Staphylococcus aureus in the skin of BALB/c mice at day 3 and 5 after infection. Bars labeled 1: infected vehicle-treated group (100 μL of sterile saline/mouse). Bars labeled 2: infected chloramphenicol-treated group (100 μL of antibiotic solution at a dose of 10 μg/mouse). Bars labeled 3: infected fixed oil-treated group (50 μL of Nigella sativa petroleum ether extract/mouse). Bars labeled 4: infected fixed oil-treated group (100 μL of Nigella sativa petroleum ether extract/mouse). Bars labeled 5: infected fixed oil-treated group (150 μL of Nigella sativa petroleum ether extract/mouse). Data are expressed as Means±SEM for six mice in each group. a: Statistically significant when compared to infected vehicle-treated group (bars labeled 1) at p<0.05. b: Statistically significant when compared to infected chloramphenicol-treated group (bars labeled 2) at p<0.05 |
This potent dose of fixed oil displayed significant reduction in bacterial count reached to around 42x105 cfu g-1 of tissue, at day 3 after infection. In contrast, vehicle- and chloramphenicol-treated groups had elevated bacterial counts, to around 86x105 and 56x105 cfu g-1 of tissue, respectively. Similar effect was observed at day 5 after infection, yet the bacterial numbers declined further to a significant level in oil-treated group and reached approximately 4x105 cfu g-1 of tissue. On the contrary, mice treated with chloramphenicol had much higher bacterial counts (11x105 cfu g-1 of tissue) which indicates that Nigella sativa fixed oil is more effective in treating staphylococcal-infected skin.
DISCUSSION
In the present study, the significant anti-bacterial properties of the fixed oil (Fig. 1) could be related to potent bactericidal and/or immune stimulating/modulating agents present in the oil. Chemical analysis of petroleum ether extract of Nigella sativa seeds revealed the presence of considerable amount of fatty acids in the oil. Linoleic acid (55.6%) and oleic acid (23.4%) are the major components of unsaturated fatty acids that constituting 82.5% of the total fatty acids identified in the oil (Nickavar et al., 2003). Previous studies confirmed that lipids have an inhibitory effect against pathogenic infections in skin and mucous membranes. For example, long chain unsaturated fatty acids, medium chain saturated fatty acids and their monoglycerides are the most active against pathogenic bacteria and viruses by killing them rapidly in large amount.
| Table 3: | Microscopic evaluation of Nigella sativa fixed oil effects on the size and density of the inflammatory response, infiltrating cells and presence of bacteria in different layers of staphylococcal-infected skin of BALB/c at day 5 after infection |
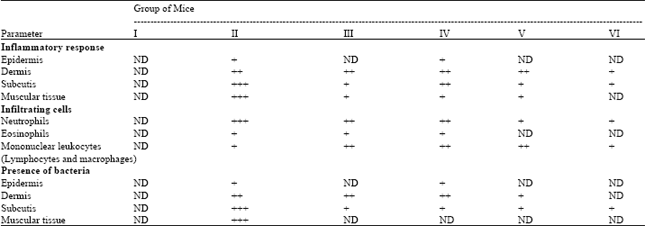 | |
| Data are expressed as Means±SEM for six mice in each group. I: Uninfected normal control; II: Infected vehicle-treated group (100 μL of sterile saline/mouse); III: Infected chloramphenicol-treated group (100 μL of antibiotic solution at a dose of 10 μg/mouse); IV: Infected fixed oil-treated group (50 μL of Nigella sativa petroleum ether extract/mouse); V: Infected fixed oil-treated group (100 μL of Nigella sativa petroleum ether extract /mouse); VI: Infected fixed oil-treated group (150 μL of Nigella sativa petroleum ether extract /mouse). ND: Not detected; +: Little/few; ++: Moderate; +++: Severe/abundant | |
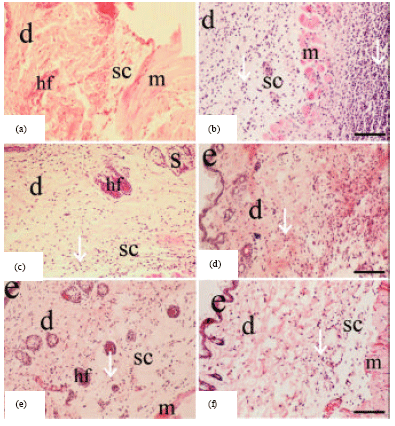 | |
| Fig. 2(a-f): | Hematoxylin and eosin histological sections of Staphylococcal-infected skin obtained from the controls and Nigella sativa treated BALB/c mice at day 5 after infection. (a) Uninfected normal skin; (b) infected vehicle-treated skin (100 μL of sterile saline/mouse); (c) infected chloramphenicol-treated skin (100 μL of antibiotic solution at a dose of 10 μg/mouse); (d) infected fixed oil-treated skin (50 μL of Nigella sativa petroleum ether extract/mouse); (e) infected fixed oil-treated skin (100 μL of Nigella sativa petroleum ether extract /mouse) and (f) infected fixed oil-treated skin (150 μL of Nigella sativa petroleum ether extract/mouse). Epidermis (e); dermis (d); hair follicle (hf); striated muscle (m), the subcutis (sc), a layer of adipose tissue; sebaceous gland (s); infiltration of inflammatory cells (white arrow head). Scale bar: 50 μm (a-f) |
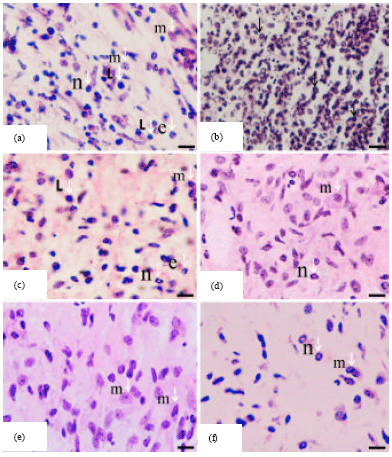 | |
| Fig. 3(a-f): | Hematoxylin and eosin histological sections of Staphylococcal-infected skin obtained from the controls and Nigella sativa treated BALB/c mice, revealing the persistence of inflammatory and immune cells at day 5 after infection. (a and b) Infected vehicle-treated skin (100 μL of sterile saline/mouse); (c) infected chloramphenicol-treated skin (100 μL of antibiotic solution at a dose of 10 μg/mouse); (d) infected fixed oil-treated skin (50 μL of Nigella sativa petroleum ether extract/mouse); (e) infected fixed oil-treated skin (100 μL of Nigella sativa petroleum ether extract /mouse); (f) infected fixed oil-treated skin (150 μL of Nigella sativa petroleum ether extract /mouse). Eosinophils (e); lymphocytes (l); active macrophages with variable appearance (m); neutrophils (n). Persistence of large numbers of inflammatory cells in considerably infected skin (black arrow head). Scale bar: 200 μm (a-f) |
This anti-bacterial effect of Free Fatty Acids (FFAs) noticed to be increased significantly by the addition of a double bond, and hence unsaturated fatty acids, such as linoleic acids (C18:2) are more effective against Gram-positive bacteria. A skin lipid was also found to destroy many species of microorganisms, for instance the FFAs of stratum corneum in the epidermis, that known to participate in skin immune defense by providing a physical barrier to bacterial invasion, exhibiting substantial anti-bacterial effect against Staphylococcus aureus (Thormar and Hilmarsson, 2007).
The significant changes in total and absolute differential WBC counts (Tables 1, 2) and marked histopathological observations (Table 3, Fig. 3) in fixed oil treated mice revealed improvement in healing staphylococcal-infected skin, when compared to infected vehicle-treated and normal untreated controls. However, the oil healing effect appeared to be dose dependent and the clear potentiality was typically observed at a dose of 150 μL fixed oil/mouse, which proved to be more effective than the standard drug, chloramphenicol in treating infection. This efficient dose of the oil attenuates bacterium-induced inflammation by reducing the size and density of inflammatory response, infiltrating cells and presence of bacteria in different layers of infected skin at day 5 after infection (Table 3, Fig. 3). Along with significant decrease in total and absolute differential WBC counts that reached near to normal levels in the treated mice (Table 2). Which indicate that the constituents of the oil might have an impact on the host-bacterium interrelationship, and as a consequence an effect on some mediators of inflammation. Such as oxidants, cytokines and lytic enzymes secreted by neutrophils and macrophages and part of the inflammatory response is the recruitment of these cells to site of infection (Salem, 2005). Reduced inflammation in the oil-treated mice seems also to be attributed to the inhibition of eicosanoid generation in leukocytes as suggested by the study of Houghton et al. (1995). On the contrary, acute inflammatory response was high-lighted by significant increase in total and absolute differential WBC counts (Table 2) along with marked dense infiltrate of cells, mainly neutrophils, to tissues of infected skin (Fig. 2) in the vehicle-treated mice at day 5 after infection, when compared to normal untreated mice. This may be attributed to the increase in the rate of neutrophil production at the bone marrow leading to neutrophil egress into tissue layers of infected skin that appeared intensely later at day 5 after infection, as a result of inflammation (Von Vietinghoff and Ley, 2008). Neutrophils are the first defense cells against invading bacteria and are rapidly recruited to the site of infection by chemotactic factors. The major roles played by neutrophils in inflammatory and immune responses are phagocytosis and killing of bacteria via the generation of reactive oxygen intermediates and the release of lytic enzymes stored in granules (Molne et al., 2000). Furthermore, marked histological findings in oil-treated mice (Table 3, Fig. 3) indicated that rapid recruitment of neutrophils and phagocytosis, in the initial stages of infection, is critical for the clearance and inferior expansion of infection. That was clearly demonstrated in vehicle-treated mice, in which the skin infection continues despite the abundant occurrence of neutrophilic granulocytes at late stage of inflammation (day 5 after infection) due to Staphylococcus aureus capability in interfering chemotaxis and impairing function of neutrophils leading to disseminate pathogen extensively in skin tissue layers and failure in the resolution of inflammation (Iwatsuki et al., 2006; Hirsch et al., 2008).
Late stage of inflammation is maintained by more complex interactions among several cell types including lymphocytes and macrophages that dominate the inflammation noticed in oil-treated mice at day 5 after infection (Fig. 3). Tissue macrophages and newly recruited monocytes which differentiate into macrophages, also function in phagocytosis and hence killing of bacteria. In addition, they contribute to tissue repair and act as antigen presenting cells that are required for the induction of specific immune responses (Young et al., 2006). In response to inflammation, infected mice have increased monocyte and lymphocyte counts and these are significantly decreased toward the normal values after treatment with oil, as a result of increased recruitment to infected skin. Similar effect observed in absolute differential counts of eosinophil and basophil, however the standard drug showed significant increase in basophil counts, revealing the occurrence of allergic skin reactions that may be due to repeated treatments of antibiotic (Table 2, Fig. 3) (Abbas and Lichtman, 2011). These results clearly demonstrate that fixed oil constituents act synergistically in reducing inflammation and stimulating immune response in staphylococcal-infected mice. Previous findings displayed that Nigella sativa exerted a stimulatory effect on macrophages through interleukin (IL)-3 which was secreted by T-lymphocytes under the effect of fixed oil (Swamy and Tan, 2000). It also increases the percentages of CD4-positive subsets of T-lymphocytes, which activates macrophages and regulate their function (Szejda et al., 1984). Production of IL-1α in a medium containing fixed oil and macrophages confirmed that the former has a stimulatory effect on macrophage either directly or through IL-3 (Haq et al., 1995). In addition, oil exhibited stimulatory effect on peritoneal macrophages in streptozotocin-induced diabetic hamsters by increasing phagocytic activity either directly or via stimulation of lymphocytes (Fararh et al., 2004). In contrast, treatment with Nigella sativa oil induced a 2-fold decrease in antibody production in response to typhoid vaccination as compared with control rats (Islam et al., 2004). Accordingly, Salem (2005) suggested that crude extracts of Nigella sativa and its active constituent, thymoquinone, may enhance T cell-mediated immune response but suppress humoral immune response, and this required further scientific verification.
The oil healing effect was also marked by the absence of local signs of infection and inflammation, such as necrotic regions and accumulation of edema in infected treated skin (Fig. 2). That may be related to antioxidant effect of the oil and contribution of active leukocytes in tissue repair at the site of infection. The production of free radicals at or around the infected site may delays the healing process through the destruction of lipids, proteins and extracellular matrix elements (Burits and Bucar, 2000). Constituents of the oil, having antioxidant property, are able to inhibit lipid peroxidation and activate antioxidant enzymes, and thus protect tissue impairment and improve healing of infected skin, as confirmed previously (El-Dakhakhny et al., 2002; Hosseinzadeh et al., 2007; Alkharfy et al., 2011).
CONCLUSION
The results of this study revealed that fixed oil of Nigella sativa seeds might has potent bactericidal, anti-inflammatory, immune stimulating and/or antioxidant agents that improve healing of staphylococcal-infected skin in BALB/c mice, by inhibiting pathogenic growth and expansion, reducing inflammation and prevent tissue impairment. These effects provide scientific basis for the use of Nigella sativa in traditional medicine to treat skin infections and inflammations.
ACKNOWLEDGMENTS
The author acknowledge Dar Al Dawa, Na'our, Jordan, for kind contribution in supplying bacterial strain, Wasfi Al-bekearat for his support and technical assistance, Al-al-Bayt University, Department of Biological Sciences, Mafraq, Jordan, Ismail Zayed for the help in photographing histological sections, Yarmouk University, Department of Biological Sciences, Irbid, Jordan and Al-al-Bayt University, Mafraq, Jordan, for providing necessary facilities to conduct this work.
REFERENCES
- Abu-Al-Basal, M.A., 2009. In vitro and In vivo anti-microbial effects of Nigella sativa linn. seed extracts against clinical isolates from skin wound infections. Am. J. Applied Sci., 6: 1440-1447.
Direct Link - Ali, B.H. and G. Blunden, 2003. Pharmacological and toxicological properties of Nigella sativa. Phytother. Res., 17: 299-305.
CrossRefPubMedDirect Link - Amara, A.A., M.H. El-Masry and H.H. Bogdady, 2008. Plant crude extracts could be the solution: Extracts showing in vivo antitumorigenic activity. Pak. J. Pharm. Sci., 21: 159-171.
PubMedDirect Link - Abu-Irmaileh, B.E. and F.U. Afifi, 2003. Herbal medicine in Jordan with special emphasis on commonly used herbs. J. Ethnopharmacol., 89: 193-197.
CrossRefDirect Link - Bourgou, S., M.E. Kchouk, A. Bellila and B. Marzouk, 2010. Effect of salinity on phenolic composition and biological activity of Nigella sativa. Acta Hortic., 853: 57-60.
Direct Link - Burits, M. and F. Bucar, 2000. Antioxidant activity of Nigella sativa essential oil. Phytother. Res., 14: 323-328.
CrossRefPubMedDirect Link - Doeing, D.C., J.L. Borowicz and E.T. Crockett, 2003. Gender dimorphism in differential peripheral leukocytes counts in mice using cardiac, tail, foot and saphanous vein puncture methods. MBC Clin. Pathol., 3: 3-3.
CrossRefDirect Link - El Gazzar, M., R. El Mezayen, J.C. Marecki, M.R. Nicolls, A. Canastar and S.C. Dreskin, 2006. Anti-inflammatory effect of thymoquinone in a mouse model of allergic lung inflammation. Int. Immunopharmacol., 6: 1135-1142.
CrossRefDirect Link - El-Dakhakhny, M., N.J. Madi, N. Lembert and H.P. Ammon, 2002. Nigella sativa oil, nigellone and derived thymoquinone inhibit synthesis of 5-lipoxygenase products in polymorphonuclear leukocytes from rats. J. Ethnopharmacol., 81: 161-164.
Direct Link - Fararh, K.M., Y. Atoji, Y. Shimizu, T. Shiina, H. Nikami and T. Takewaki, 2004. Mechanisms of the hypoglycaemic and immunopotentiating effects of Nigella sativa L. oil in streptozotocin-induced diabetic hamsters. Res. Vet. Sci., 77: 123-129.
PubMed - Francois, P., A. Scherl, D. Hochstrasser and J. Schrenzel, 2010. Proteomic approaches to study Staphylococcus aureus pathogenesis. J. Proteomics, 73: 701-708.
PubMed - Griffith, D.C., L. Harford, R. Williams, V.J. Lee and M.N. Dudley, 2003. In vivo antibacterial activity of RWJ-54428, a new cephalosporin with activity against gram-positive bacteria. Antimicrob. Agents Chemother., 47: 43-47.
PubMed - Hahn, B.L., C.C. Onunkwo, C.J. Watts and P.G. Sohnle, 2009. Systemic dissemination and cutaneous damage in a mouse model of staphylococcal skin infections. Microb. Pathog., 47: 16-23.
PubMed - Hanafy, M.S.M. and M.E. Hatem, 1991. Studies on the antimicrobial activity of Nigella sativa seed (black cumin). J. Ethnopharmacol., 34: 275-278.
CrossRefDirect Link - Hirsch, T., M. Spielmann, B. Zuhaili, T. Koehler and M. Fossum et al., 2008. Enhanced susceptibility to infections in a diabetic wound healing model. BMC Surgery, 8: 5-5.
CrossRef - Hosseinzadeh, H., S. Parvardeh, M.N. Asl, H.R. Sadeghnia and T. Ziaee, 2007. Effect of thymoquinone and Nigella sativa seeds oil on lipid peroxidation level during global cerebral ischemia-reperfusion injury in rat hippocampus. Phytomedicine, 14: 621-627.
CrossRefPubMedDirect Link - Houghton, P.J., R. Zarka, B. de las Heras and J.R.S. Hoult, 1995. Fixed oil of Nigella sativa and derived thymoquinone inhibit eicosanoid generation in leukocytes and membrane lipid peroxidation. Planta Med., 61: 33-36.
CrossRefPubMedDirect Link - Islam, S.N., P. Begum, T. Ahsan, S. Huque and M. Ahsan, 2004. Immunosuppressive and cytotoxic properties of Nigella sativa. Phytother. Res., 18: 395-398.
CrossRef - Kugelberg, E., T. Nostrom, T.K. Petersen, T. Duvold, D.I. Anderson and D. Hughes, 2005. Establishment of a superficial skin infection model in mice by using Staphylococcus aureus and Streptococcus pyogenes. Antimicrob. Agents Chemother., 49: 3445-3451.
Direct Link - Kuo, C.F., C.C. Chen, Y.H. Luo, R.Y. Huang, W.J. Chuang, C.C. Sheu and Y.S. Lin, 2005. Cordyceps sinensis mycelium protects mice from group a streptococcal infection. J. Med. Microb., 54: 795-802.
CrossRef - Lev, E. and Z. Amar, 2002. Ethnopharmacological survey of traditional drugs sold in the Kingdom of Jordan. J. Ethnopharmacol., 82: 131-145.
CrossRefDirect Link - Mashhadian, N.V. and H. Rakhshandeh, 2005. Anti-bacterial and anti-fungal effects of Nigella sativa extracts against S.aureus, P. aeruginosa and C. albicans. Pak. J. Med. Sci., 21: 47-52.
Direct Link - Molne, L., M. Verdrengh and A. Tarkowski, 2000. Role of neutrophil leukocytes in cutaneous infection caused by Staphylococcus aureus. Infect. Immun., 68: 6162-6167.
Direct Link - Nickavar, B., F. Mojab, K. Javidnia and M.A.R. Amoli, 2003. Chemical composition of the fixed and volatile oils of Nigella sativa L. from Iran. Zeitschrift Naturforschung C, 58: 629-631.
CrossRefDirect Link - Said, O., K. Khalil, S. Fulder and H. Azaizeh, 2002. Ethnopharmacological survey of medicinal herbs in Israel, the Golan Heights and the West Bank region. J. Ethnopharmacol., 83: 251-265.
CrossRefDirect Link - Salem, M.L., 2005. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int. Immunopharmacol., 5: 1749-1770.
CrossRefPubMedDirect Link - Salman, M.T., R.A. Khan and I. Shukla, 2008. Antimicrobial activity of Nigella sativa Linn. seed oilagainst multi-drug resistant bacteria from clinical isolates. Nat. Prod. Radiance, 7: 10-14.
Direct Link - Swamy, S.M.K. and B.K.H. Tan, 2000. Cytotoxic and immunopotentiating effects of ethanolic extract of Nigella sativa L. seeds. J. Ethnopharmacol., 70: 1-7.
CrossRefDirect Link - Szejda, P., J.W. Parce, M.S. Seeds and D.A. Bass, 1984. Flow cytometric quantitation of oxidative product formation by polymorphonuclear leukocytes during phagocytosis. J. Immunol., 133: 3303-3307.
PubMed - Thormar, H. and H. Hilmarsson, 2007. The role of microbicidal lipids in host defense against pathogens and their potential as therapeutic agents. Chem. Phys. Lipids, 150: 1-11.
CrossRef - Von Vietinghoff, S. and K. Ley, 2008. Homeostatic regulation of blood neutrophil counts. J. Immunol., 181: 5183-5188.
PubMed