
Z. Esfahanibolandbalaie
Biotechnology Center, Iranian Research Organization for Science and Technology, No. 71, Forsat Street, Enghelab Ave., P.O. Box 15815-3538, Tehran 15819, Iran
K. Rostami
Biotechnology Center, Iranian Research Organization for Science and Technology, No. 71, Forsat Street, Enghelab Ave., P.O. Box 15815-3538, Tehran 15819, Iran
S. S. Mirdamadi
Biotechnology Center, Iranian Research Organization for Science and Technology, No. 71, Forsat Street, Enghelab Ave., P.O. Box 15815-3538, Tehran 15819, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 22 | Page No.: 2553-2559







ABSTRACT
The extracellular α-amylase production by Aspergillus oryzae was studied in submerged fermentation using an Adlof-Kuhner orbital shaker. The effect of initial pH values in the range of 4 to 7.5 on enzyme production was investigated and initial pH medium of 6.2±0.1 resulted in enhanced α-amylase production. The effect of carbon and nitrogen source and composition was examined and it has been observed that corn starch concentration of 15 g L-1 has sound effect on enzyme production. The medium containing corn starch, sodium nitrate resulted in considerable higher enzyme production. Further, the yeast extract of 2.5 g L-1 in the medium produced higher enzyme in view to other organic nitrogen sources. The effect of temperature on α-amylase production from 20 to 40°C has been studied and at 35±1°C higher α-amylase has been obtained. The effect of shaker’s speed on α-amylase production from 50 to 200 rpm was investigated. And at about 180 rpm higher enzyme production has been observed. In the present study, it has been found that glucose has repressing effect on α-amylase production using A. oryzae PTCC5164.
PDF Abstract XML References Citation
How to cite this article
Z. Esfahanibolandbalaie, K. Rostami and S. S. Mirdamadi, 2008. Some Studies of α-Amylase Production Using Aspergillus oryzae. Pakistan Journal of Biological Sciences, 11: 2553-2559.
DOI: 10.3923/pjbs.2008.2553.2559
URL: https://scialert.net/abstract/?doi=pjbs.2008.2553.2559
DOI: 10.3923/pjbs.2008.2553.2559
URL: https://scialert.net/abstract/?doi=pjbs.2008.2553.2559
INTRODUCTION
The amylase group of enzymes is of grate significance because of its wide range of industrial and domestic application. Nevertheless, the first enzyme produced industrially was an amylase from a fungus in 1894 (Crueger and Crueger, 1984). However, these fungi are widely available, almost harmless and secret a number of extracellular enzymes in large quantities. A. oryzae has been used in Far East Asian food preparation for thousands of years and to produce α-amylase industrially (Morkeberg et al., 1995). α-amylase (E.C. 3.2.1.1: endo-1,4 α-D-glucan gluco-hydrolase) are produced by eukaryotes (fungi including animal and plant) and prokaryotes, which randomly cleave the 1,4-α-D-glucosidic linkage between adjacent glucose units in the linear amylose chain. It has been well documented that the α-amylase production is secreted as a primary metabolite and its production is growth associated (Spohr et al., 1998; Sudo et al., 1994). Alpha-amylase is produced by submerged (Egas et al., 1998; Aguilar et al., 2000) or solid-state fermentation (Viswanthan and Surlikar, 2001). Amylases have extensive applications in the food, fermentation, textile, paper, pharmaceutical and fine chemicals sectors (Pandey et al., 2000; Rosell et al., 2001). Several thousand tones of commercial enzymes are produced from microbial source and a few are extracted from plant and animal tissues. Among which 11, 12 and 14% have been used in textile, starch and dairy sectors (Fogarty and Kelly, 1980), respectively. Further, with the advent of new frontiers in biotechnology the range of α-amylase application has opened-up into many challenging areas such as medical, clinical and analytical chemistries. There are several processes in the clinical and medicinal fields that amylases are used (Witezak, 1999; Lepp et al., 1996). α-amylase has been used for serum evaluation, a liquid-stable reagent containing α-amylase for the new Olympus AU 600 Chemistry, a-revised analyzer, a revised amylase method for the Du Pont Dimension and Ciba-Corning Express clinical chemistry system (Chiu and Chandler, 1995). However, production of extracellular α-amylase employing microorganisms and submerged fermentation has favorable advantages, such as: reliable scale up, simple process control, reasonably high yield and easily recoverable by a series of simple fractionation and purification steps. There is always a tendency to look for a new strain in order to remain in competitive market which can utilize low priced agrochemical side product like molasses as carbon source to produce enzyme. Microorganisms used for production of industrial scale of enzymes such as amylase and protease usually consume complex agro-carbon source such as molasses, wastewater of potato, glucose, starch and pulp and paper. Nitrogen and carbon sources in the medium have strong effect on α-amylase production. The nitrogen is metabolized to produce nucleic acids, primarily amino acids, proteins and cellular components. Production of bulk enzymes over a longer period requires maintenance of high cell density under conditions that promote formation of desirable enzymes while holding the cell growth minimal. Glucose has acted as catabolic repressor for α-amylase production using certain fungi (Morkeberg et al., 1995). Kundu et al. (1973), have reported that nitrogen source such as sodium nitrate could be beneficial for α-amylase production. Production of various enzyme especially amylase, amyloglucosidase and related glucanases by fungi appears to be promising. Therefore in the present study, Aspergillus oryzae PTCC 5164 was used to examine its production capacity using submerged technique. Corn starch and yeast extract was used as carbon and nitrogen source, respectively. To have uniform size pellets, affects of inoculum size and shaker’s speed were in investigated within 48 h. Rate of reaction and denaturation of fungi producing enzyme depend on medium temperature according to Arrhenius equation, therefore it was studied in the range of 20-40°C using Adlof-Kuhner rotary orbital shaker. Effect of initial pH as an important parameter on the enzyme production was examined in the range of 4 to 7.5 and the optimized pH condition for enzyme production has been found at about 6.2±0.1.
MATERIALS AND METHODS
Microorganism preparation: Cultures of A. oryzae PTCC 5164 were maintained on Potato Dextrose Agar (PDA) slants at 4°C. Preculture were routinely prepared every one month. Prior to the experiments the organism was subcultured on the same medium, spore suspension of 1-2 weeks old was used as the inoculum. The fungal spores were grown for 2 days in Erlenmeyer containing 100 mL potato dextrose broth and an inoculum of 3x105 spores mL-1 was used. All the culture experiments were performed in 500 mL Erlenmeyer flasks containing 100 mL of production medium. The Erlenmeyer flasks containing required amount of medium was inoculated under sterile conditions and were stoppered with cotton bungs and covered with aluminum seals. A typical medium as given by Sudo et al. (1993) containing; MgSO4.7H2O 0.5, CaCl2 0.1, KH2PO4 1.00, FeSO4.7H2O, 0.01, 20 and 10 g of starch and nitrogen source were dissolved in l L of distilled water, respectively. Effect of NaNO3, corn-starch, yeast extract, peptone, corn steep liquor, glucose, lactose and maltose on the enzyme production was studied. Glucose, lactose, maltose, corn steep liquor and starch were separately sterilized for 10 min at 100°C and under aseptic condition were added. The initial pH of the medium was varied from 4 to 7.5 in the process, where fermentation temperature was maintained at 35±1°C. The medium pH was adjusted to designed values with diluted HCl and NaOH. About 0.075% v/v Tween 80 was added to the mixture and cells were pelleted by centrifugation (10000 rpm for 20 min). The supernatants were retained for enzyme assay.
Enzyme assay: α-amylase activity was measured according to the method of Bernfeld using 3,5-dinitrosaliculic acid (Bernfeld, 1955) and A540 was measured using Unicom 8620 UV/VIS spectrophotometer. One unit of α-amylase activity has been defined as the amount of enzyme that produced reducing sugar equivalent to 1 μmole of maltose per minute under the assay conditions. A calibration chart using maltose was constructed and used throughout the experiments.
Protein estimation: Protein concentration of the enzyme sample was analyzed employing Bradford method (Bradford, 1976). Bovine Serum Albumin (BSA) was used as standard. A calibration chart was constructed and used throughout the present study.
Reproducibility: All the experiments were carried out in triplicate and the average values have been reported. Values plotted are Mean±SD deviations of three replicates.
RESULTS
Effect of pH: pH is a key parameter in most biological processes and affect the pellets formation, the growth of biomass, synthesis of extracellular α-amylase, it’s secretion and the stability of the amylolytic system (Fogarty, 1983). The effect of initial pH on α-amylase production was investigated in the range of 4 to 7.5, where, 0.1 M NaOH and 0.1 M hydrochloric acid was used to adjust the medium pH, Nahas and Waldemarin (2002) and Morkeberg et al. (1995) have used the approach. The fermentation was carried out at 35±1°C for 72 h. As the medium initial pH was increased from 4 to 5 has shown meager effect on the enzyme production, however increasing it from 5 to 6 it has considerably affected on α-amylase production. Further, increasing the initial pH from 6.7 to 7.5 it has shown adverse affect on enzyme production. It has been presented in Fig. 1 performing the study with medium of initial pH of 6.2±0.1 higher amount of α-amylase was has been produced.
Effect of carbon: The effect of various carbon sources of 15 g L-1 concentration on α-amylase production was investigated. It has been observed that the effect of soluble starch, corn-starch and maltose on enzyme production have approximately the same edge (Fig. 2).
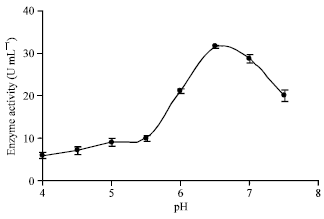 | |
| Fig. 1: | Effect of initial pH on enzyme production |
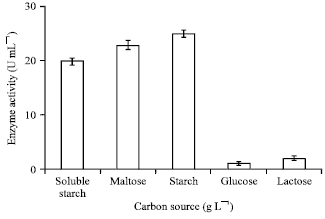 | |
| Fig. 2: | Effect of different carbon source on enzyme production |
Further, glucose and lactose was used as carbon sources and have shown narrow effect on enzyme production. The effect of glucose concentration on enzyme production was verified from 1 to 10 g L-1, on the contrary to biomass vegetation the enzyme production in the medium was negligible. The effect of lactose concentration from 5 to 15 g L-1 on α-amylase production was also investigated and it has been found that the biomass growth and enzyme production was negligible in using lactose (result not shown here). The effect of maltose, soluble starch and corn-starch on enzyme production was investigated from 5 to 30 g L-1, which has been shown in Fig. 3. It can be seen that as the concentration of corn-starch was increased from 5 to 15 g L-1 the α-amylase production increased. Increasing corn starch concentration more than 15 g L-1 has decreased enzyme production. Further, corn-starch concentration of about 15 g L-1 has contributed in higher α-amylase production. Furthermore, as the concentration of maltose was increased from 5 to 15 g L-1 which had a conservative effect on α-amylase production and further increasing it’s concentration from 15-30 g L-1,had inversed effect on enzyme production. Soluble starch of 5 to 15 g L-1 has shown substantial affect on α-amylase production.
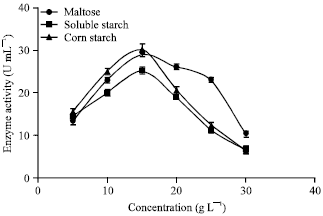 | |
| Fig. 3: | Effect of soluble starch, corn starch and maltose on enzyme production |
 | |
| Fig. 4: | Effect of nitrogen source on enzyme production |
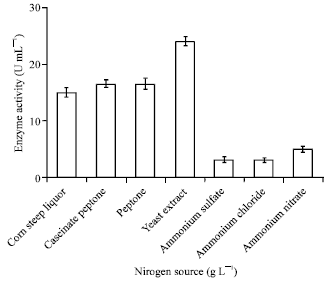
Effect of nitrogen: The effect of inorganic nitrogen sources such as ammonium nitrate, sulfate, chloride and sodium nitrate on α-amylase production was studied. Further, the effect of organic nitrogen source like powdered Corn Steep Liquor (CSL) having Dry Solid (DS) of 9.6 g L-1, yeast extract, peptone caseinate and peptone on α-amylase production was examined (Fig. 4).
Effect of inorganic nitrogen: Ammonia is the major regulator of nitrogen assimilation and in its presence the utilization of other nitrogen sources such as nitrate, amino acids and proteins is repressed. The effect of various nitrogen sources such as ammonium chloride, nitrate and sulfate of 10 g L-1, as described by Sudo et al. (1993) and using corn starch as carbon source on α-amylase production was studied.
 | |
| Fig. 5: | Effect of sodium nitrate on enzyme production |
 | |
| Fig. 6: | Effect of organic nitrogen on enzyme production |
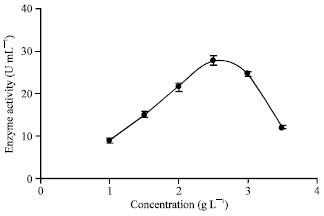
Using ammonium chloride as a nitrogen source during fermentation pH of the medium decreased rapidly and inconsiderable amount of the enzyme has been produced. The effect of ammonium nitrate and sulfate of the same w/v (%) on α-amylase production has shown approximately similar result like ammonium chloride, considering higher edge of ammonium nitrate and the result has been presented in Fig. 4. However, ammonium nitrate has influenced the enzyme production relatively inconsiderable. The effect of sodium nitrate concentration on α-amylase production was also investigated. It has been presented in Fig. 5 that by increasing the sodium nitrate concentration from 0.5 to 2.5 g L-1 α-amylase production has increased. Further, increasing the sodium nitrate concentration from 2.5 to 3.5 it has adverse affect on the enzyme production.
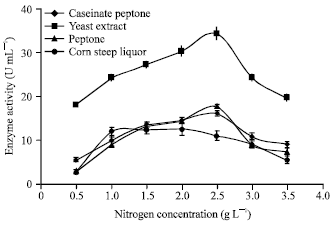
Effect of organic nitrogen: It has been presented in Fig. 6 that as the yeast extract concentration was increased from 0.5 to 2 g L-1 the enzyme production has increased. Further, higher amount of α-amylase has been produced at about 2.5 g L-1 yeast extract. Sound effect of yeast extract on α-amylase production might be due to presence of vitamin B group (promoting growth), free amino acids and carbohydrate.
 | |
| Fig. 7: | Effect of inoculum concentration on enzyme production |
The effect of peptone and peptone caseinate on α-amylase production was examined in the range of 0.5 to 3.5 g L-1. The result been presented in Fig. 6. The affect of peptone and peptone caseinate concentration in the range of 1.0 to 2.5 g L-1 on α-amylase production are marginal, except a peak at about 2.5 g L-1. Further enhancement in the concentration of the peptone and peptone caseinate in the batch system had shown adverse affect on enzyme production. Furthermore, it can be observed that as the concentration of corn steep liquor was increased from 0.5-2.5 g L-1 a conservative affect on α-amylase production has been achieved. Moreover, as the corn steep powder concentration was increased from 0.5 to about 1.0 g L-1 had meager effect on the α-amylase production comparing with other three types of nitrogen sources.
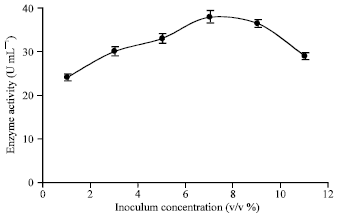
Effect of inoculum size: The medium was inoculated with preculture media in the range of 1-11 v/v (%) having spore concentration of 3x105 mL-1. The counting was performed using the Neubar lamel method (Florence et al., 1992). Preculture were subcultured on the Potato Dextrose Agar (PDA) medium, which was maintained at 4°C and 2-3 weeks old inoculums was used. The preculture were routinely prepared every 2 months. It could be seen as the inoculum’s size was increased from 4 to 6.5 v/v (%) α-amylase production has increased (Fig. 7). Further, as the inoculum size was increased from 6.5 to 7.5 v/v (%) higher amylase production has been produced. It has been depicted in Fig. 7 that by increasing inoculum’s concentration higher than 8 v/v(%) about 9 the enzyme production has ceased gradually. Furthermore, increasing of inoculum addition more than 9.5 has shown adverse effect ceasing α-amylase production. Therefore, inoculum size of 7 v/v (%) was used throughout the present study.
 | |
| Fig. 8: | Effect of shaker’s speed on enzyme production |
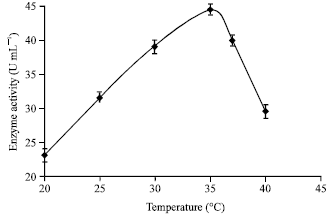 | |
| Fig. 9: | Effect of temperature on enzyme production |
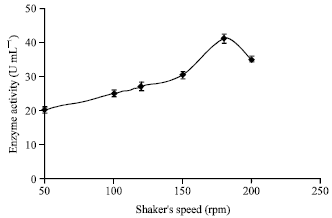
Effect of shaker’s speed: The effect of shaker’s speed on secretion of α-amylase was investigated in the range of 50 to 200 rev min-1. An Adolf-Kuhner orbital shaker was used to cultivate the medium for α-amylase production. It has been presented in Fig. 8, that as the shaker’s speed was increased from 50 to 180 rev min-1, the enzyme production has increased. Further, increasing in shaker’s speed has shown no sound effect on the enzyme production. Therefore, the optimal range of shaker’s speed on pellets size and α-amylase production has been found in the present investigation to be about 180 rpm.
Effect of temperature: The effect of temperature on α-amylase production in the range of 20 to 40°C was examined. It can be observed that as the medium temperature was increased from 20 to 35°C the enzyme production has increased and at about 35±1°C higher enzyme production was achieved (Fig. 9). Further, it has been observed that by increasing the temperature of the fermentation system above 35°C had adverse effect on the enzyme production using A. oryzae in the present study. Furthermore, as the temperature was decreased below 20°C very low enzyme activity has been detected.
DISCUSSION
pH of the medium has governing role on pellets size, growth of fungi, production and stability of amylolytic enzyme. At initial pH of 6 and 7±0.1 pellets were more approximately spherical in the medium (visual observation) and at about 6.2±0.1 more enzymes production has been detected. Therefore, it can be concluded that initial pH of 6.2±0.1 has considerable influence on the enzyme production, which shows coherence with optimum activity of α-amylase by 1 value. The carbon sources and compositions have key influence on α-amylase production. Therefore, it was desirable to utilize relatively cheap economical source of carbon such as molasses for the enzyme production. However, due to inconsistence results obtained in the course of fermentation its consumption was omitted in the future studies. The present result may be due to incomplete consumption of molasses or presence of undesirable species like heavy metals or pesticide etc. in the medium. Molasses normally contain mud, sodium chloride, calcium carbonate, gum material, heavy metals, pesticide and undesirable microbial contamination which may have sever effect on the enzyme production. Therefore, effect of carbon material such as: starch, corn starch, glucose, maltose and lactose on the enzyme production were investigated. Effect of glucose concentration on the enzyme production was studied and on the contrary to remarkable biomass production, negligible enzyme activity has been detected. Therefore, it has been realized that glucose has acted as catabolic repressor for the enzyme production using the present Aspergillus oryzae strain. The present result corroborates the report of Djekrif-Dakhmouche et al. (2006) and contradicts the result of Spohr et al. (1998). Effect of soluble starch, corn starch and maltose in the range of 5 to 30 g L-1 on the enzyme production was performed and concentration of 15 w/v (%) in the medium resulted in higher α-amylase production. Further, it has been observed that maltose has conservative effect on α-amylase production in comparison to corn starch. Considerable effect of corn starch on α-amylase production in comparison to soluble starch, which means it could be used in large scale operation as more economical source. The nitrogen is metabolized to produce primarily amino acids, nucleic acid, proteins and cellular component in fungi. Production of α-amylase is affected by selected and low nitrogen concentration; therefore it is required to pin point proper type and percentage of necessary nitrogen compound. In the course of examining the effect of ammonium salts such as ammonium chloride, nitrate and sulfate on α-amylase production it has been realized that ammonium chloride and sulfate have meagerly affected α-amylase production, which may be due to changing the production environment to more acidic. Further, it was observed that ammonium nitrate has not served any significant changes on the result of enzyme production comparatively. It may be because as the ammonium ion is taken-up the ionic balance of the cell is maintained by the extrusion of hydrogen ions. This phenomenon can result in the formation of more acidic environment unsuitable for the fungi species used; however its effect depends on the co-ionic species. The sound effect of sodium nitrate in the range of 2-2.5 g L-1 in the medium may be on the count of maintaining low biomass vegetation in the process, which is desirable as process parameter employing A. oryzae for α-amylase production. Furthermore, as the sodium nitrate concentration was increased above more than 2.5 g L-1 the enzyme production has decreased, since sodium nitrate at high concentration in the medium could enter the cell by diffusing down a gradient created by its rapid metabolism within the cell. However, at low concentration it is probable that active transport takes place. Effect of organic nitrogen source such as yeast extract, corn steep liquor, peptone and peptone caseinate on α-amylase production has been shown in Fig. 6. The considerable influence of yeast extract concentration on α-amylase production may be due to presence of vitamin B group (promoting growth), free amino acids and carbohydrate. However at higher concentration of yeast extract say about 2.5 g L-1 the enzyme production has decreased, which may be due to changes has occurred in C/N ratio and a more detailed comprehensive study is recommended. A combination of yeast extract and specified amount of sodium nitrate while maintaining certain C/N ratio resulted in considerable amount of α-amylase production (result not shown here). The marginal effect of peptone and peptone caseinate on α-amylase production may be due to presence of free ammonium in the medium. Further, as the concentration of corn steep liquor was increased from 0.5-2.5 g L-1 has shown adverse effect on the enzyme production, which may be due to presence of glucose in the composition. However, a detailed study is required to investigate the effect of corn steep liquor on α-amylase production, since it may be used as economical nitrogen raw material source. Effect of inoculum size on fungi growth form in submerged culture is of considerable importance in fermentation technology as it influences, growth rate, aeration, power consumption for stirring and the ease with which biomass can be separated from the broth. In addition inoculum size is one of the factor influencing the growth form (filamentous or pellet). The medium was inoculated in the range of 1-11%v/v containing spore concentration of 3x105 mL-1 this was performed because pellets of compact and spherical size is preferred during α-amylase production. Moreover, in order to maintain balanced substrate composition in the fermentation process for the enzyme production. However, it has been observed that as the inoculum size was increased from 1-6.5 and further to 7.5 v/v (%) in the medium higher α-amylase has been produced. This is because at lower inoculum; fungi have to undergo extensive growth and branching before nutrient utilization, resulting in conservative production of α-amylase. On the other hand, when high inoculum concentration was used from 9-11 v/v (%) has resulted in irregular shape, big size pellets (visual observation) and might have consumed major amount of substrate at initial stage of fermentation. Depriving nutrient availability in the latter part of log phase which has resulted in adverse effect on α-amylase production. Therefore, in studies a head 7 v/v (%) inoculum was used. Effect of shaker speed on fungal medium is of considerable important since; (1) oxygen has a low solubility in water and hence diffuses slowly into static medium (2) oxygen solubility in fermentation medium decreases since carbon, nitrogen, minerals and vitamin etc. act as impurities, in addition the fermentation temperature during fungi cultivation is above 30°C. Some fungi are exclusively oxidative in their metabolism and are unable to survive under poor aeration, or completely anaerobic. The metabolic activity of fungi produces a variety of gaseous or volatile compounds such as carbon dioxide which is highly soluble in water. Therefore, aeration and agitation sweep away undesirable gases such as carbon dioxide (CO2) and provide oxygen for metabolic activity and maintain homogenous medium. However, a considerable attention is required to avoid oxygen toxicity. We have observed that as shaker’s speed was increased from 50 to 180 rpm the enzyme production has enhanced. However, increasing the shaker’s speed to 200 rpm has resulted in production of pellets of very small size (visual observation) and toxic the fungi by providing excess oxygen in the medium resulting in adverse effect on α-amylase production. Therefore, shaker’s speed of 180 rpm was used in further studies as potential parameter. Temperature of fermentation medium has strong influence on solubility of desirable and undesirable gases such as oxygen, carbon dioxide, nutrient species, rehelogy and reaction rate. According to Arrhenius equation increasing in temperature 8-10°C doubles the rate of reaction. This is because at low medium temperature, say, 20°C rate of reaction of α-amylase production is slow. The present study reported that by increasing the medium temperature from 20 to 35±1°C higher amount of enzyme has been produced. Further, at reasonably high temperature, about 40°C the enzyme production has ceased, which may be due to a type of malfunctioning similar to denaturing taking place in the microorganism. Therefore, conducting the fermentation process at about 35±1°C leads to higher enzyme production using the present species of Aspergillus oryzae. It is desirable to address the effect of corn starch on the enzyme production in fed-batch mode of operation, as the initial concentration of carbon has determined role on carbon balance (C/N) in the production process. Further, it is recommended a detailed study on exploiting molasses (beet root and sugar cane) and corn steep liquor as carbon and nitrogen sources, respectively.
ACKNOWLEDGMENT
The authors appreciate the support provided by Biotechnology Center of Iranian Research Organization for Science and Technology (IROST), Tehran, Iran.
REFERENCES
- Aguilar, G., J. Morlon-Guyot, B. Trejo-Aguilar and J.P. Guyot, 2002. Purification and characterization of an extra cellular alpha amylase produced by Lactobacillus manihotivorans LMG 1801 (T), an amylolytique lactic acid bacterium. Enzyme Microb. Technol., 27: 406-413.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Djekrif-Dakhmouche, S., Z. Gherib-Aoulmi, Z. Meraihi and L. Bennamoun, 2006. Application of a statistical design to the optimization of culture medium for the α-amylase production by Aspergillus niger ATCC 16404 grown on orange waste powder. J. Food. Eng., 73: 190-197.
CrossRefDirect Link - Morkeberg, R., M. Carlsen and J. Nielsen, 1995. Induction and repression of α-amylase production in batch and continuous cultures of Aspergillus oryzae. Microbiology, 141: 2449-2454.
CrossRefDirect Link - Nahas, E. and M.M. Waldemarin, 2002. Control of amylase production and growth characteristics of Aspergillus ochraceus. Rev. Latinoam. Microbiol., 44: 5-10.
PubMedDirect Link - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Rosell, C.M., M. Haros, C. Escriva and C.B. de Barber, 2001. Experimental approach to optimize the use of alpha amylases in bread making. J. Agric. Food Chem., 49: 2973-2977.
CrossRefDirect Link - Spohr, A., M. Carlsen, J. Nielsen and J. Villadsen, 1998. Amylase production in recombinant Aspergillus oryzae during fed-batch and continues cultivation. J. Ferment. Bioeng., 86: 49-56.
CrossRefDirect Link - Sudo, S., T. Ishikawa, Y. Takayasu-Sakamoto, K. Sato and T. Oba, 1993. Characteristics of acid-stable -amylase production by submerged culture of Aspergillus kawachini. J. Ferment. Bioeng., 76: 105-110.
CrossRefDirect Link - Sudo, S., T. Ishikawa, K. Sato and T. Oba, 1994. Compression of acid stable amylase production by Aspergillus kawachini: Solid-state and submerged cultures. J. Ferment. Bioeng., 77: 483-489.
CrossRefDirect Link - Witezak, Z., 1999. Thio-sugars: Biological relevance as potential new therapeutics. Current. Med. Chem., 6: 165-178.
PubMed - Egas, M.C., M.S. da Costa, D.A. Cowan and E.M. Pires, 1998. Extracellular α-amylase from Thermus filiformis Ork A2: Purification and biochemical characterization. Extremophiles, 2: 23-32.
CrossRef