
P. Thobunluepop
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Talard Sub-District, Muang District, Maha Sarakham 44000, Thailand
E. Pawelzik
Institute of Agricultural Chemistry, Section of Quality of Plant Products, Georg-August University of Goettingen, Carl-Sprengel-Weg , D-37075, Goettingen, Germany
S. Vearasilp
Department of Agronomy, Faculty of Agriculture, Chiang Mai University, 50000, Thailand
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 19 | Page No.: 2291-2299







ABSTRACT
This study aimed to evaluate the perspective changes of several physiological performances of rice seeds cv. KDML 105 which were coated with various seed coating substances [chemical fungicide, captan (CA) and biological coating polymers; chitosan-lignosulphonate polymer (CL) and eugenol incorporated into chitosan-lignosulphonate polymer (E+CL)] during storage (12 months). CA significantly increased seed moisture content and seed water activity through out the storage period. The qualities and viability of the seeds were seriously declined by this treatment. Moreover, CA inhibited the shoot and root development, seedling dry weight accumulation, delayed the seed germination and seedling growth rate. CA treated seeds were susceptible to stress conditions that declined the seed germination potential under cold, high moisture and temperature stress conditions. Nevertheless, CL and E+CL coating polymer could maintain seed storability, which significantly improved seed germination and seedling performances. These improvements were attributed to maintain the nutritive reserve and dehydrogenase activity in seeds. Moreover, the biological seed treatment stimulated the embryo growth and so speeding up the seedling emergence when compared untreated seeds.
PDF Abstract XML References Citation
How to cite this article
P. Thobunluepop, E. Pawelzik and S. Vearasilp, 2008. The Perspective Effects of Various Seed Coating Substances on Rice Seed Variety Khao Dawk Mali 105 Storability I: The Case Study of Physiological Properties. Pakistan Journal of Biological Sciences, 11: 2291-2299.
DOI: 10.3923/pjbs.2008.2291.2299
URL: https://scialert.net/abstract/?doi=pjbs.2008.2291.2299
DOI: 10.3923/pjbs.2008.2291.2299
URL: https://scialert.net/abstract/?doi=pjbs.2008.2291.2299
INTRODUCTION
Nowadays, synthetic fungicides is traditionally used seed treatment to protect seed- and soil-borne fungi immediately after planting (Neergaard, 1997). However, they may produce phytotoxicity effects in the seeds that mignht cause losses of germinability and seedling vigor. Von Pine et al. (1995) found that captan and pirimifos reduced maize seed storability, which influenced the following behavior under field conditions. Silva et al. (1996) also reported clorpirifos and captan reduced the germination and vigor of maize seeds during storage. Therefore, the alternative ways to replace chemical fungicides application that receive increasingly attention were mechanical methods, resistant plant varieties, improved cultural practices and botanical fungicides. Currently, increasing academic interest has emerged on the botanical fungicides as an alternative to synthetic fungicides, which might have the potential to minimize undesirable side effects. The use of medical plant extracts as pesticides is a promising way in a chemical-free agricultural system (Anonymus, 2000). Eugenol and eugenol acetate are active compounds contained in clove. They act simultaneously as bactericide, fungicide and virucide (Thobunluepop et al., 2008). Additionally, these extracts are affordable and can easily be obtained from readily available plant (Thobunluepop et al., 2007). However, some essential oils are able to produce phytotoxic effects on seed viability when used at high concentrations. Tworkoski (2002) reported that essential oils; red thyme, summer savory, cinnamon and clove (5-10%v/v) were at most phytotoxic and caused electrolyte leakage resulting in cell death of rice seed.
According to Yildirim and Hoy (2003), coating seeds with chemicals or alternative substances could reduce the percentage of plant damage by pest. They found that when comparing with direct drench application methods, the seed coating technology significantly decreased the percentage of plant damage at 85% of the level of pesticide utilization. In the past decade, the seed coating technology has been continuously improved, particularly in European countries and North America (Struszczyk and Pospieszny, 1997). The new techniques often require improving application systems for better establish dosages and coverage of materials. The coating material is thin and water-permeable. It can be mixed with natural products or other compounds, which have protective effects on seeds. Seed coating techniques have been used to reduce the seedling emergence time, produced synchronized emergence, improved emergence rate and increased seedling stand production in many field crops like wheat (Basra et al., 2005) and rice (Farooq et al., 2004).
Seed quality has pronounced effects of progeny survival and fitness, especially under storage conditions and stressful conditions (Agrawal, 2002). The morphological and chemical attributes of these seed components are critical for successful seedling establishment under various storage or stress conditions. Moreover, seedlings development depends on seed reserves in the early stages of development. This is particularly relevant under storage conditions. For example, cotyledons that emerge before the canopy closes in spring are exposed to fungicides levels that are capable of damaging plant DNA and photosynthetic machinery, which is the cause of abnormal seedling (Berjak and Villiers, 1972). Thus, the present study aimed to investigate the perspective effects of various seed coating substances on rice seed cv. KDML 105 storability.
MATERIALS AND METHODS
In 2007, the study was conducted at the Seed Science and Technology Laboratory, Faculty of Agriculture, Department of Agronomy, Chiang Mai University, Thailand. Dry graded rice seeds (Oryza sativa L. cv. KDML 105) from one seed lot were supplied by Bureau of Seed Multiplication of Thailand. The initial seed moisture content (MC) and germination percentage were 10.65 and 96.00%. The split-plot design with four replications was applied. The main plot was various seed coating substances [chemical fungicide; captan (CA) and biological coating polymers; chitosan-lignosulphonate polymer (CL) and eugenol incorporated into chitosan-lignosulphonate polymer (E+CL)] and untreated (control) seeds. The sub-plot was the storage duration (12 months). The seeds of each treatment were randomly stored in plastic bag sealed in an incubation chamber (KPB6395FL, Termaks, S/N 2-858 Germany), at controlled temperature of 30±2°C and relative humidity at 40±5%. Seeds were sampled immediately and then collected each month for the determination of seed qualities and the seedling performances.
The preparation of traditional captan treatment (CA): Captan fungicide (1, 2, 3 and 6-tetrahydro-N-(trichloromethyl thio) phthalimide was applied as slurry dust in a solution of polyethylene glycol (PEG 8000 at -2 MPa) (4 g of captan per 1 kg of the seeds). After that, seeds were dried to 10±2% of MC at 35°C to obtain a similar MC as the control (untreated) seeds.
The preparation of chitosan-lignosulphonate coating polymer (CL): Three percentage of chitosan-lignosulphonate coating polymer was prepared by adding 3 g chitosan (Biolife®ELAND CROP, Thailand) into 100 mL of 1% (v/v) acetic acid combined with 1% (w/v) sodium-lignosulphonate (Fluka, Germany) in distilled water. Then, 10 mL of chitosan-lignosulphonate polymer was sprayed and mixed well into 500 g seeds. The MC of 10±2% was obtained after drieing at 35°C.
The preparation of Eugenol incorporated into chitosan-lignosulphonate coating polymer (E+CL): One percentage of eugenol incorporated into chitosan-lignosulphonate coating polymer was prepared by adding 0.5 mL eugenol solution (Fluka, Germany) into 50 mL of 3% chitosan-lignosulphonate coating polymer binder. Then, it was sprayed onto mixed-well 500 g seeds samples. The seeds were dried at 35°C to obtain MC of 10±2%.
Seeds quality assessment: The seeds moisture content (MC) was determined by hot air oven method and calculated on a wet mass basis (ISTA, 2006). Testo 650 apparatus was used for analyzing the seed water activity (Aw). Seed viability was measured as percentage of normal seedlings by standard germination test (Germ) at 28°C for 14 days (ISTA, 2006). Dehydrogenase activity of seeds was analyzed by Tetrazolium Test (TZ) according to ISTA (2006): fifty seeds were cut longitudinally through the embryo and soaked in 0.20% (w/v) of 2, 3 and 5-triphenyltetrazolium chloride solution for 24 h at 2°C in the darkness, then scored according to intensity and location of red stain on seed.
Seed vigor was estimated by assessing the following parameters: Shoot (Sh) and root (Ro) lengths of 20 normal seedlings grown in moist towel paper kept at optimum temperature (28°C) were measured in centimeter (cm) on the day of the final count (14 days) (ISTA, 2006).
The electrical conductivity test (Cond) was used to determine seed membrane integrity by measuring the seed leakage. This test followed procedures described by ISTA (2006); 100 seeds were weighted and soaked into de-ionized water. Then, they were placed at 25°C for 24 h. Finally, the solution was determined for electrical conductivity by the conductivity meter (Sartorius, model PP-20).
Seedling Growth Rate (SGR) to assess the amount of dry seed weight that was respired for producing 1 g of dry root and shoot was metabolic efficiency of the seed. Thus, higher the value of SGR, which meant the efficiency of seed as more seed reserve are mobilized for producing root and shoot (ISTA, 2006). The amount of SGR was calculated as:
| SGR | = | SDW – (SHW + RTW + RSW) |
Where:
| SDW | = | Dry weight of seed before germination |
| SHW | = | Dry weight of shoot |
| RTW | = | Dry weight of root |
| RSW | = | Dry weight of seed after germination |
Seedlings dry weight (SDW) determining by the dry weights (milligram, mg) of seedlings was determined on the 14th day after germination. After that, seedlings were oven dried at 80°C for 24 h (ISTA, 2006).
Germination Index (GI) is speed of germination as an indication of vigorous seed lots. The number of germinating seeds was counted every day from the first day (14 days). The cumulative index was calculated by the following formula (ISTA, 2006):
GI = (n1/1 + n2/2 + n3/3 + ...+ nx/x) |
where, n1...nx: are the number of germinated seed on day 1 to day x.
Accelerated Aging Test (AAT) was evaluated by keeping un-imbibed seeds at high temperature (45°C) and relative humidity (around 100%) for 3 days. The seeds were then removed from the stress conditions and placed under optimum germination conditions (28°C) for 14 days (ISTA, 2006). After germinating, the number of normal seedling was counted as mean the potential of the seed could produce normal seedlings after exposed to stress conditions.
The cold emergence test (Cold) is an index for identifying the seed vigor which is identical to the standard germination test. However, the seeds were exposed to 10°C for 7 days prior to the warm greenhouse bench environment. The normal seedlings were counted after 14 days (ISTA, 2006).
The percentage of field emergence (Field) and uniformity of seedling under field condition were determined by Brix` test (ISTA, 2006).
Calculation and statistical analysis: The data are presented as Mean±SD. The analysis of variance (ANOVA) technique was performed for data analysis and differentiated with least significant difference (LSD) test (p = 0.05) using the software STATISTIX (SX-release 8.0, Analytical software, Tallahassee, USA). Regression analysis was carried out to establish the relationship between various seed quality and quantify the same.
RESULTS
MC at the beginning of storage (0 month) ranged from 11.18-11.50 g 100g-1 DM in each treatment. However, MC increased after 12 months storage especially the CA. CL and E+CL was no significantly different from CO (Fig. 1a). At the beginning of storage, Aw ranged from 0.628-0.642 in each treatments. During the storage period, Aw was significantly affected by the seed treatment variants. After 12-months storage, CA seeds had significantly increased Aw to 0.824. However, CL and E+CL treatments were non-significantly different when compared with CO (Fig. 1b).
As presented in Fig. 2a, the potential of the seed to germinate before storage ranged from 94-98% in each treatment. After 12-months storage, the germination of CA seeds was decreased to 60% and the number of abnormal seedlings was increased. The seedlings abnormalities were appeared in roots and coleoptiles. These varied significantly between CA, CL and E+CL treatments, thick and short roots showed higher incidence followed by thin, weak and long roots. In most cases, coleoptiles were either disproportionately short or missing. However, the biological coated seed could maintain the high level of germination percentage, which were non-significantly different from CO (Fig. 3).
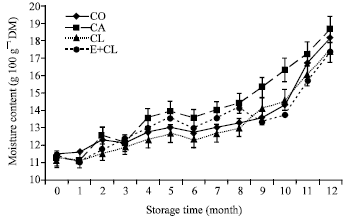 | |
| Fig. 1a: | The effects of seed coating substances and storage time on seeds moisture content (MC). The data are presented as Mean±SD |
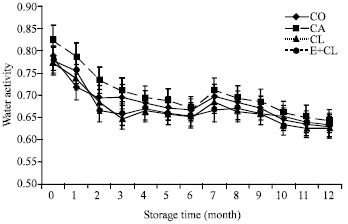 | |
| Fig. 1b: | The effects of seed coating substances and storage time on seeds water activity (Aw). The data are presented as Mean±SD |
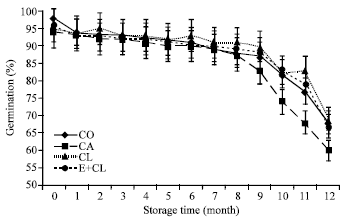 | |
| Fig. 2a: | The effect of seed coating substances and storage time on seeds germination. The data are presented as Mean±SD |
At the beginning of storage (0 month), seed treatment variants did not affected seed viability. However, the seed viability of CA declined rapidly to 85% after storage for 8 months and reduced further to 62% after 12 months of storage. In the case of biological treatments, seed viability did not changed appreciably during storage although slightly changes were observed. This result was not significantly different when compared with CO (Fig. 2b).
A reduction of both shoot and root length was affected by CA. Dynamic changes in shoot and root length occurred during storage period. Biological treatments did not significantly affected on the reduction of both shoot and root length (Fig. 4a, b).
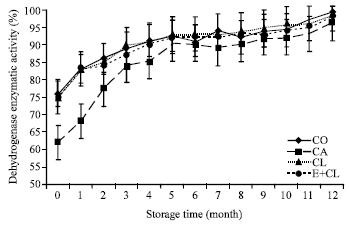 | |
| Fig. 2b: | The effect of seed coating substances and storage time on seed viability (dehydrogenase enzymatic activity). The data are presented as Mean±SD |
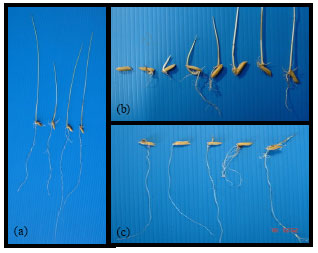 | |
| Fig. 3: | Seedlings damage caused by CA that impacted to the coleoptiles and shoot compared with normal seedlings (A) Normal seedlings (B) Decay and no root and (C) Decay in coleoptiles and shoots |
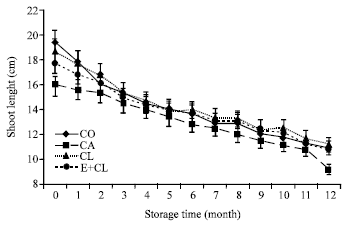 | |
| Fig. 4a: | The effect of seed coating substances and storage time on seedlings shoot length. The data are presented as Mean±SD |
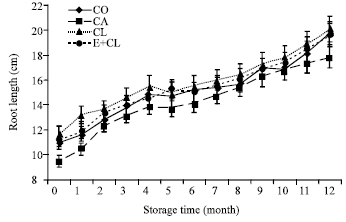 | |
| Fig. 4b: | The effect of seed coating substances and storage time on seedlings root length (B). The data are presented as Mean±SD |
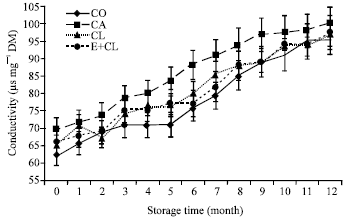 | |
| Fig. 5a: | The effect of seed coating substances and storage time on seeds conductivity. The data are presented as Mean±SD |
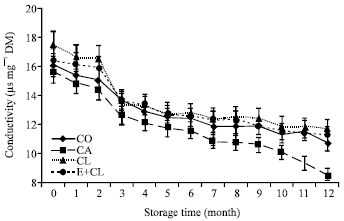
At the beginning of storage (0 month), conductivity values of the CA, CL and E+CL were 69.61, 65.42 and 66.22 μS mg-1, respectively. The level of conductivity of CO was the lowest (62.37 μS mg-1). The conductivity increased throughout the storage time. After 12-months storage, the highest value of conductivity was reached in CA. However, the difference of conductivity between CL, E+CL and CO was found to be non-significant (Fig. 5).
CA also reduced the potential of the seed for the efficient usage of the reserve to produce a vigorous seedling. Figure 5 shows significant differences in SGR between the seed treatment variants. Both CL and E+CL treatments showed constantly the highest level of SGR throughout the storage duration. These treatments provided significantly higher SGR than control seeds. On the contrary, CA treatment showed the lowest of SGR. Moreover, SGR was also affected by the storage duration. The reduction of SGR was observed throughout the storage duration.
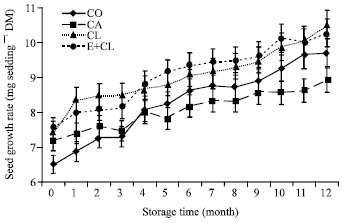 | |
| Fig. 5b: | The effect of seed coating substances and storage time on seedling growth rate. The data are presented as Mean±SD |
 | |
| Fig. 5c: | The effect of seed coating substances and storage time on seedling dry weight. The data are presented as Mean±SD |
CA did not affect only shoot and root length, but it led to the reduction of seedling dry weight. The results demonstrated a strong correlation between cell elongation and seedling dry weight, which was affected by CA. However, seedling dry weight was reduced more drastically than root and shoot lengths (Fig. 5). This result indicated that CA affected the reduction of seedling vigor greater than the biological coated seeds.
At the beginning of storage (0 month), CA treatment showed the lowest GI (Fig. 5d) (16.1). However, the biological coating substances did not adversely affected GI. This result is non-significant different with CO. After 12-months storage, both CL and E+CL treatment maintain the high level of GI, but the CA treatment inhibited seedling development as it is shown by the significant lower GI level compared with the control.
Figure 6a shows, at the beginning of storage (0 month), that AAT results were not affected by seed coating variants. However, the germination declined after 9-months of storage, which was similar to the trends of other vigor indices.
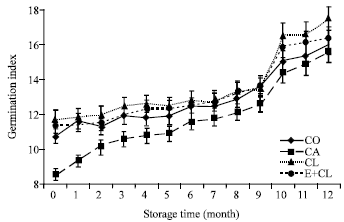 | |
| Fig. 5d: | The effect of seed coating substances and storage time on germination index. The data are presented as Mean±SD |
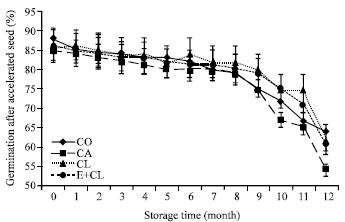 | |
| Fig. 6a: | The effect of seed coating substances and storage time on seed that could germinated under stressful conditions; seed germination under high moisture and humidity (accelerate aging test). The data are presented as Mean±SD |
The tests were highly correlated across all treatments, especially CA. However, CL and E+CL remained a high germination rate of the aged seed. Furthermore, after 12-month storage, CA seriously reduced the potential of seeds that could germinate under stress conditions. On the contrary, CL and E+CL kept the high level of germination of the aged seeds, which were non-significant different with CO.
At the beginning of storage time (0 month), CA showed the lowest germination percentage after being exposed to cold stress condition but the germination of CO and biological coated seeds were similar (Fig. 6b). Eventually, the results indicated that CA caused losses of the viability under cold stress conditions. After stored for 12 months, CA apparently decreased the germination percentage. This result was significantly different from CO. CL and E+CL could maintain the germination percentage, which was significantly different from CO.
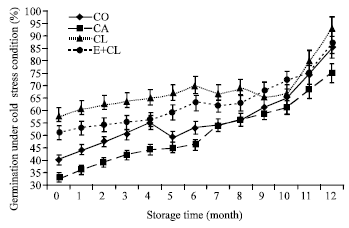 | |
| Fig. 6b: | The effect of seed coating substances and storage time on seed that could germinated under low temperature (cold test). The data are presented as Mean±SD |
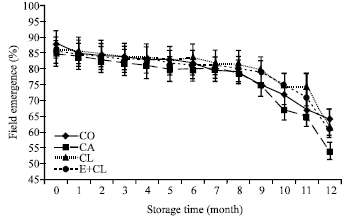 | |
| Fig. 7: | The effect of seed coating substances and storage time on field emergence. The data are presented as Mean±SD |
Both seed treatment variants and storage time decreased the number of germinating seedling under field conditions. At the beginning of storage, CA gave a lower number of germinating seedlings than CO, CL and E+CL did (Fig. 7). After 7-months of storage, CA conclusively inhibits seeds that could germinate under field conditions. These results were significantly different with biological treatment. After 12-months storage, CA inhibited the potential of seeds to germinate under field condition (Fig. 4). The observed results were significantly different in biological coated seeds. Those variants indicated the highest potential to germinate under field condition.
According to the correlation analysis, there was a positive correlation between several seeds quality; Germ, TZ, Sh, Ro, SGR, SDW, GI, AAT and Cold and field emergence. On the other hand, MC, Aw and Cond had reverse correlation with field emergence (Table 1).
| Table 1: | Correlation between various seed quality attributes and field emergence as affected by seed coating substances |
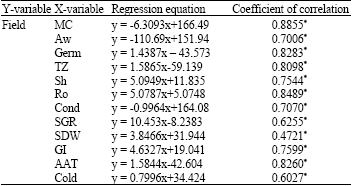 | |
| *: Indicate the statistically significant difference by LSD at 5% level | |
DISCUSSION
In rice seed, as well as in other crops, seed properties are the most commonly used criteria for measuring seed deterioration. The results of this study have enhanced the understanding of the effects of various seed coating substances and storage time on seed quality. CA increased MC and Aw, the increasing rate of water uptake by the CA is probably caused by the decrease of water potential gradient between seeds and their surrounding media. This is the primary effect of CA, which induces the seed moisture content as confirmed by the positive correlation between seeds MC and field emergence and Aw and field emergence (Table 1) (US patent, 48121159). Moreover, the present study showed that CA decreased seed germination and seedling vigor greater than the CL and E+CL did. According to Bugbee (1994) captan produced Cl– which is translocated across the cell membrane into the cytoplasm of the cells unless an active metabolic pump prevents accumulation of the ions. Cl– in the cytoplasm can result in toxic accumulation of a particular ion or decreased availability of some essential nutrients (Bewley and Black, 1994). Additionally, the accumulation of Cl- in the cell affected the potential of seeds to germinate and the seedling performances (Bradford, 1995). Moreover, captan contain active ingredients from the triazole family and both tetrahydrophthalimide and tetrahydrophthalic acid (Paulsen et al., 1981). These compounds delay or even reduce seedling performances (Lin and Sung, 2001). Seedlings from CL and E+CL coated seeds showed long stem and the extensive root system. The results suggested that chitosan treatment might induce the activity of phytohormones inside the seeds. Normally, after seeds imbibitions, phytohormones are activated to induce seed germination and seedling development. Under chemical stress conditions it is possible that the activation of plant hormones in seeds is declined (Hutton et al., 1982). Exogenous application of phytohormones could be ameliorate the inhibitory effect of chemical stress and induce seed to germinate (Chandrkrachang, 2002). Several studies reported that chitosan polymer acting like phytohormones, i.e. auxin (Khairullin et al., 2003), kinetin (Bozcuk, 1981) and cytokinin (Kurosaki et al., 1987), which alleviated fungicide stress conditions. Furthermore, because of the increased cell division and elongation, chitosan strengthen shoot and root systems (Petrukhina et al., 1994). In contrast, after CA treatment, the shoot and root systems were weak. Hirano et al. (2000) reported that captan inhibits seedling growth by reducing both cell division and cell enlargement via directly inhibition of the production and activation of hormones in the seeds. Kabar and Baltepe (1990) supposed that cytokinin, auxin and kinetin levels could be a limitation factor under captan stress conditions. Therefore, their study explained that an exogenous application of chitosan like-phytohormone effects resulted in improved seed germination and growth of rice seedlings. Many studies explained that the loss of seed viability was caused by damages of macromolecules within the embryogenic tissue, which was resulted from fungicide treatments (Villiers, 1972). However, such damages could be more after chemical than after biological seed treatment. Captan affected the repairing of the macromolecules damages in the seedlings during storage (Berjak and Villiers, 1972).
In CO, fungal infection is one most important factor that induces seed deterioration during storage. E+CL acts as antifungal agent that can protect the seeds from fungal infection during storage. This finding was in agreement with Khin et al. (2006) and Thobunluepop et al. (2007). Chitosan polymer has a very effective combination of attractive characteristics, i.e., stimulates plant growth, provides plant protection and can be mixed well with fungicide agents (Thobunluepop et al., 2008). Therefore, chitosan could be a very useful biocontrol polymer with a good perspective for plant production systems (Bautista-Banos et al., 2003).
However, at high concentrations, essential oils might produce phytotoxicity, which affected the seed viability. Tworkoski (2002) found that essential oils from red thyme, summer savory, cinnamon and clove were the most phytotoxic and caused electrolyte leakage resulting in cell death of rice seed when applied at high concentrations (5 to 10% v/v). Thymol, eugenol and citronellal significantly inhibited corn seed germination and development if applying at the concentration higher than 4 %v/v (Waliwitiya et al., 2005). Others studies on rice (Paranagama et al., 2003), tomato, radish, lettuce and wheat (Mazzafera, 2003) reported that eugenol affected the seed germination when treated at concentrations higher than 4 % v/v.
CONCLUSIONS
In order to ensure a successful rice production, uniform and vigorous seedling is a key element. Without a good stand, the effectiveness of other agronomic inputs is drastically reduced and usually such inputs can never compensate the negative impact of a poor stand. This study has proved that seeds coated with biological substances (CL and E+CL) showed less phytotoxic effects which indicated a good seed quality as high germination, vigor and seedling establishment. On the other hand, germination and vigor declined with increasing age of seeds. However, this study focuses only on seedling properties. It is necessary for future works to study the effects of both chemical and biological coating substances on chemical and biochemical seed properties. The results could be useful for better understanding of seed deterioration mechanisms that are affected by various coating substances.
ACKNOWLEDGMENTS
Authors acknowledge the Nation Research Council of Thailand (NRCT), Royal Thai Government, for research financial support of the present studies, gratefully thanks to Prof. Dr. Udo ter Meullen, Prof. Dr. Wolfgang Lüke for they are valuable suggestions through out the research project.
REFERENCES
- Agrawal, A.A., 2002. Herbivory and maternal effects: Mechanisms and consequences of transgenerational induced plant resistance. Ecology, 83: 3408-3415.
Direct Link - Basra, S.M.A., M. Farooq, R. Tabassam and N. Ahmad, 2005. Physiological and biochemical aspects of pre-sowing seed treatments in fine rice (Oryza sativa L.). Seed Sci. Technol., 33: 623-628.
CrossRefDirect Link - Bautista-Banos, S., M. Hernandez-Lopez, E. Bosquez-Molina and C.L. Wilson, 2003. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Protect., 22: 1087-1092.
CrossRefDirect Link - Berjak, P. and T.A. Villiers, 1972. Ageing in plant embryos. V. Lysis of the cytoplasm in non-viable embryos. New Phytol., 71: 1075-1079.
CrossRef - Bugbee, B., B. Spanarkel, S. Johnson, O. Monje and G. Koerner, 1994. CO2 Crop growth enhancement and toxicity in wheat and rice. Adv. Space Res., 14: 257-267.
CrossRef - Ester, A., H. De-Putter, J.G.P.M. Van-Bilsen, 2003. Film coating the seed of cabbage (Brassica oleracea L. convar. Capitata, L.) and cauliflower (Brassica oleracea L. var. Botryti L.) with imidacloprid and spinosad to control insect pests. Crop Prot., 22: 761-768.
CrossRefDirect Link - Farooq, M., S.M.A. Basra, H.A. Kareem and I. Afzal, 2004. Optimization of seed hardening techniques for rice seed invigoration. Emirates J. Agric. Sci., 16: 48-57.
Direct Link - Hirano, S., M. Hayashi and S. Okuno, 2001. Soybean seeds surface-coated with depolymerised chitins: Chitinase activity as a predictive index for the harvest of beans in field culture. J. Sci. Food Agric., 81: 205-209.
CrossRef - Hutton, M.J., J. Van Staden and J.E. Davey, 1982. Cytokinins in germinating seeds of Phaseolus vulgaris L. I. Changes in endogenous levels within the cotyledons. Ann. Bot., 49: 685-691.
Direct Link - Kabar, K. and S. Baltepe, 1990. Effects of kinetin and gibberellic acid in overcoming high temperature and salinity (NaCl) stresses on the germination of barley and lettuce seeds. Phyton (Horn, Austria), 30: 65-74.
Direct Link - Khin, L.N., N. Nitar, S. Chandrkrachang and F.S. Willem, 2006. Chitosan as a growth stimulator in orchid tissue culture. Plant Sci., 170: 1185-1190.
CrossRefDirect Link - Lin, J.M. and J.M. Sung, 2001. Pre-sowing treatments for improving emergence of bitter gourd seedlings under optimal and sub-optimal termperatures. Seed Sci. Technol., 29: 39-50.
Direct Link - Mazzafera, P., 2003. Effect of the alcoholic extract of eugenol on seed germination. Revista Brasil. Bot., 26: 231-238.
CrossRef - Paranagama, P., T. Abeysekera, L. Nugaliyadde and K. Abeywickrama, 2003. Effect of the essential oils of Cymbopogon citratus, C. nardus and Cinnamomum zeylanicum on pest incidence and grain quality of rough rice (paddy) stored in an enclosed seed box. Food Agric. Environ., 1: 134-136.
Direct Link - Paulsen, M.R., W.R. Nave and L.E. Gray, 1981. Soybean quality as affected by impact damage. Trans. ASAE, 24: 1577-1582.
Direct Link - Thobunluepop, P., C. Jatisatienr, A. Jatisatienr, E. Pawelzik and S. Vearasilp, 2007. Comparison of the inhibitory effect of captan, chitosan-lignosulphonate polymer and eugenol coated seeds against rice seed borne fungi. Proceedings of the Tropentag Conference on International Agricultural Research for Development, October 9-11, 2007, University of Kassel-Witzenhausen and University of Göttingen, pp: 9-11.
- Thobunluepop, P., C. Jatisatienr, A. Jatisatienr, E. Pawelzik and S. Vearasilp, 2008. In vitro Screening of the antifungal activity of plant extracts as fungicides against rice seed borne fungi. Proceedings of the Asia-Pacific Symposium on Assuring Quality and Safety of Agricultural-Foods, August 4-6, 2008, Bangkok, Thailand, pp: 29-29.
- Waliwitiya, R., M.B. Isman, R.S. Vernon and A. Riseman, 2005. Insecticidal activity of selected monoterpenoids and rosemary oil to Agriotes obscurus (Coleoptera: Elateridae). J. Econ. Entomol., 98: 1560-1565.
PubMedDirect Link - Yildirim, E. and C.W. Hoy, 2003. Cyromazine seed treatments to control onion maggot, Delia antiqua, on green onions. J. Econ. Entomol., 96: 1494-1499.
CrossRef