
M. Eghbali
Division of Theriogenology, Department of Veterinary Clinical Science, Faculty of Veterinary Medicine, University of Urmia, Urmia, 57153, Iran
S.M. Alavi-Shoushtari
Division of Theriogenology, Department of Veterinary Clinical Science, Faculty of Veterinary Medicine, University of Urmia, Urmia, 57153, Iran
S. Asri Rezaii
Department of Veterinary Clinical Pathology, Faculty of Veterinary Medicine, University of Urmia, Urmia 57153, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 15 | Page No.: 1964-1968







ABSTRACT
To investigate the effects of copper and superoxide dismutase (SOD) content of seminal plasma on buffalo semen characteristics, 54 semen samples collected from buffalo bulls by a bovine artificial vagina were used. Semen characteristics (motility, viability, morphology, concentration and volume) were recorded. Seminal plasma was harvested by centrifugation and kept frozen until analysis. Seminal plasma copper content was determined by atomic absorption procedure and SOD was measured by using a kit. The mean total copper value of seminal plasma was recorded as 2.51 ± 0.04 mg kg-1 (Mean ± SEM) and the mean total SOD values was 39.02 ± 0.81 IU mL-1. To reduce the range of variability, the data were categorized according to their motility records in 3 groups of Excellent (Ex, >90% motile, n = 33), Good (Go, 80-89% motile, n = 15) and Moderate (Mo, <79% motile, n = 6). The mean motility, viability, copper and SOD values in Ex group was recorded as 92.24 ± 0.51%, 94.00 ± 0.48%, 2.56 ± 0.04 mg kg-1 and 39.52 ± 0.57 IU mL-1, respectively. These values were 81.66 ± 0.62%, 85.26 ± 0.95%, 2.38 ± 0.11 mg kg-1 and 36.48 ± 1.51 IU mL-1 in Go group and 71.66 ± 1.05%, 77.00 ± 2.94%, 2.55 ± 0.10 mg kg-1 and 50.66 ± 2.51 in Mo group, respectively. The mean copper value in Ex group was highly (r = 0.600) correlated with SOD and correlated with sperm motility (r = 0.372) and viability (r = 0.363), while, in Go group it was highly correlated (r = 0.945) with SOD and sperm viability (r = 0.652) and in Mo group it was correlated (r = 0.874) with semen volume only. The mean SOD values in Ex group was highly correlated with sperm motility (r = 0.492) and viability (r = 0.490) and mean copper values, in Go group, it was highly correlated whit sperm viability (r = 0.659) and mean copper values and in Mo group it had no significant correlations with semen parameters. These results suggest that copper and SOD content of the buffalo seminal plasma have an influence on the sperm motility and viability which are the most important factors in semen fertility.
PDF Abstract XML References Citation
How to cite this article
M. Eghbali, S.M. Alavi-Shoushtari and S. Asri Rezaii, 2008. Effects of Copper and Superoxide Dismutase Content of Seminal Plasma on Buffalo Semen Characteristics. Pakistan Journal of Biological Sciences, 11: 1964-1968.
DOI: 10.3923/pjbs.2008.1964.1968
URL: https://scialert.net/abstract/?doi=pjbs.2008.1964.1968
DOI: 10.3923/pjbs.2008.1964.1968
URL: https://scialert.net/abstract/?doi=pjbs.2008.1964.1968
INTRODUCTION
Copper appears to be involved in spermatozoa mobility and it may also act on the pituitary receptors which control the release of LH. In the seminal fluid, the level of copper appears to fall in cases of azoospermia and to increase in oligo- and asthenozoospermia (Pleban and Mei, 1983; Skandhan, 1992) but the findings of different authors are somewhat contradictory. It is true that the concentrations in the ejaculate vary considerably from one day to the next and that they also vary in different fractions from a single ejaculate. The toxic effect of copper on spermatozoa has often been reported. Copper reduces the oxidative processes and glucose consumption, which reduces or abolishes mobility (Skandhan, 1992).
The production of free radicals, either by extracellular lipid peroxidation or in situ by the way of oxidative phosphorylation in the mitochondria, could damage the nuclear membrane (Vishwanath and Shannon, 1997).
Superoxide dismutase (SOD) scavenges both extracellular and intracellular superoxide anion and prevents lipid peroxidation of the plasma membrane. In order to act against H2O2, it must be conjugated with catalase or glutathione peroxidase (GPx). SOD also prevents premature hyperactivation and capacitation induced by superoxide radicals before ejaculation (Agarwal and Prabakaran, 2005).
SODs are ubiquitous metalloproteins that catalyze the dismutation of superoxide anion into hydrogen peroxide and molecular oxygen. Appreciable amounts of SOD and much lower concentrations of GPx and catalase have been reported in ejaculated ram sperm (Abu-Erreish et al., 1978). A correlation between SOD activity and the time of motility loss has been found in human sperm, after exposure to H2O2 (Martí et al., 2003).
Copper is necessary for many enzymes like the Cu-Zn-superoxide-dismutase (SOD), which is involved in cell protection against free (oxygen) radicals. Elevated copper concentrations reduce oxidative processes and glucolysis that may cause immotility and reduced viability (Pesch et al., 2006).
Human seminal plasma possesses both superoxide dismutase (SOD)-like and catalase-like activities. However, little is known about the putative source of these key antioxidant activities in human semen. Zini et al. (2002) reported that catalase-like and SOD-like activities (two major antioxidant activities) are primarily derived from post-epididymal (i.e., seminal vesicle, prostate) secretions.
The findings on the seminal plasma catalase and SOD activities and total antioxidant capacity (TAC) are controversial. Sanocka et al. (1996) showed a significant elevation in intracellular activity of SOD and decreasing in catalase activity in infertile samples. Zini et al. (2002) study showed that seminal plasma activity of SOD in infertile men is significantly greater than in fertile men while catalase activity is not different between these groups.
The seminal plasma of mammals has been reported to contain large amounts of superoxide dismutase (SOD) activity, much higher than that found in other extracellular fluids. Spermatozoa have also been reported to contain SOD activity (Peeker et al., 1997).
Sikka et al. (1995) reported an association between leukocytospermia and ROS has been found to correlate with increased chemokine (Interleukin 8; IL-8) and decreased SOD activity.
Toxic effects of copper on seminal plasma are manifested in the decrease of motile spermatozoa percentage and in decrease of malformed sperm cells (Massanyi et al., 2005).
In boar semen, copper can inhibit enzymes having functional sulfidryl groups, bind to and affect the confirmation of nucleic acids, disrupt pathways of oxidative phosphorylation, although the precise response depends upon the individual properties of metal (Massanyi et al., 2003a).
There is little information available about copper and SOD contents in the buffalo seminal plasma. This study was carried out to: (1) estimate the copper and superoxide dismutase contents of the seminal plasma in buffalo bulls, (2) test whether any correlation exists between these parameters and semen characteristics.
MATERIALS AND METHODS
Animals: Fifty four semen samples were collected by a bovine artificial vagina from sexually mature buffalo bulls (4-5 years old) kept in The Buffalo Breeding Center Northwest of Iran, Urmia (37 ° 33` N, 45 ° 4` E) during the summer and autumn of 2007.
Semen evaluation: Immediately after collection, the ejaculate was placed in a 37 °C water bath and the volume was recorded. Semen motility was evaluated immediately after collection. Gross motility was scored from 0 to 5 on a wet mount of neat semen at 100x magnification (0 = cells present without motion; 5 = very rapid dark swirls). The percentage of progressively motile spermatozoa was estimated by microscopic examination at x400 magnification on a pre-warmed slide (37 °C) according to procedure of Ax et al. (2000). Sperm concentration was measured using spectrophotometer (Vital Scientific- IMV, The Netherlands) and the percentage of viable spermatozoa was estimated by viewing 200 spermatozoa under x1000 magnification using eosin-nigrosin staining method of Barth (2007). The semen samples were cooled at room temperature and transported to the laboratory within 2 h.
Preparation of seminal plasma: Fresh semen was centrifuged (Clements, model 2000, Sydney, Australia) at 5,000 rpm for 10 min. The supernatants were transferred into 1.5 mL tubes, re-centrifuged to eliminate the remaining cells and kept frozen (-20 °C) until analysis within a week.
Data analysis: The obtained data was analyzed by using SPSS software (version 11.5 for Windows; SPSS Inc., Chicago, IL, USA) computer program. Results are quoted as arithmetic mean ± standard error of mean (SEM) and significance was attributed at p<0.05.
Pearson`s correlation coefficient (two tailed) test was used to examine the correlation between all the parameters of the semen. The comparison of the semen parameters and copper and SOD contents of the seminal plasma in groups of samples was carried out by Tukey HSD test.
RESULTS
The results of the semen evaluation in Table 1 shows the total copper content of the seminal plasma was recorded as 2.51 ± 0.04 mg kg-1, while, the superoxide dimutase value was 39.92 ± 0.81 IU mL-1. In order to have a better insight of these results and make the range of variations narrower, the samples were categorized in three groups of Excellent (Ex, >90% motile) (n = 33), Good (Go, 80-89% motile) (n = 15) and Moderate (Mo, <79% motile) (n = 6) according to their progressive motility rates. The mean values for progressive motility were recorded as 92.24 ± 0.51% in Ex, 81.66 ± 0.62% in Go and 71.66 ± 1.05% in Mo groups, which were significantly different. The mean copper value in Ex group (2.56 ± 0.04 mg kg-1) was positively correlated with sperm progressive motility and viability (p = 0.033 for both) and highly positively correlated with seminal plasma SOD values (p = 0.000). The mean copper value in Go group (2.38 ± 0.11 mg kg-1) was highly positively correlated with sperm viability and SOD values (p = 0.008 and p = 0.000), but in Mo group (2.55 ± 0.10 mg kg-1) had a positive significant correlation with semen volume (p = 0.022) only (Table 2). The observed negative correlations of the mean copper value in this group (Mo) with sperm gross and progressive motility, viability, concentration and seminal plasma SOD values were not statistically significant.
| Table 1: | Semen characteristics of the buffalo semen (Mean ± SEM) (n = 54) |
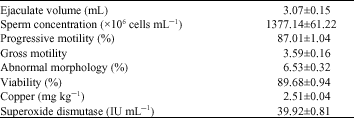 | |
| Table 2: | The comparison of the characteristics of three groups of seminal plasma samples |
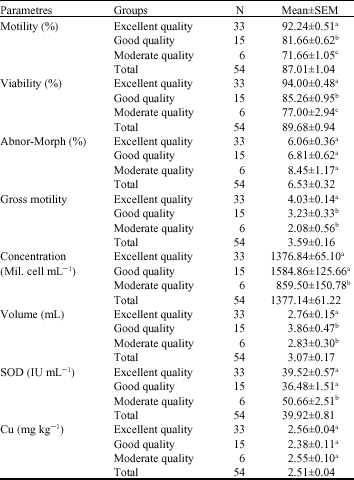 | |
| Mean values denoted by different letter(s) (a, b and c) are significantly different (p<0.05) | |
DISCUSSION
The copper content of the semen has been reported to have an effect on spermatozoa number and motility (Skandhan, 1992) and also on prevention of lipid peroxidation in spermatozoa membrane via the activity of superoxide dismutase. This study was performed to investigate the copper content of the seminal plasma in water buffalo bulls and its correlation with superoxide dismutase, that is reported to be involved in its structure (Pesch et al., 2006) and with other parameters used for semen evaluation.
The mean total copper value of the buffalo seminal plasma in this study was recorded as 2.51 ± 0.04 mg kg-1. Since the specific gravity for seminal plasma in this study was recorded as 1030 mg, by conversion of the unit, the total copper content of one litter of the seminal plasma may be a little less (2.43 mg L-1) which does not affect the results noticeably. The mean copper value in Ex group (2.56 ± 0.04 mg kg-1) which was higher but not significantly different with the other two (Go and Mo) groups, was positively correlated with sperm progressive motility and viability and highly positively correlated with seminal plasma SOD values. The mean copper value in Go group (2.38 ± 0.11 mg kg-1) was highly positively correlated with sperm viability and SOD values, but in Mo group (2.55 ± 0.10 mg kg-1) had a positive significant correlation with semen volume only. The observed negative correlations of the mean copper value in this group with sperm gross and progressive motility, viability, concentration and seminal plasma SOD values were not statistically significant. This may be the effect of a small number of samples in this group.
Massányi et al. (2003b) determined the seminal concentrations of copper and other trace elements in different animals. They observed that the seminal copper concentration was significantly higher in ram (2.49 ± 0.18 mg kg-1) and fox (2.16 ± 0.53 mg kg-1 than that in bull (1.64 ± 0.21 mg kg-1), boar (1.64 ± 0.28 mg kg-1) and stallion (0.86 mg kg-1).
Klemmt and Scialli (2005) stated that human seminal fluid chemical concentrations are typically similar to or lower than blood concentrations, although some antimicrobial agents achieve higher concentrations in semen than in blood. This may be an important factor when adding supplementary copper to the animals` ration.
A positive correlation of copper content of buffalo seminal plasma with sperm motility observed in this study is in agreement with the report of Skandhan (1992) and Massanyi et al. (2005) for the sperm motility but not for the sperm concentration.
Pant and Srivastava (2003) reported that there was no significant difference in copper levels among the different infertile categories. They observed a positive correlation between copper and fructose (r = 0.81, r = 0.72, p<0.05) in oligoasthenospermic and azoospermic men, respectively. This is in agreement with our results for Moderate (Mo) group although we did not further categorized this group.
The total SOD activity observed in buffalo seminal plasma here was 39.92 ± 0.81 IU mL-1. Nair et al. (2006) reported SOD activity in seminal plasma of buffalo bulls in India as 0.86 ± 0.03 U mg-1 protein and it increased to 1.91 ± 0.02 U mg-1 after 72 h storage at refrigerator temperature which is very different with our results. The SOD activity in seminal plasma has been reported to be 27 ± 10.8 U mL-1 in the canine (Cassani et al., 2005) and 5.89 ± 0.96 in normal human subjects (Khosrowbeygi and Zarghami, 2007).
Siciliano et al. (2001) and Hsieh et al. (2002) showed that there is no significant difference in seminal plasma or sperm SOD activity between normozoospermic and oligo-or asthenozoospermic males and also Khosrowbeygi and Zarghami (2007) observed that activities of SOD did not correlate significantly with sperm motility and concentration. This is in agreement with our results of correlations between SOD and sperm concentrations and contradictory to the results of its correlation with sperm motility in Ex group.
Vishwanath and Shannon (1997) reviewed the details of the peroxidation reactions in semen storage and described the exact role of copper and SOD in these reactions.
In mammals there are three SOD isoenzymes, the cytosolic dimeric CuZn-SOD, the mitochondrial matrix Mn-SOD and the secretory tetrameric extracellular SOD (EC-SOD). Plasma also contains EC-SOD with reduced heparin-affinity resulting from truncations and other modifications of the carboxyterminal ends. Since the substrate, the superoxide anion radical, crosses membranes poorly, the SOD isoenzymes must exert distinct protective roles in their respective compartments (Peeker et al., 1997). In the Peeker et al. (1997) study the occurrence and distribution of CuZn-SOD, Mn-SOD and EC-SOD in human seminal plasma and spermatozoa were investigated. The activity of the cytosolic CuZn-SOD of spermatozoa was exceptionally high, but the activity of the mitochondrial Mn-SOD was low. The content of the secretory EC-SOD was very low. Prostate gland appears to be the main source of CuZn-SOD according to the split ejaculate study. The activity in the final seminal plasma is ~10% of that of prostate gland cytosol. Some apocrinic secretion may occur in the prostate gland epithelium, which might explain the presence of CuZn-SOD in the seminal plasma (Peeker et al., 1997).
Cassani et al. (2005) stated that increased SOD activity in seminal plasma, when there is a decreased activity in spermatozoa, could be attributed to the enzyme loss from damaged spermatozoa. The increase of the plasma membrane permeability in this type of spermatozoa seems to permit the loss of the enzyme as was detected in spermatozoa with altered plasma membrane permeability as a consequence of the cryopreservation process. This may be the case for a significantly higher (50.66 ± 2.51 versus 39 ± 0.57 IU mL-1) level of SOD in our Mo group.
Nissen and Kreysel (1983) reported that SOD detected in human seminal plasma inhibits lipid peroxidation and there is a good relationship between SOD-activity and sperm motility; a similar effect has been reported by Skandhan (1992). This is in agreement with our results in Ex group.
Lewis et al. (1995) stated that superoxide dismutase (SOD) activity in seminal plasma appears to have no correlation with percentage motility in infertile men. This does not apply to our results of Ex group but is in agreement with the results of Go and Mo groups.
These observations reveal that copper and SOD content of seminal plasma are important parameters correlated with sperm motility and viability, which are considered to be the most important factors in semen fertility after being inseminated to the females.
CONCLUSION
It can be concluded that copper and superoxide dismutase content of the seminal plasma in buffalo bulls have an effect on sperm motility and viability. Low levels of copper in seminal plasma may be observed in the semen samples of poor quality, while its high levels, accompanied with high levels of SOD, might be indicative of some cell damage.
ACKNOWLEDGMENTS
We would like to thank the authorities of the Buffalo Breeding Center in Northwest of Iran for their support and the supply of buffalo semen.
REFERENCES
- Abu-Erreish, G., L. Magnes and T.K. Li, 1978. Isolation and properties of superoxide dismutase from ram spermatozoa and erythrocytes. Biol. Reprod., 18: 554-560.
PubMed - Agarwal, A. and S.A. Prabakaran, 2005. Oxidative stress and antioxidants in male infertility: A difficult balance. Iran. J. Reprod. Med., 3: 1-8.
Direct Link - Alvarez, J.G. and B.T. Storey, 1983. Role of superoxide dismutase in protecting rabbit spermatozoa from O2 toxicity due to lipid peroxidation. Biol. Reprod., 28: 1129-1136.
PubMed - Hsieh, Y.Y., Y.L. Sun, C.C. Chang, Y.S. Lee, H.D. Tsai and C.S. Lin, 2002. Superoxide dismutase activities of spermatozoa and seminal plasma are not correlated with male infertility. J. Clin. Lab. Anal., 16: 127-131.
PubMed - Klemmt, L. and A.R. Scialli, 2005. The transport of chemicals in semen. Birth Defects Research Part B. Birth Defects Res. Part B: Dev. Reprod. Toxicol., 74: 119-131.
CrossRefPubMedDirect Link - Khosrowbeygi, A. and N. Zarghami, 2007. Levels of oxidative stress biomarkers in seminal plasma and their relationship with seminal parameters. BMC Clin. Pathol., 7: 1-6.
PubMed - Lewis, S.E.M., P.M. Boyle, K.A. McKinney, I.S. Young and W. Thompson, 1995. Total antioxidant capacity of seminal plasma is different in fertile and infertile men. Fertil. Steril., 64: 868-870.
PubMed - Massanyi, P., J. Trandzik, P. Nad, R. Toman, M. Skalicka and B. Korenekova, 2003. Seminal concentrations of trace elements in various animals and their correlations. Asian J. Androl., 5: 101-104.
PubMedDirect Link - Massanyi, P., J. Trandzik, P. Nad, M. Skalicka and B. Korenekova et al., 2005. Seminal concentration of trace elements in fox and relationships to spermatozoa quality. J. Environ. Sci. Health Part A, Tox Hazard Subst. Environ. Eng., 40: 1097-1105.
PubMed - Nair, S.J., A.S. Brar, C.S. Ahuja, S.P.S. Sangha and K.C. Chaudhary, 2006. A comparative study on lipid peroxidation, activities of antioxidant enzymes and viability of cattle and buffalo bull spermatozoa during storage at refrigeration temperature. Anim. Reprod. Sci., 96: 21-29.
CrossRefPubMedDirect Link - Nissen, H.P. and H.W. Kreysel, 1983. Superoxide dismutase in human semen. Klin Wochenschr, 61: 63-65.
PubMed - Pant, N. and S.P. Srivastava, 2003. Correlation of trace element concentration with fructose, gamma-glutamyl transpeptidase and acid phosphatase in seminal plasma of different categories of infertile men. Biol. Trace Element Res., 93: 31-38.
PubMed - Peeker, R., L. Abramsson and S.L. Marklund, 1997. Superoxide dismutase isoenzymes in human seminal plasma and spermatozoa. Mol. Hum. Reprod., 3: 1061-1066.
PubMed - Pesch, S., M. Bergmann and H. Bostedt, 2006. Determination of some enzymes and macro- and microelements in stallion seminal plasma and their correlations to semen quality. Theriogenology, 66: 307-313.
CrossRefPubMedDirect Link - Pleban, P.A. and S. Mei, 1983. Trace elements in human seminal plasma and spermatozoa. Clin. Chem. Acta, 133: 43-50.
PubMed - Sanocka, D., R. Meisel, P. Jedrzejczak and M.K. Kurpisz, 1996. Oxidative stress and male infertility. J. Androl., 17: 449-454.
PubMed - Siciliano, L., P. Tarantino, F. Longobardi, V. Rago, C. De Stefano and A. Carpino, 2001. Impaired seminal antioxidant capacity in human semen with hypervicosity or olygoasthenozoospermia. J. Androl., 22: 798-803.
PubMed - Sikka, S.C., M. Rajasekaran and W.J. Hellstrom, 1995. Role of oxidative stress and antioxidants in male infertility. J. Androl., 16: 464-481.
Direct Link - Skandhan, K.P., 1992. Review on copper in male reproduction and contraception. Revue Francaise Gynecologie d'Obstetrique, 87: 594-598.
PubMedDirect Link - Vishwanath, R. and P. Shannon, 1997. Do sperm cells age? A review of the physiological changes in sperm during storage at ambient temperature. Reprod. Fert. Dev., 9: 321-331.
PubMed - Zini, A., K. Garrels and D. Phang, 2000. Antioxidant activity in the semen of fertile and infertile men. Urology, 55: 922-926.
PubMed