
M. Boungou
Laboratoire de Biologie et Physiologie Animales, Unité de Formation et de Recherche en Sciences de la Vie et de la Terre (UFR/SVT), Université de Ouagadougou, 03 BP 7021, Ouagadougou 03, Burkina Faso
G.B. Kabre
Laboratoire de Biologie et Physiologie Animales, Unité de Formation et de Recherche en Sciences de la Vie et de la Terre (UFR/SVT), Université de Ouagadougou, 03 BP 7021, Ouagadougou 03, Burkina Faso
A. Marques
UMR 5119 ECOLAG cc 93 Université Montpellier II, 34095 Montpellier Cedex 05, France
L. Sawadogo
Laboratoire de Biologie et Physiologie Animales, Unité de Formation et de Recherche en Sciences de la Vie et de la Terre (UFR/SVT), Université de Ouagadougou, 03 BP 7021, Ouagadougou 03, Burkina Faso
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 10 | Page No.: 1317-1323







ABSTRACT
The study of the gill fishes of 176 Oreochromis niloticus (L.) was undertaken in the dam of Loumbila (Burkina Faso), in order to obtain the first information on the dynamics of population of the Monogeneans parasites in a population of wild hosts. These data are essential in the search for means to avoid the enormous losses that often occur in intensive pisciculture. This analysis revealed the presence of five species of Monogeneans (Cichlidogyrus tilapiae, Cichlidogyrus halli, Cichlidogyrus thurstonae, Cichlidogyrus rognoni and Scutogyrus longicornis). On the levels of the recorded infestations, these species taken individually probably do not control the population of Oreochromis niloticus. In the dam of Loumbila, the fluctuations of parasitic abundances are neither seasonal nor cyclic. The size either does not have an influence on parasitism. However, the sex effect was noted with C. halli, the females thus were more infested than the males. O. niloticus is parasitized all the year, because the recruitment of these organisms, although relatively weak, is continuous; this logically results in their accumulation in this fish.
PDF Abstract XML References Citation
How to cite this article
M. Boungou, G.B. Kabre, A. Marques and L. Sawadogo, 2008. Dynamics of Population of Five Parasitic Monogeneans of Oreochromis niloticus Linne, 1757 in the Dam of Loumbila and Possible Interest in Intensive Pisciculture. Pakistan Journal of Biological Sciences, 11: 1317-1323.
DOI: 10.3923/pjbs.2008.1317.1323
URL: https://scialert.net/abstract/?doi=pjbs.2008.1317.1323
DOI: 10.3923/pjbs.2008.1317.1323
URL: https://scialert.net/abstract/?doi=pjbs.2008.1317.1323
INTRODUCTION
The Monogeneans are ectoparasites, mostly of fishes; living hung by their haptor on the skin, fins and gills of their hosts. Nevertheless, some species live in more hidden habitats such as the rectal cavity, nostrils, the urethras, stomach and so on. Their development cycle is direct (no host intermediary) and has a larva with cilia often swimming (oncomiridia), which is the only free stage (Pariselle and Euzet, 1998). In natural environment, Monogeneans generally expand in limited number and are in apparent equilibrium with the host fish. In the conditions of fish culture where the fishes are more concentrated than in natural environment, we sometimes notice a large amount of parasites, leading to epizootics (Buchmann and Lindenstrom, 2002), resulting in high mortality of fishes and, therefore, to a loss of productivity in basins (Aloo, 2002). In addition, severe secondary infections, favored by lesions caused by the hooks of haptor and the despoiling action of the parasites, can contribute to this mortality (Mouton et al., 2001). Therefore, a pre-requisite for the success of fish breeding is to prevent epidemics and the development of potential infections (Obiekezie et al., 1992). This requires a good taxonomic and biological knowledge of pathogen agents.
Monogeneans parasites of continental fishes are mostly known in their taxonomical aspect. Indeed, only the species has caught the attention of most of the researchers. They were interested in the taxonomy, the specificity and biological cycle of Monogeneans parasites (Tombi and Bilong Bilong, 2004). Quantitative data on population dynamics of Monogeneans parasites remain scarce, particularly in tropical Africa (Mouton et al., 2001). So far, only a few study such as Mbahinzireki (1980), Khidr (1990), Obiekezie et al. (1992), Aloo (2002) and Bilong Bilong and Tombi (2005) indicate that almost the prevalence of parasites of fishes of African continental have been identified.
The results presented here are part of a comprehensive study of parasites of freshwater fishes from Burkina Faso and concern the dynamic of populations of five gills Monogeneans of Oreochromis niloticus in the dam of Loumbila (Burkina Faso). Are considered here the rates of infestation of different stages of maturity of parasites in function of time, sex and size of the host. The objective of this research is to study the demographic profiles of Monogeneans parasites of the fish model, in a semi-confined area close to that of the basins of pisciculture, in order to gather the first data relating to parasitism of Burkina Faso`s fishes.
MATERIALS AND METHODS
The study site:Loumbila`s dam is located at 20 km from Ouagadougou in the department that bears the same name and in the province of Oubritenga. Its geographic coordinates are the following: 12°29`34" north latitude and 01°24`05" west longitude. This water retention has a capacity of 42 million m3 (ONEA and CASST, 2004). This dam is inhabited by ichthyology fauna that allow the development of a traditional fishing. Three families of fishes coexist: Cichlid especially those represented by O. niloticus and very little by Sarotherodon galilaeus are numerically greater than that of Claridae with Clarias sp. and that of the Centroponidae with Lates niloticus.
The host:The host used in this study is Oreochromis niloticus (L.). It is a fish that has great economic interest in Burkina Faso (Baijot et al., 1994). Its use is therefore widespread in development projects of the fish culture in that country. This study was carried from February 2004 in January 2005. During this period, we harvested and autopsied 176 individuals from Oreochromis niloticus (L.), or an average of 15 fish per month.
The sex ratio for this sub-population is 0.83 for males, who were 96 individuals against 80 females. Fish, whose Standard Length (LS) ranged from 70 to 194 mm, have been placed in 08 classes sizes, which are: Class A (LS <85 mm), with 9.09% of the specimens; Class B (85 = LS <95 mm), with 5.11% of specimens; Class C (95 = LS <105 mm) with 6.81%, the D (105 = LS <115 mm) with 15.90% of specimens; class E (115 = LS <125 mm) with 14.20% of specimens; class F (125 = LS <135 mm) with 31.81% of specimens; class G (135 = LS <145) with 7.38% of specimens and class H (LS (145 mm) with 9.65% of specimens.
Fishing and fixing of the biological material:The fish examined were caught by fishermen using fishnets. Once caught, the fish were immediately sacrificed; their gill arches were isolated from the bucco-pharyngeal cavity by dorsal and ventral sections and then frozen in liquid nitrogen.
Research, coloration and identification of the parasites:In the laboratory, after thawing, the parasites were detached from the gills using strong water current and transferred individually with a needle directly into a drop of ammonium picrate-glycerine mixture (Malmberg, 1957). The preparation was then covered with a cover slip and sealed with Glyceel (Gurr, BDH Chemicals).
The identification of species and determining the degree of maturity of individuals` parasites were made under a microscope. Two stages of maturity have been identified: adult or mature (testis, ovary and sclerotised pieces of the observable apparatuses copulator) and the young or immature (presence of only male genitalia or total absence of genitalia apparatuses).
Epidemiological approach:We have studied the parasitism of this host by Monogeneans in function of time, size and sex of the hosts. As a result, the abundance or infestation rate (expressed as a percentage) is the total number of parasites in a sample of hosts divided by the total number of hosts examined (Margolis et al., 1982). We used the software STATISTICA version 6 to analyze the data and the comparison of the rates of infestation was made by the means of Chi-square tests (χ2). The level of significance was fixed to 0.05.
RESULTS
Five species of Monogeneans were collected on the gills of the fish: Cichlidogyrus tilapiae, Cichlidogyrus halli, Cichlidogyrus thurstonae, Cichlidogyrus rognoni and Scutogyrus longicornis.
Temporal variation of the occurrence of the parasites
Cichlidogyrus thurstonae:C. thurstonae was present in O. niloticus throughout the study period. The lowest rate of infestation was observed in July 2004 (21.42%). The highest values were observed during the month of June 2004 with a rate of 68.75% and in the month of September 2004 with a rate of infection of 73% (Table 1). The Chi-square test (χ2) applied to the temporal variations of the occurrence of C. Thurstonae shows that the values taken in a comprehensive manner are not statistically significant at 5% (χ2 = 13.51, df = 11).
Cichlidogyrus halli:This species of Monogenean was present in O. niloticus during all the study period. Although the values of the rates of infestation are overall a little low compared to the first parasite described, we observed that the variation of the infestation rate were serrated. The lowest rate of infestation was observed in April 2004 with a value of 20%. The highest value in the rate of infestation was observed in September 2004 (60%). The Chi-square test (χ2) applied to the temporal variations of the occurrence of C. halli shows that these values
| Table 1: | Temporal variation of different species |
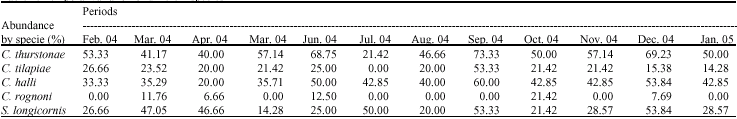 | |
taken in a comprehensive manner are not statistically significant at 5% (χ2 = 7.19; df = 11), like that was show by the Table 1.
Cichlidogyrus tilapiae:The rate of infestation varies from 26.66% in February with a peak of 53.33% in September. There was a lack of parasites in July 2004. The Chi-square test (χ2) applied to the temporal variations of the occurrence of C. tilapiae shows that these values taken in a comprehensive manner are not statistically significant at 5% (χ2 = 13.65, df = 11).
Cichlidogyrus rognoni:We observed that the parasite was present in March, April, October and December 2004 with low values respectively 11.76, 6.66, 12.5, 21.42 and 7.69%. The Chi-square test (χ2) applied to the temporal variations of the occurrence of C. rognoni shows that these values taken in a comprehensive manner are not statistically significant at 5% (χ2 = 16.72, df = 11).
Scutogyrus longicornis:We note that this species of Monogeneans was present in O. niloticus throughout the study period. Although the values of infestation rates are low in all, we observed a variation in the rates of infestation was serrated. The lowest rate of infestation was observed in May 2004 with a value of 14.28% (Table 1). The highest rate of infestation was observed during the month of September 2004 (53.33%) and in December 2004 (53.84%). The Chi-square test (χ2) applied to the temporal variations of the occurrence of S. longicornis shows that these values taken in a comprehensive manner are not statistically significant at 5% (χ2 = 14.59, df = 11).
Occurrence of the parasites according to the sex of the host
Cichlidogyrus thurstonae:The rate of infestation of O. niloticus by C. thurstonae was 48.95% for male fish and 56.25% for female fish. These values were not statistically significant at 5% (χ2 = 0.92, df = 1).
Cichlidogyrus halli:The rate of infestation of O. niloticus by C. halli was 33.33% for male fish and 50% for female fish. We note that the females were much more parasitized than males. However, the Chi-square test (χ2) applied at the occurrence of Cichlidogyrus halli depending on the sex of its host, shows that these values are statistically significant (χ2 = 5.01, df = 1).
Cichlidogyrus tilapiae:The rate of infestation of O. niloticus by C. Tilapiae was 21.87% (21 male fish parasites on the 96 examined) and 22.50% (18 female fish parasitized on the 80 examined). These values were not statistically significant at 5% (χ2 = 0.01, df = 1).
Cichlidogyrus rognoni:The rate of infestation of O. niloticus by C. tilapiae was 4.16% for male fish (4 fish parasites males on the 96 examined) and 7.50% for female fish (6 female fish parasitized on the 80 examined). These values were not statistically significant at 5% (χ2 = 0.90, df = 1).
Scutogyrus longicornis:The rate of infestation of O. niloticus by S. longicornis was 31.25% for male fish and 38.75% for female fish. These values were not statistically significant at 5% (χ2 = 1.08, df = 1).
Occurrence of the parasites according to the size of the host
Cichlidogyrus thurstonae:The lowest rate of infection was observed in fish belonging to the size class H (41.17%). The highest value was observed in the fish of the Class B (66.66%). The Chi-square test (χ2) applied at the occurrence of C. thurstonae depending on the size of O. niloticus shows that these values are not statistically significant at 5% (χ2 = 2.20; df = 7). So, C. thurstonae may parasitize O. niloticus at any age (Table 2).
Cichlidogyrus halli:The highest rate of infestation was encountered in classes A, E, F and G respectively with 56.25, 48, 42.85 and 53.84%. The Chi-square test (χ2) applied at the occurrence of C. halli depending on the size of O. niloticus shows that these values are not statistically significant at 5% (χ2 = 4.93, df = 7). So, C. halli may parasitize O. niloticus at any age.
Cichlidogyrus tilapiae:All classes of fish harvested and studied had hosted C. tilapiae. Classes C, D and H present rate of infestation less important than the other classes, respectively with 33.33, 28.57 and 29.41.
| Table 2: | Occurrence of species depending to the size of the host |
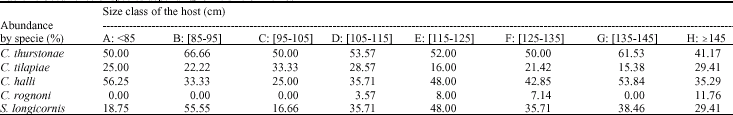 | |
The Chi-square test (χ2) applied at the occurrence of C. tilapiae depending on the size of O. niloticus shows that these values are not statistically significant at 5% (χ2 = 2.81, df = 7). So, C. tilapiae may parasitize O. niloticus at any age.
Cichlidogyrus rognoni:We observed that the fish of size A, B and C had not been infected by C. rognoni. Fish belonging to other classes are parasitized all except the size class G. The small sample size may explain the absence of parasites here. However, the Chi-square test (χ2) applied at the occurrence of C. rognoni depending on the size of O. niloticus shows that these values are not statistically significant at 5% (χ2 = 5.28, df = 7). So, C. rognoni may parasitize O. niloticus at any age.
Scutogyrus longicornis:We note that all classes of O. niloticus collected and studied had harbored S. longicornis. The lowest rate of infection is observed in fish belonging to the size class C (16.66%). The highest value was observed in the fish of the Class B (55.55%). The Chi-square test (χ2) applied at the occurrence of S. longicornis depending on the size of O. niloticus shows that these values are not statistically significant at 5% (χ2 = 7.45, df = 7). So, Scutogyrus longicornis may parasitize O. niloticus at any age (Table 2).
DISCUSSION
During present study on Monogeneans gill parasites of Oreochromis niloticus (L.), we found five species belonging to two genera: the genus Cichlidogyrus with four species and the genus Scutogyrus with one specie.
The exploitation of a host fish by several genera of Monogeneans has been already reported by several authors such as Birgi (1988b) with the species of Clarias jaensis and Clarias pachynema parasitized by the genera Quadriacanthus and Claridectes, Obiekezie et al. (1992), which reported the presence in Clarias gariepinus, the genus Gyrodactylus and several specie of Ancylodiscoides; Koskivaara and Valtonen (1992), who found genera Datylogyrus, Paradiplozoon and Gyrodactylus in Rutilus rutilus; Bilong Bilong (1995) shows that Hemichromis fasciatus was parasited by the genera Cichlidogyrus and Onchobdella; Douellou and Chishawa (1995) who collected the species of genus Quadriacanthus and Macrogyrodactylus in Clarias gariepinus and Tombi and Bilong Bilong (2004) who made an inventory of the genera Dactylogyrus, Dogelius, a larva of Gyrodactylus and of Polystomatidae in Barbus martorelli.
Moreover, the exploitation of hosts by several congeneric species was also reported by Buckmann and Lindenstrom (2002), Tombi (2005) and Simkova et al. (2006). As thought by the authors mentioned above, our observations can be explained by the fact that in natural environment, the parasitic densities are generally weak and therefore, the niches are always available on the gill biotope (Gutiérrez and Martorelli, 1999; Buckmann and Lindenstrom, 2002; Simkova et al., 2006), facilitating the simultaneous colonization of the same host by several species of Monogeneans. In Oreochromis niloticus (L.), we have not observed any significant difference in the rate of the parasitism according to the classes of size. Our results are similar to those of Koskivaara and Valtonen (1992) who did not observe any significant difference of P. Homoion between different classes of sizes R. rutilus or of Pseudodactylogyrus bini and P. anguillae in Anguilla anguilla. However, our observations on the parasitism following the different classes are contrary to those of Bakke et al. (2002) who observed a negative correlation between the growth of the population of Gyrodactylus salaris and age of salmonidae. They explain this by the immune answer that increase with the age of the fish. Present results are also contrary to those of Morand et al. (2002) and Tombi and Bilong Bilong (2004), who found out a positive correlation between the parasitic abundances or intensities with the size of the fish. These authors explained their observation by the fact that as soon as the fish grows up the gill surface enlarges. Gutiérrez and Martorelli (1999) and Simkova et al. (2006) also think that since in big fish the volume of water passing through the gills are higher; the number of oncomiridia might also be high. We think that considering the mode of infestation of Monopisthocotylea the volume of water passing through the gills should not influence the prevalence or intensity parasitic on those biotopes: Indeed, the infesting larva here settle first on the skin before scrawling and settling on the gills (Combes and Jourdane, 2003).
On one side, we have not observed any significant difference between the rates of parasitic infestation in virtually all of the males and females of Oreochromis niloticus, what confirms the observations of Silan et al. (1996) and Tombi and Bilong Bilong (2004). On the other side, following the occurrence of C. halli according to males and females of O. niloticus (L.), the statistics data give a significant difference. The females fish are more attacked by the parasites than males. Present results are similar to those of Tombi (2005), who found that the species D. bopeleti and D. insolitus are more frequently isolated in the female Barbus. These results are also similar to those of Silan et al. (1996), which explained that the females more-sedentary during the period of reproduction are more infested by the Diplectanidae than the males of Dicentracnus labrax.
Concerning the temporal variation of the occurrence of parasites, all species of Monogeneans identified have been present the whole year in the host fish (O. niloticus), except C. rognoni which was absent during in July, August and September (Table 1). Thus, the host fish is vulnerable at any period. Under the tropics, analogous remarks have been reported. Indeed, in Cameroon, Bilong Bilong and Njiné (1998) considered eutrophic basin in urban areas and showed that in this lentic environment the Monogeneans gill parasites of the fish Hemichromis fasciatus appear the whole year. Later, Bilong Bilong and Tombi (2005) studied the temporal structure of six species gill parasites of Barbus martorelli of the Mfoulou stream (under-affluent of the Sanaga river) in the surrounding areas of Yaounde (Cameroon). This study, conducted in oligotrophic water showed that, except for the species of Monogeneans Dactylogyrus maillardi absent during the month of July, all the species of xeno community are present on that Cyprinid during the whole year. The same, in Nigeria, Obiekezie et al. (1992) observe that Macrogyrodactylus clarii appears in a river the whole year. In Zambia, an artificial lake the Monogeneans of the genus Cichlidogyrus in three species of Cichlid are met equally the whole year (Batra, 1984).
Bilong Bilong and Tombi (2005) sums up the importance of fish parasites as follows: they may reduce population numbers by causing mortalities and sometimes mass mortalities, they may affect the reproductive organs and reduce the number of offsprings and thus the population size and they may reduce the weight of fish. Combes and Jourdane (2003) also stressed that the pathogenic effect is rarely caused by one parasite species. It was hereby shown that O. niloticus Monogenean community was made up of cinq species of Dactylogyridae; taking into account all parasite species, infracommunity sizes could be considerable and the sum of pathogenic effects so important that it could cause cases of morbidity or mortality even in a natural environment.
In the prospected site, the addition of the pathogenic effects of the five parasitic species can become important and provoke cases of morbidity (or even of mortality), particularly for fish accommodating greater infra communities. Such mortality can be reinforced by secondary infections due to lesions caused by the hooks of parasites and the spoliatory action of those organisms. It can seem insignificant in a pond of the size of the one prospected, but would certainly expand in ponds in small-scale diversion (250-40 m2, etc.) where the density of fish increase fast enough and where the transmission rate of parasites will therefore be increase. In such a situation and for same species of host, one can fear serious general pathology strongly influenced by the prevalence, abundance and distribution (aggregated or not) of the parasites (Paperna and Smirnova, 1997).
CONCLUSION
At the dam of Loumbila, the fluctuations of the parasitic abundance are neither seasonal nor cyclic. Oreochromis niloticus is infested the entire year; indeed, the recruitment of those organisms, though relatively weak, is continuous. The result is logically their accumulation in that fish.
The struggle against direct cycle parasites like the Monogeneans can be done either by the treatment of hosts or by the breaking off of the development cycle in eliminating the infesting larva from the living environment of the fish. To reduce the parasitic pressure on the host and to prevent the potential damage of those Monogeneans, one can disinfect the genitors before the putting them in the basins. However, if the sowing is done with young fish, which can be less resistant to the effect of pesticides, one can wait until the emptying of water. One will then treat the re-population stock in the basins. After the emptying during which one can spill pesticides into the basins, a lack of parasites should be observed. Even the use of any pesticide require the opinion of a specialist who must respect the local policy (regulating the use of pesticides) and take into account: (a) problems of preserving the environment (b) cases of resistance of the parasites to some molecules as those reported in Nigeria (Opara, 2002). The choice of any substance must therefore be preceded by a test.
REFERENCES
- Bakke, T.A., P.D. Harris and J. Cable, 2002. Host specifity dynamics: Observations on gyrodactylid monogeneans. Int. J. Parasitol., 32: 281-308.
CrossRefDirect Link - Batra, V., 1984. Prevalence of helminth parasites in three species of cichlids from man-made lake in Zambia. Zool. J. Linn. Soc., 82: 319-333.
CrossRefDirect Link - Douellou, L. and A.M.M. Chishawa, 1995. Monogeneans of three Siluriform fish species in Lake Kariba, Zimbabwe. J. Afr. Zool., 109: 99-115.
Direct Link - Gutierrez, P.A. and S.R. Martorelli, 1999. Hemibranch preference by freshwater monogeneans a function of gill area, water current, or both?. Folia Parasitologica, 46: 263-266.
Direct Link - Khidr, A.A., 1990. Population dynamics of Enterogyrus cichlidarum (Monogenea: Ancyrocephalinae) from the stomach of Tilapia spp. In Egypt. Int. J. Parasitol., 20: 741-745.
PubMedDirect Link - Koskivaara, M. and E.T. Valtonen, 1992. Dactylogyrus (Monogenea) communities on the gills parasites of roach in three lakes in Central Finland. Parasitologia, 104: 263-272.
Direct Link - Margolis, L., G.W. Esch, J.C. Holmes, A.M. Kuris and G.A. Shad, 1982. The use of ecological terms in parasitology (Report of an ad hoc committee of the American Society of Parasitologists). J. Parasitol., 68: 131-133.
CrossRefDirect Link - Morand, S., A. Simkova, I. Matejusova, L. Plaisance, O. Verneau and Y. Desdevises, 2002. Investigating patterns may reveal processes: Evolutionary ecology of ectoparasitic monogeneans. Int. J. Parasitol., 32: 111-119.
Direct Link - Mouton, A., L. Basson and D. Impson, 2001. Health status of ornamental freshwater fishes imported to South Africa: A pilot study. Aquarium Sci. Conser., 3: 313-319.
CrossRefDirect Link - Obiekezie, A.I., K.Anders, R. Lick, H. Moeller and H. Palm, 1992. External lesions and flesh parasites in commercial fishes of Nigerian inshore waters. Aquat. Living Resour., 5: 173-183.
CrossRefDirect Link - Opara, K.N., 2002. Population dynamics of Piscicola geometra (Hirudinea: Rhynchobdellida) on Oreochromis niloticus (Cichlidae) cultured in a rainforest fish pond, South Eastern Nigeria. J. Environ. Sci., 14: 536-540.
Direct Link - Paperna, I. And M. Smirnova, 1997. Branchiomyces-like infection in a cultured tilapia (Oreochromis hybrid, Cichlidae). Dis. Aquat. Organisms, 31: 233-238.
Direct Link - Pariselle, A. and L. Euzet, 1998. Five new species of Cichlidogyrus (Monogenea: Ancyrocephalidae) from Tilapia brevimanus, T. buttikoferi and T. cessiana from Guinea, Ivory Coast and Sierra Leone (West Africa). Folia Parasitologica, 45: 275-282.
Direct Link - Simkova, A., O. Verneau, M. Gelnar and S. Morand, 2006. Specificity and specialization of congeneric Monogeneans parasiting Cyprinid. Evolution, 60: 1023-1037.
Direct Link - Tombi, J. and C.F.B. Bilong, 2004. Distribution of gill parasites of the freshwater fish Barbus martorelli Roman, 1971 (Teleostei: Cyprinidae) and tendency to inverse intensity evolution between myxosporidia and monogenea as a function of the host age. Revue d Elevage et de Medecine Veterinaire des Pays Tropicaux, 57: 71-76.
Direct Link - Buchmann, K. and T. Lindenstrom, 2002. Intractions between monogenean parasites and their fish hosts. Intl. J. Parasitol., 32: 309-319.
CrossRefDirect Link