
Salil Kumar Bhowmik
Department of Biotechnology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh
Mirza Mofazzal Islam
Bangladesh Institute of Nuclear Agriculture, P.O. Box 4, Mymensingh 2200, Bangladesh
Reza Mohammad Emon
Bangladesh Institute of Nuclear Agriculture, P.O. Box 4, Mymensingh 2200, Bangladesh
Shamsun Nahar Begum
Bangladesh Institute of Nuclear Agriculture, P.O. Box 4, Mymensingh 2200, Bangladesh
Aysha Siddika
Department of Biotechnology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh
Sharmin Sultana
Department of Biotechnology, Bangladesh Agricultural University, Mymensingh 2202, Bangladesh
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 24 | Page No.: 4449-4454







ABSTRACT
Eleven genotypes, including the salt tolerant cultivar Pokkali as check, were used to evaluate salinity tolerance phenotypically and genotypically. Three selected SSR primers viz., RM7075, RM336 and RM253 were used to evaluate rice genotypes for salt tolerance. Two setups were maintained for this study viz., the seedling and reproductive stages. Phenotyping at the seedling stage was done in hydroponic system using salinized (EC 12 dS m-1) nutrient solution and at the reproductive stage using salinized tap water (EC 6 dS m-1). IRRI standard protocol was followed to evaluate salinity tolerance in rice. The genotypes having similar banding pattern with Pokkali were considered as tolerant. Phenotypically, three genotypes Pokkali, THDB and TNDB-100 and five genotypes RD-2586, TNDB-100, Dhol Kochuri, PNR-519 and Pokkali were identified as salt tolerant at the seedling and reproductive stages, respectively. These genotypes were also identified as salt tolerant genotypically (with markers). Through phenotypic and genotypic study, five genotypes viz., Pokkali, Dhol Kochuri, RD-2586, TNDB-100 and PNR-519 were identified as salt tolerant. Therefore, these microsatellite markers used in this study could be efficiently used for identification of salt tolerant rice varieties in marker-assisted breeding and quantitative trait loci analysis.
PDF Abstract XML References Citation
How to cite this article
Salil Kumar Bhowmik, Mirza Mofazzal Islam, Reza Mohammad Emon, Shamsun Nahar Begum, Aysha Siddika and Sharmin Sultana, 2007. Identification of Salt Tolerant Rice Cultivars Via Phenotypic and
Marker-Assisted Procedures. Pakistan Journal of Biological Sciences, 10: 4449-4454.
DOI: 10.3923/pjbs.2007.4449.4454
URL: https://scialert.net/abstract/?doi=pjbs.2007.4449.4454
DOI: 10.3923/pjbs.2007.4449.4454
URL: https://scialert.net/abstract/?doi=pjbs.2007.4449.4454
INTRODUCTION
Rice (Oryza sativa L.) plant is one of the most suitable crops for saline soil, though it is considered moderately sensitive to saline (Mori and Kinoshita, 1987). The common practice is to cultivate rice with elevated level of salt tolerance on salt affected marginal lands. Today, the most economic and sustained way to overcome the problems of food scarcity and salt stress is to develop salt tolerant varieties. Salinity affects rice growth in varying degrees at all stages starting from germination through maturation (Manneh, 2004). So far, conventional breeding methods for salt tolerance have been found ineffective due to the strong environmental effects on genotypic expression and the low narrow sense heritability of salt tolerance (Gregorio, 1997; Gregorio and Senadhira, 1993). This hinders the development of an accurate, rapid and reliable screening technique under natural conditions. On the other hand, screening under controlled condition has the benefit of reduced environmental effects and the hydroponics system is free from the difficulties associated with soil related stress factors.
With the recent development in the field of molecular markers analysis, it is now feasible to analyze both the simply inherited traits as well as the quantitative traits and then identify the individual genes controlling the traits of interest. Molecular markers now can be used to tag Quantitative Trait Loci (QTLs) and evaluate their contributions to the phenotype by selecting for favourable alleles at these loci in a Marker-Assisted Selection (MAS) scheme that aim to accelerate genetic advancement in rice. Individual genotypes with target gene in a segregating population can be identified with the assistance of DNA markers. Identifying molecular markers that are linked to genes controlling salinity tolerance could facilitate selection in rice for this low heritable trait. MAS in rice is faster, more efficient and cost-effective than conventional screening under saline field conditions. Progress in rice breeding for salt tolerance constituted the identification of the major locus conferring a salt tolerance gene at different growth stages.
The current level of genome coverage provided by microsatellite markers in rice is sufficient to be useful for DNA fingerprinting and variety protection (Smith and Helentjaris, 1996) and screening of agronomic performance (Xiao et al., 1996). Microsatellite markers are proving valuable tool for tagging and mapping of salt tolerance genes (Nguyen et al., 2001; Islam, 2004; Niones, 2004). The objectives of the present investigation were screening of rice germplasms under salt stress at the seedling and reproductive stages and evaluation of rice germplasms using SSR markers for salt tolerance.
MATERIALS AND METHODS
Plant materials: Eleven genotypes were investigated for salinity tolerance at seedling and reproductive stages at Bangladesh Institute of Nuclear Agriculture (BINA) glasshouse. Among the test entries, six were Bangladeshi landraces, four were advanced mutants and one was salt tolerant Indian cultivar, Pokkali. Pokkali was used as check to evaluate the salt tolerance of other rice genotypes.
Phenotyping salinity tolerance: IRRI standard protocol (Gregorio et al., 1997) was used to evaluate for salt tolerance of rice germplasms. Salinized and non-salinized setups with three replications were maintained both at seedling and reproductive stages.
Evaluation of rice genotypes at seedling stage: At seedling stage, pregerminated seeds were sown in hydroponic system with tap water. After 3 days, tap water was replaced with nutrient solution. Nutrient solution was prepared following Yoshida et al. (1976). Salinity level was raised to EC 12 dS m-1 and maintained at this level for 30 days and pH value was maintained at 5.5. Nutrient solution was also replaced in every 8 days. The modified Standard Evaluation System (SES) of IRRI was followed to assess the visual symptoms of salt toxicity (Table 1). Initial and final scoring was done at 15 and 21 day after salinization, respectively. Data on plant height and biomass weight were collected.
Evaluation of rice germplasms at reproductive stage: At reproductive stage, perforated pots with fertilized soil were placed in the glassfibre trays, which were served as a water tank. Seedlings were sown in pots and watered with tap water. After 21 days of sowing, tap water was replaced with salinized water for salinized setup and EC was maintained 6 dS m-1. Yield, yield components and biomass weight were collected.
CTAB extraction DNA: Modified CTAB (Cetyl Trimethyl Ammonium Bromide) mini-prep method was followed to extract DNA from leaf samples.
| Table 1: | Modified Standard Evaluation System (SES) of visual salt injury at seedling stage |
 | |
Amplification of microsatellite markers and evaluation of genotypes: Selected nine primers were used for this study as those were used previously by Islam (2004); Bonilla et al. (2002), Niones (2004) and Gregorio et al. (2002) in recombinant inbred lines (RILs) of Pokkali X IR29 for tagging salt tolerance genes, where Pokkali was salt tolerant and IR29 was salt susceptible. Among them RM7075, RM336 and RM253 were polymorphic and showed clear bands and used to reveal polymorphism. Each PCR reaction carried out with 15.0 μL reactions containing 1.5 μL 10 X buffer, 0.75 μL dNTPs, 1 μL primer forward, 1 μL primer reverse, 0.5 μL taq polymerase, 8.25 μL ddH2O and 2.0 μL of each template DNA samples. PCR profile was maintained as initial denaturation at 94°C for 5 min, followed by 34 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min and polymerization at 72°C for 2 min and final extension by 7 min at 72°C. Banding pattern of the genotypes was scored comparing the banding pattern of Pokkali. Germplasms that showed similar banding pattern with Pokkali were considered as tolerant, whereas those with different banding pattern were considered as susceptible.
RESULTS
Screening of genotypes for salt tolerance at seedling stage: Eleven genotypes showed wide variation in phenotypes. Seedlings grown in salinized condition showed different visual symptoms of salt injury. The symptoms were prominent on the first and second leaves and were visualized by leaf rolling, formation of new leaf, brownish and whitish of leaf tip, drying of leaves and also by reduction in root growth, stunted growth and stem thickness leading to complete cessation of growth and dying of seedlings.
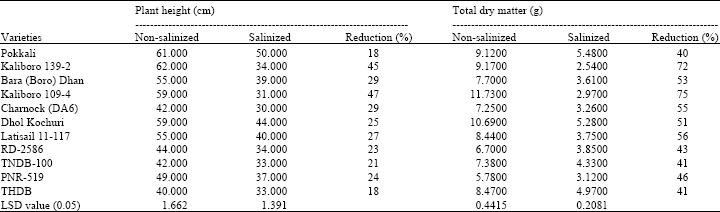
Salt tolerant seedlings were distinguished from the sensitive seedlings grown in salinized condition. The genotypes scored 1 (highly tolerant) to 9 (highly susceptible) according to SES system (Fig. 1). The most salinity tolerant germplasms were Pokkali, THDB and TNDB-100. Four moderately salinity tolerant genotypes were identified as RD-2586, PNR-519, Dhol Kochuri and Bara Dhan.
| Table 2: | Performance of plant height and total dry matter of 11 rice genotypes at seedling stage |
 | |
 | |
| Fig. 1: | Susceptibility of 11 rice genotypes under salt stress at the seedling stage. Genotype: 1 = Pokkali, 2 = Kaliboro 139-2, 3 = Bara (Boro) cDhan, 4 = Kaliboro 109-4, 5 = Charnock (DA6), 6 = Dhol Kochuri, 7 = Latisail 11-117, 8 = RD-2586, 9 = TNDB-100, 10 = PNR-519 and 11 = THDB. Salt tolerance: 1 = highly tolerant and 9 = highly susceptible |
The most susceptible salt tolerant genotypes were Kaliboro 139-2 and Kaliboro 109-4. The modified standard evaluation system (SES) of IRRI (Gregorio et al., 1997) was used in rating the visual symptoms of salt injury.
Plant heights were shorter in salinized condition, compared to the plants grown in non-salinized conditions. Plant height and total dry matter of susceptible genotypes showed higher percent reduction than those of tolerant genotypes. Lower percent reduction of plant height was recorded in genotypes Pokkali and THDB followed by genotypes TNDB-100, RD-2586, PNR-519 and Dhol Kochuri (Table 2). On the other hand, higher percent reduction of plant height was showed by genotypes, Kaliboro 139-2 and Kaliboro 109-4. The percent reduction of total dry matter ranged from 40-75. Lower percent reduction of total dry matter was found in genotypes Pokkali, TNDB-100 and THDB. In contrast, Kaliboro 139-2 and Kaliboro 109-4 showed higher percent reduction of total dry matter.
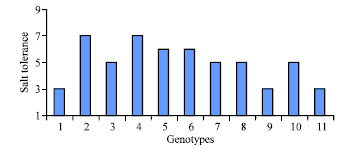
Screening of genotypes for salt tolerance at reproductive stage: Yield and yield components or reproductive stage is more important parameter than that of vegetative or seedling stage tolerance especially where salinity occurs at the reproductive stage. Genotypes RD-2586, TNDB-100, Dhol Kochuri and PNR-519 showed the lowest percent reduction in plant height, total dry matter and number of filled grains at the reproductive stage (Fig. 2). On the other hand, genotypes Kaliboro 139-2 and Kaliboro 109-4 had of the highest percent reductions in the above traits. The genotype RD-2586 showed the lowest reduction in plant height (8.1) followed by TNDB-100, Dhol Kochuri, PNR-519 and Pokkali. The lowest reduction in total dry matter was found for the genotype RD-2586 (24.3) followed by Dhol Kochuri, TNDB-100, Pokkali and PNR-519. Mutant THDB showed the lowest reduction in number of filled grains (11.9) followed by RD-2586, Dhol Kochuri, TNDB-100 and PNR-519.
On the basis of SES score and phenotypic performance, three genotypes, viz. Pokkali, THDB and TNDB-100 were identified as salt tolerant at seedling stage. The genotypes RD 2586, Dhol Kochuri, PNR-519 and Bara (Boro) Dhan were moderately tolerant at the seedling stage, while at the reproductive stage, five genotypes viz. RD 2586, TNDB-100, Dhol Kochuri, PNR-519 and Pokkali were identified as salt tolerant. Finally, the genotypes Pokkali and TNDB-100 showed salt tolerance at both the stages.
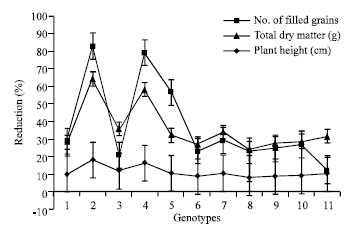
Screening of salt tolerance using SSR markers: Three markers (RM7075, RM336 and RM253) were used to evaluate germplasms for salinity tolerance with reference to Pokkali. Considering the primer RM7075, genotypes Charnock (DA6), Dhol Kochuri, Latisail 11-117, RD-2586, TNDB-100, PNR-519 and THDB were found as tolerant. Where as Kaliboro 139-2, Bara (Boro) Dhan and Kaliboro 109-4 were found as susceptible when the marker RM7075 was used (Fig. 3).
 | |
| Fig. 2: | Percent reduction of plant height, total dry matter and No. of filled grain of 11 rice genotypes at the reproductive stage, when grown in salinized conditions and in relevance to non salinized conditions. 1 = Pokkali, 2 = Kaliboro 139-2, 3 = Bara (Boro) Dhan, 4 = Kaliboro 109-4, 5 = Charnock (DA6), 6 = Dhol Kochuri, 7 = Latisail 11-117, 8 = RD-2586, 9 = TNDB-100, 10 = PNR-519 and 11 = THDB |
 | |
| Fig. 3: | Polymorphism exhibition of primer RM 7075 among 11 rice genotypes. Lane 1 = 1 Kb+ ladder, 2 = Pokkali, 3 = Kaliboro 139-2, 4 = Bara (Boro) Dhan, 5 = Kaliboro 109-4, 6 = Charnock (DA6), 7 = Dhol Kochuri, 8 = Latisail 11-117, 9 = RD-2586, 10 = TNDB-100, 11 = PNR-519 and 12 = THDB |
Bara (Boro) Dhan, Charnock (DA6), Dhol Kochuri, Latisail 11-117, RD-2586, TNDB-100, PNR-519 and THDB were identified as tolerant and Kaliboro 139-2 and Kaliboro 109-4 were identified as susceptible in reaction with RM336, where the findings were almost similar in case of the marker RM7075. Bara (Boro) Dhan, Latisail 11-117, RD-2586, TNDB-100, PNR-519 and THDB were found as tolerant in reaction with RM253 and Kaliboro 139-2, Kaliboro 109-4, Charnock (DA6) and Dhol Kochuri were found as susceptible.
Eight tolerant and three susceptible genotypes were identified using RM7075 with respect to salt tolerant cultivar Pokkali. Out of eight salt tolerant genotypes (in reaction with RM7075), three were tolerant and five were performed as moderately tolerant based on phenotypic performance at the seedling stage, where as five tolerant and three moderately tolerant cultivars were observed at the reproductive stage. Considering RM336, nine genotypes exhibited as salt tolerant and two genotypes were susceptible. Out of nine genotypically identified tolerant genotypes, phenotypically three tolerant and six moderately tolerant were at seedling stage and five tolerant and four moderately tolerant genotypes were identified at reproductive stage. Seven tolerant and four susceptible cultivars were found when genotypes were tested with RM253. Out of these seven salt tolerant genotypes, phenotypically three were tolerant and four moderately tolerant at the seedling stage, whereas at the reproductive stage four were tolerant and three moderately tolerant.
DISCUSSION
Munns et al. (1982) reported that salinity might directly or indirectly inhibit cell division and enlargement in the plant’s growing period. Reduced shoot growth caused by salinity originates in growing tissues, not in mature photosynthetic tissues. As a result, leaves and stems of the affected plants appear stunted. Salinity causes some morphological changes like reduction of shoot (Misra et al., 1995) and root length (Evers et al., 1997) and restriction of rooting (Lopez and Satti, 1996). Peng et al. (1999) reported that increasing plant height would allow greater biomass production. Zhang et al. (2004) supported these findings in their doubled haploid (DH) population which consists of 81 DH lines and they reported that under salt stress, increase of plant height was responsible for increase of biomass; so as to increase yield potential. Islam (2004) found positive and significant correlations among reduction in number of filled grains, percent fertility, grain yield per plant and total biomass weight at reproductive stage in 80 RILs of Pokkali/IR29. The determination of salinity sensitive parameters, for example, thresholds of salinity effects on growth and yield components and interrelationships among yield components under salinity stress, helps developing better management practices for growing rice under salinity and improve understanding of salt tolerance in rice.
Tolerant cultivars showed less growth reduction than sensitive genotypes under salinized conditions (Qian et al., 2001; Suplick-Ploense et al., 2002). Yield components related to final grain yield were severely affected by salinity (Zeng and Shannon, 2000). Salinity affected more the yield and yield components at the reproductive stage, rather than at the vegetative stage as reported by Baloch et al. (2003). In present study considerable effects due to salinity were observed for most of the traits evaluated during the reproductive stage. Salinity significantly reduced the filled grain weight (grain yield), biomass (leaf, shoot and rachis) weight and total biomass weight in RILs of IR29/Pokkali (Salim et al., 1990). The reduction in total biomass weight is the major cause of stand loss in salt affected rice fields. Yield sink capacity is always one of the primary objectives in crop breeding for increasing crop yield. The yield components were believed to have their own critical development periods that can affect final grain yield (Zeng and Shannon, 2000). Salinity response to rice occurs in varying degrees at all growth stages from germination to maturity. Reproductive stage tolerance is a more distinct varietal difference than the vegetative and seedling stage salt tolerance for rice (Mishra et al., 1990).
Genotypes identified as salt tolerant phenotypically were also identified as salt tolerant with markers. Considering phenotypic and genotypic observations, five genotypes RD-2586, TNDB-100, PNR-519, Dhol Kochuri and Pokkali were identified as salt tolerant. Four genotypes viz., THDB, Bara (Boro) Dhan, Latisail 11-117 and Charnock (DA6) were found as moderately tolerant. Bhuiyan (2005) identified 158 tolerant individuals in F2 and F3 populations of BRRI Dhan 28 X PSBRc88 with the marker RM493, with 105 of the individuals being also phenotypically tolerant. Nguyen et al. (2001) found that the marker RM315 was associated with NaCl tolerant alleles under EC 18 dS m-1 and salt stress genes were located in chromosomes 1 and 8. Lang et al. (2000) found that RM223 was closely linked to rice salt tolerance gene in chromosome 8. Foolad (1999) found that only a few major QTLs could account for a large portion of the total phenotypic variation for salt tolerance, indicating that marker-assisted selection for salt tolerance should be feasible, providing the opportunity to rapidly develop germplasms with enhanced salt tolerance at more than one stage of plant development.
Salt stress affected the yield and yields components both at seedling and reproductive stages. Performance in terms of agronomic traits showed lower salinity affection in case of tolerant genotypes than in case of susceptible ones. Phenotypically, Pokkali, Dhol Kochuri, RD-2586, TNDB-100 and PNR-519 showed lower reduction under salt stress as they were found as salt tolerant and these genotypes were also identified tolerant genotypically (with markers). Selected salt tolerant genotypes could be used as a donor for transferring tolerant genes into other desirable cultivars to develop salt tolerant high yielding cultivars. Since, the markers were used in this study showed polymorphism among the genotypes; these markers could be used in tagging salt tolerant genes, in marker-assisted selection and QTL mapping.
ACKNOWLEDGMENT
We would like to express our sincere thanks to International Rice Research Institute (IRRI), Philippines for providing seed materials in this study.
REFERENCES
- Baloch, A.W., A.M. Soomro, M.A. Javed, H.R. Bughio and S.M. Alam et al., 2003. Induction of salt tolerance in rice through mutation breeding. Asian J. Plant Sci., 2: 273-276.
CrossRefDirect Link - Bonilla, P.S., J. Dvorak, D. Mackill, K. Deal and G. Gregorio, 2002. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza saliva L.) using recombinant inbred lines. Philippines Agric. Sci., 85: 64-76.
Direct Link - Evers, D., C. Schmit, Y. Mailliet and J.F. Hausman, 1997. Growth characteristics and biochemical changes of poplar shoots in vitro under sodium chloride stress. J. Plant Physiol., 151: 748-753.
Direct Link - Gregorio, G.B. and D. Senadhira, 1993. Genetic analysis of salinity tolerance in rice (Oryza sativa L.). Theor. Applied Genet., 86: 333-338.
CrossRefDirect Link - Gregorio, G.B., D.R.D. Senadhira, R.D. Mendoza, N.L. Manigbas, J.P. Roxas and C.Q. Guetra, 2002. Progress in breeding for salinity and associated abiotic stresses in rice. Field Crops Res., 76: 91-101.
Direct Link - Lopez, M.V. and S.M.E. Satti, 1996. Calcium and potassium-enhanced growth and yield of tomato under sodium chloride stress. Plant Sci., 114: 19-27.
CrossRefDirect Link - Misra, M., N. Das and A.N. Misra, 1995. Sodium chloride salt stress induced changes in protein content and protease activity in callus culture of pearl millet (Pennisetum glaucum L. R. br.). Acta Physiol. Plant, 17: 371-374.
Direct Link - Peng, S., K.G. Cassman, S.S. Vimani, J. Sheehy and G.S. Khush, 1999. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Sci., 39: 1552-1559.
Direct Link - Xiao, J., J. Li, S.R. McCouch and S.D. Tanksley, 1996. Genetic diversity and its relationship to hybrid performance and heterosis in rice as revealed by PCR-based markers. Theor. Applied Genet., 92: 637-643.
Direct Link - Zeng, L. and M.C. Shannon, 2000. Salinity effects on seedling growth and yield components of rice. Crop Sci., 40: 996-1003.
CrossRefDirect Link - Zhang, Z.H., P. Li, L.X. Wang, Z.L. Hu, L.H. Zhu and Y.G. Zhu, 2004. Genetic dissection of the relationships of biomass production and partitioning with yield and yield related traits in rice. Plant Sci., 167: 1-8.
CrossRefDirect Link