
Samaneh Rahmatzadeh
Department of Biology, Urmia University, Urmia, Iran
Jalil Khara
Department of Biology, Urmia University, Urmia, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 23 | Page No.: 4275-4278







ABSTRACT
UV-C radiation (220-280 nm) is known to causing damage in some physiological growth parameters such as chlorophyll, carotenoid, protein and sugar contents. In this study, effect of some species of vesicular arbuscular mycorrhizal fungi on tolerance of UV-C radiation in wheat plants was studied. Wheat (Triticum aestivum L. cv.Azar2) plants colonized by three species of mycorrhizae namely Glomus etunicatum, Glomus intraradices and Glomus veruciforme were used in this study. They have been exposed to UV-C (254 nm) light for 7 h (28 days, 15 min each day). We measured total proteins, sugars, chlorophyll a and b and carotenoids. Our study showed that UV-C radiation decreases chlorophylls, carotenoids and sugars contents. But this effect on total proteins content has not been significant. However, mycorrhizal fungi could increase all of these factors in comparison to non-mycorrhizal samples. Therefore, these fungi species can increase above growth factors of wheat plants, apparently.
PDF Abstract XML References Citation
How to cite this article
Samaneh Rahmatzadeh and Jalil Khara, 2007. Influence of Ultraviolet-C Radiation on Some Growth Parameters of Mycorrhizal Wheat Plants. Pakistan Journal of Biological Sciences, 10: 4275-4278.
DOI: 10.3923/pjbs.2007.4275.4278
URL: https://scialert.net/abstract/?doi=pjbs.2007.4275.4278
DOI: 10.3923/pjbs.2007.4275.4278
URL: https://scialert.net/abstract/?doi=pjbs.2007.4275.4278
INTRODUCTION
UV-C (λ<280 nm) radiation is one of the solar spectra that affect on metabolic processes in plants. UV radiations are known to inhibit photosynthesis, induce DNA damage as well as causing damage in a wide variety of proteins and lipids (Cockell, 1998).
Arbuscular Mycorrhizae (AM) are beneficial symbionts for plant growth. They are associated with higher plants by a symbiotic association and benefit plants in uptake of phosphorus, production of growth hormones, increase of proteins, lipids and sugar levels, help in heavy metal binding, salinity tolerance (Selvaraj and Padmanabhan, 2006).
The effect of UV-C stress on the growth parameters have been measured in pea and wheat seedlings, in one study. The data indicated that plants response to this stress condition by reducing in chlorophyll content (Alexieva et al., 2001). In another study, seedlings of Taxus cuspidate showed that the content of soluble proteins, carotenoids and chlorophylls have been decreased by UV radiation (Du et al., 2000).
The present investigation was designed to study the effect of mycorrhizal fungi in resistance against UV-C radiation in wheat plants.
MATERIALS AND METHODS
Inoculum production: Pot cultures of the Arbuscular Mycorrhizal Fungi (AMF) Glomus etunicatum, Glomus intraradices and Glomus veruciforme were initiated on corn (Zea mays L.) in a greenhouse during the April to July 2006. Soil used for production of mycorrhizal inoculum was collected from the field and mixed with sand (1:5 w/w) and 100 g fungal inoculum. Soil and sand were autoclaved before mixing at 121°C for 4 h. Plants were grown at 32°C under 18 h light and 8 h dark periods and were illuminated by white fluorescent light and sodium lamp with total irradiance of about 75 μ Em-2 sec-1. Rorison’s solution was used as nutrient medium. Finally, roots were removed from the soil, cut and then mixed with the soil. This culture included soil/sand mixture, hyphae, spores and colonized roots.
Plant treatment: Wheat (Triticum aestivum L. cv. Azar2) seeds were sterilized with 10% sodium hypochlorite solution for 10 min and washed thoroughly with distilled water. Then, seeds were planted in autoclaved soil/sand mixture (1:3 w/w) and 50 g of fungal inoculum in AM treatments. Each treatment was replicated 3 times. Plants were grown at 27°C under 14 h photoperiods and were illuminated by 75 μ Em-2 sec-1. They were exposed to UV-C lamp after 28th day. UV-C radiation was produced by a germicidal lamp (254 nm) that providing an irradiation dose of approximately 40 Wm-2. Plants were grown for 50 days and exposed to UV-C lamp for 7 h (28 days, 15 min each day). The distance between lamps and samples was 50 cm. Plants watered with half strength of Hoagland’s nutrient solution.
Photosynthetic pigments were extracted from leaves in 80% aqueous acetone and content of chlorophylls a, b and carotenoids were determined spectrophotometrically by reading absorbance at 470, 645 and 662 nm as described by Lichtenthaler (1985).
Total protein content in shoot and root were measured by Folen-Lowry method (Lowry et al., 1951). Finally, the total sugar content in roots and shoots systems were determined by Anthrone method described by Fales (1951).
RESULTS
Figure 1 shows the effect of UV-C radiation on total sugar content in shoot of mycorrhizal and non-mycorrhizal wheat plants. UV-C radiation caused a decrease in total sugar of treated samples. But, mycorrhizal plants have higher sugar content than that of non-mycorrhizal samples. Similar results have been observed for total sugars content of the roots (Fig. 2).
Measurement of proteins content in shoots and roots of UV-C treated wheat plants showed that the proteins content decreased under UV treatment. However, this reduction was not significant. These results showed that mycorrhizal inoculated plants had higher levels of proteins in comparison to non fungal samples. Therefore, mycorrhizal fungi can increase proteins content in wheat plants (Fig. 3 and 4).
UV-C radiation and AM fungal inoculation both had significant effects on chlorophylls content. AM fungi generally increased the chlorophylls a, b and carotenoids content and this reduction was higher for chlorophylls b content. However, UV-C radiation treated plants had higher chlorophylls a and b and carotenoids content (Fig. 5, 6 and 7).
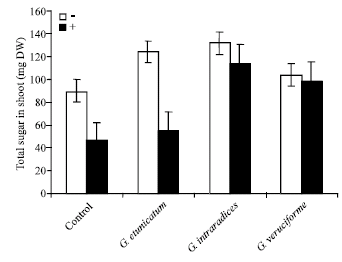 | |
| Fig. 1: | Effect of UV-C on total sugars of shoots in mycorrhizal and non-mycorrhizal wheat plants |
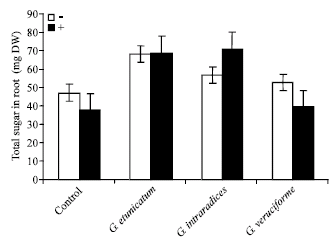 | |
| Fig. 2: | Effect of UV-C on total sugar of roots in mycorrhizal and non-mycorrhizal wheat plants |
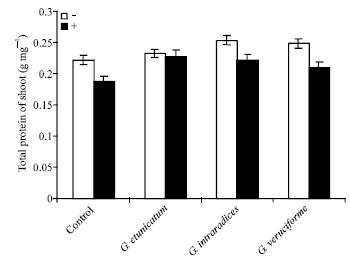 | |
| Fig. 3: | Effect of UV-C on total protein of shoots in mycorrhizal and non-mycorrhizal wheat plants |
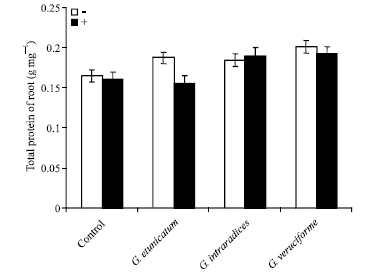 | |
| Fig. 4: | Effect of UV-C on total protein of roots in mycorrhizal and non-mycorrhizal wheat plants |
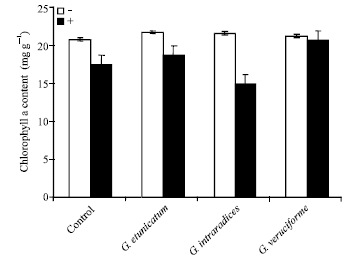 | |
| Fig. 5: | Effect of UV-C on chlorophyll a content in mycorrhizal and non-mycorrhizal wheat plants |
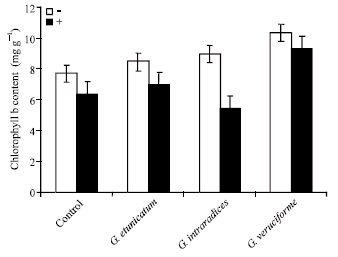 | |
| Fig. 6: | Effect of UV-C on chlorophyll b content in mycorrhizal and non-mycorrhizal wheat plants |
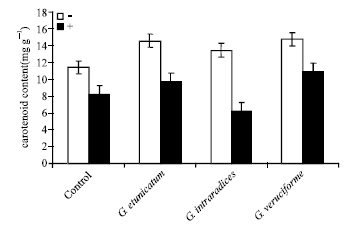 | |
| Fig. 7: | Effect of UV-C on carotenoid content in mycorrhizal and non-mycorrhizal wheat plants |
DISCUSSION
Mycorrhizal fungi are known to modify several aspects of plant physiology, such as mineral nutrient composition, hormonal balance and C allocation patterns (Demir, 2004). In this study, the effects of UV-C radiation on some of the growth parameters of mycorrhizal wheat plants have been investigated. Figure 1 and 2 showed the effect of UV-C radiation on total sugar content of shoots and roots in wheat plants. These results showed that mycorrhizal plants had a higher level of sugar in comparison to non-mycorrhizal samples. Selvaraj (1998) reported that soluble sugars in both leaves and roots had been increased in Glomus fasciculatum inoculated Prosopis juliflora. Mycorrhizal inoculation involves in the increase of reducing sugars in the inoculated plants. In plants, UV-induced structural damage to chloroplasts and to the D1 and D2 proteins of the photosystem II core have been linked to decreases in photosynthetic capacity and growth rate (Teramura, 1996). The organic carbon derived from photosynthesis is transferred to mycorrhizal fungi and this substance maintains the development of spores and fruit bodies in most mycorrhizal types by translocation of the substance to the growing margins of the extraradical mycelium (Smith and Read, 1997). As is known, phosphorous has an important role as an energy carrier during photosynthesis (Jacobsen, 1975). AM fungi increase the uptake of P and they may also increase the synthesis of carbon compounds (Graham, 2000). In one study Du (2003) showed that the PS II electron transport activity in the isolated chloroplasts of Taxus cuspidate needles treated with UV-C, had been decreased. These results indicated that membrane lipid peroxidation of chloroplasts caused by UV-C radiation was the reason of the decrease of PSII electron transport activity and it was caused to decrease in carbohydrate content.
Figure 3 and 4 showed the effect of UV-C on total protein content in mycorrhizal and non-mycorrhizal wheats. These results suggested that mycorrhizal samples had a higher level of proteins in comparison to non-mycorrhizal plants, but this increase was not significant. Absorption of UV radiation by aromatic amino acids and nucleotides can lead to disabling photochemical changes in the corresponding proteins and nucleic acids (Teramura, 1996). Nemec and Meredith (1981) found that Glomus etunicatum inoculated Citrus limon leaves had higher total amino acids than control. Selvaraj (1998) also reported increased levels of total amino acids in Prosopis juliflora inoculated with Glomus fasciculatum. Protein content, increased in arbuscular mycorrhizal inoculated plants, too.
One of the most important indicators of physiological activity is the rate of photosynthesis, which is related to the chlorophyll content of plants (Demir, 2004). Present study indicated that UV-C radiation has been reduced chlorophylls a and b and carotenoids in wheat plants, but this reduction was lower in non-mycorrhizal plants. Investigations showed that the carotenoids and chlorophyll decreased with the increasing time of the UV-C treatment (Du and Jin, 2000). In another study, it has been shown that the carotenoids content decrease after exposure to UV-C light in the snow alga Chlamydomonas nivalis (Duval et al., 2006). In the other hand, it has been shown that the chlorophyll content of mycorrhizal pepper plants was higher than that of non-mycorrhizal control plants in one investigation (Demir, 2004). Also, AM fungi have been shown to increase chlorophyll concentration in the grass Bouteloua gracilis (Allen and Allen, 1981).
Therefore, photosynthesis activity and chlorophyll content increase under UV-C radiation as a result of mycorrhizal association so they have a close relationship with the increase in the function of mycorrhizal fungi.
REFERENCES
- Alexieva, V., I. Sergiev, S. Mapell and E. Karanov, 2001. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ., 24: 1337-1344.
CrossRefDirect Link - Demir, S., 2004. Influence of arbuscular mycorrhiza on some physiological growth parameters of pepper. Turk. J. Biol., 28: 85-90.
Direct Link - Du, Y. and Y. Jin, 2000. Effect of far-ultraviolet radiation on lipid peroxidation and inherent protection system in seedlings of Taxus caspidata. Ying Yong Sheng Tai Xue Bao., 11: 660-664.
Direct Link - Du, Y., P. Jiang, B. Wang and Y. Shi, 2003. Effect of ultraviolet-C irradiation on membrane lipid peroxidation and activity of PSII electron transport in chloroplasts of Taxus cuspidate needles. Ying Yong Sheng Tai Xue Bao., 14: 1218-1222.
PubMedDirect Link - Fales, F.W., 1951. The assimilation and degradation of carbohydrates by yeast cells. J. Biol. Chem., 193: 113-124.
PubMedDirect Link - Lichtenthaler, H.K. and A.R. Wellburn, 1985. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biol. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Selvaraj, T. and C. Padmanabhan, 2006. Arbuscular mycorrhizae: A diverse personality. J. Central Eur. Agric., 7: 349-358.
Direct Link