
Mehmet Karakas
Department of Biology, Faculty of Science and Engineering, Ankara University,
06100 Tandogan, Ankara, Turkey
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4152-4155







ABSTRACT
The life cycle and mating behavior of Zygotylenchus guevarai were observed in vitro on excised roots of Petroselinum crispum in gnotobiotic culture. Eggs hatched into juveniles whose appearance and structure were similar to those of the adults. Juveniles grew in size and each juvenile stage was terminated by a molt. Z. guevarai had four juvenile stages with the first molt occurred outside the egg shortly after hatching. After the final molt the juveniles differentiated into adult males and females. Mating was required for reproduction. After mating, fertilized females began to lay eggs. The life cycle from second stage juvenile to second stage juvenile was completed in 43 days.
PDF Abstract XML References Citation
How to cite this article
Mehmet Karakas, 2007. Life Cycle and Mating Behavior of Zygotylenchus guevarai (Nematoda: Pratylenchidae) on Excised Petroselinum crispum Roots. Pakistan Journal of Biological Sciences, 10: 4152-4155.
DOI: 10.3923/pjbs.2007.4152.4155
URL: https://scialert.net/abstract/?doi=pjbs.2007.4152.4155
DOI: 10.3923/pjbs.2007.4152.4155
URL: https://scialert.net/abstract/?doi=pjbs.2007.4152.4155
INTRODUCTION
One of the most damaging plant parasitic nematodes of the world is the root lesion nematode Zygotylenchus guevarai (Tobar Jiménez, 1963) Braun and Loof, 1966. It has a high damage potential at relatively low population densities and parasitizes a wide range of hosts among agricultural and horticultural crops such as broad bean (Vicia faba), carrot (Daucus carota), celery (Apium graveolens), chick pea (Cicer arietinum), corn (Zea mays), cotton (Gossypium hirsutum), cypress (Cupressus sempervirens), hemp (Cannabis sativa), kidney bean (Phaseolus vulgaris), melon (Cucumis melo), oats (Avena sativa), parsley (Petroselinum crispum), pea (Pisum satinvum), pepper (Capsicum annuum), potato (Solanum tuberosum), tomato (Lycopersicon esculentum) and wheat (Triticum durum) (Siddiqi, 1975; Vovlas et al., 1976; CAB International, 2001). The migratory endoparasitic nematode Z. guevarai invades the root cortex causing root lesions and the formation of large cavities, destroying the cortical tissues (Varo Alcala et al., 1970; Vovlas et al., 1976).
Carrot disc cultures have been used to propagate a number of root lesion nematodes providing large numbers of highly infective nematodes (O’Bannon and Taylor, 1968; Moody et al., 1973; Huettel and Rebois, 1985; Lawn and Noel, 1986; Gugino et al., 2005). Since contamination due to bacteria associated to the carrot tissue sometimes occurs in carrot disc cultures, Agrobacterium rhizogenes transformed potato root cultures were considered as an alternative to propagate Z. guevarai. Transformed root cultures have been successfully used to propagate sedentary endoparasitic nematodes (Paul et al., 1987; Verdejo et al., 1988; Johannes et al., 2005; Han et al., 2006; Jordan and Mitkowski, 2006). The monoxenic culture of Z. guevarai in both carrot and transformed potato root is reported by Verdejo and Pinochet (1991).
In this study, life cycle and mating behavior of Z. guevarai in gnotobiotic culture were investigated.
MATERIALS AND METHODS
Zygotylenchus guevarai was obtained from infested parsley roots in K![]() z
z![]() lcahamam, Turkey (2005-May) and cultured on parsley seedlings in sand-loam soil in the greenhouse (24±2°C and 70% relative humidity). Then a population of Z. guevarai was established in petri dishes on excised parsley roots, Petroselinum crispum (Miller) A.W. Hill., supported by Gamborg’s B5 medium in 1.5% agar adjusted to pH 6.0 and maintained in darkness at 24±2°C (Huang and Becker, 1997).
lcahamam, Turkey (2005-May) and cultured on parsley seedlings in sand-loam soil in the greenhouse (24±2°C and 70% relative humidity). Then a population of Z. guevarai was established in petri dishes on excised parsley roots, Petroselinum crispum (Miller) A.W. Hill., supported by Gamborg’s B5 medium in 1.5% agar adjusted to pH 6.0 and maintained in darkness at 24±2°C (Huang and Becker, 1997).
To study development, nematode eggs from the in vitro culture of Z. guevarai were aseptically transferred onto 1% water agar plates and incubated at 24±2°C overnight. Hatched second stage juveniles (J2) were inoculated onto 10 day old parsley root cultures. Five excised parsley roots were cultured on each of four replicate culture dishes and were inoculated with 100 J2 of Z. guevarai. These plates were incubated under the above conditions. Nematode development and behavior in all replicates were observed daily with dissecting microscope and TZ 240 model Euromex binocular under cold light. The first occurrence of each molt and development stage among the replicates was the criterion used to determine the time periods of the life cycle which reflects a typical time course based on many repeated observations of each event.
Parsley root culture dishes were prepared and divided into three groups with 10 replicates each: (i) one molting female fourth stage juvenile (J4) in each dish, (ii) one molting female J4 and 10 males in each dish, (iii) one molting female J4 and 10 males in each dish with males removed after the first egg appeared in the medium in order to prevent further mating. Molting J4 could be sexually differentiated on the basis of body morphology. After the female J4 finished molting and developed into adults, they were considered virgin females and could only be fertilized by the males added to the same dish. These dishes were incubated as described above and nematode behavior was observed daily for 120 days. Once egg deposition in a dish occurred, the eggs were transferred onto corresponding 1.5% agar dishes and observed for hatching. Hatched juveniles were removed immediately from this agar dish to avoid being mixed with juveniles hatching later. The number of eggs produced by each female was also recorded.
RESULTS AND DISCUSSION
The J2 to J2 life cycle of Z. guevarai was completed under gnotobiotic conditions at 26±2°C in 43 days (Fig. 1). The J2 moved to the root tips and began feeding within 2.0 h after inoculation. Feeding lasted for 12 to 24 h, then the J2 became immobile and remained positioned like a C or a closed circle. The second molt (M2) started 6 days after inoculation. The most significant change during molting occurred in the oesophageal region. During the first 12 to 24 h of molting, the stylet shaft, oesophageal lumen and median bulb became invisible. Only the stylet cone remained discernible.
Twelve hours later, the new cuticle became visible inside the old one, followed by the appearance of the new stylet shaft. Then the oesophageal lumen and the median bulb emerged and gradually became more distinctive. The juvenile body progressively elongated until it was confined by the old cuticle. At this time the new stylet began to probe the old cuticle at the rate of once every 5 to 10 sec, associated with contraction of the median bulb once every 4 to 7 probings. The nematode finally broke through the old cuticle and migrated out. This molting period (M2) lasted for 3 days. The third stage juvenile (J3) began feeding again. At 16 days after inoculation the J3 entered the third molting (M3) period, which lasted for 3 days and resulted in the emergence of the fourth stage juvenile (J4). The J4 started feeding on the roots again, followed by the fourth molting (M4) period. Juveniles that developed into males started the M4 27 days after inoculation.
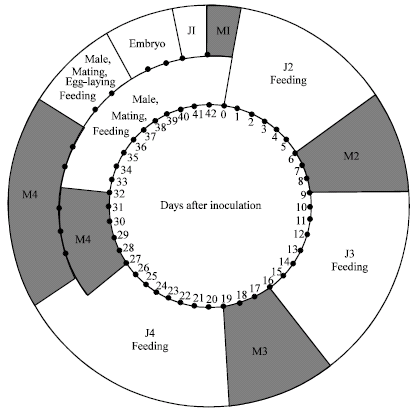 | |
| Fig. 1: | Schematic representation of the life cycle of Zygotylenchus guevarai on excised roots of Petroselinum crispum J1. first-stage juvenile, J2. second-stage juvenile, J3. third-stage juvenile, J4. fourth-stage juvenile, M1. first molt, M2. second molt, M3. third molt, M4 fourth molt |
By the end of the 5 days molting period, the male gonad, the spicules and the caudal alae had formed and the male migrated out of the old cuticle 32 days after inoculation. Juveniles that developed into females started M4 at 28 days after inoculation, which lasted for 7 days. By the end of the molting period, the female gonads and the vulva had formed. The female migrated out of the old cuticle 35 days after inoculation. Faster development of males than females has been observed with other nematodes, e.g., Heterodera schachtii (Johnson and Viglierchio, 1969) and Belonolaimus longicaudatus (Huang et al., 1997) in gnotobiotic cultures.
Males of sting nematode Belonolaimus longicaudatus Rau. approached females soon after the females finished the last molt. Often two or more males surrounded a female, which caused more competition for mates (Perry and Rhodes, 1982; Todd, 1989; Huang et al., 1997). These findings were similar with Z. guevarai. The males seemed to be directly attracted by the females and gathered around them quickly. The males moved around the female and began to intensely rub the side of the female body with the lateral side of their lip region. The rubbing movement of the female head was perpendicular to the axis of the female body. In the meantime one of the males moved toward the female head so that its bursa finally touched the female body. This male would move farther ahead, continuously rubbing the female body until its bursa reached the vulva region of the female. The male then moved back and forth and the female also twisted its body until finally the spicules penetrated through the vulva with the bursa covering the area around the vulva. Then, the body movement of both nematodes slowed down. Mating in this manner lasted for 10-12 min, during which fertilization presumably took place. The male then withdrew its spicules and both nematodes moved away. This mating behavior was observed at least 6 times and each time the mating occurred on the surface of the culturing medium. It was not determined whether a female mated more than once during its life.
After mating, both females and males fed on the surface of the culturing medium. Meanwhile, eggs began forming within female uteri and were clearly visible. Before females began to lay eggs, they stopped feeding and moved slowly within or on the surface of the medium. The eggs in the uterus were pushed toward the vagina. The egg-shall was very flexible and was squeezed to pass the shallow lumen of the vagina and delivered through the vulva. Egg deposition was completed in approximately 5 min, during which the female did not move. The egg resumed its shape outside the female body. The first eggs were laid 38 days after inoculation. All juvenile stages as well as the adult stages of both genders fed on the host.
Zuckerman and Strich-Harari (1963) found no evidence that females of banana spiral nematode could reproduce in the absence of males. The present study allowed examination of this hypothesis in more detail and confirmed that sexual reproduction was obligatory. In the treatment with only one female in each of 10 dishes the females produced in total only 0.5±0.4 eggs during 90 days incubation and none of these eggs hatched. When males were always present in the medium, each female produced an average of 120±15 eggs in 90 days and all the eggs hatched. When the males were removed after the first eggs appeared, each female still produced an average of 128±18 eggs in 90 days and all the eggs hatched. The number of eggs in treatments with fertilized females were not significantly different from each other (Tukey’s test, p = 0.01). Therefore fertilized females produced eggs without the continued presence of males during the 90 day observation. Females collected from field locations almost always have spermathecae filled with sperm (Shahina and Maqbool, 1992b). Under the present culture conditions, each fertilized female produced an average of 2.06±0.26 eggs per day for 90 days. It was not determined how many eggs a female can produce throughout its life time.
In vitro culture of plant nematodes allows continuous observation of the nematodes and has been utilized in nematological studies in the 1950s (Zuckerman, 1971). It has proven to be helpful in studying nematode life cycle and host-parasite relationships. By means of this technique, the life cycle of Heterodera zeae, H. glycines and H. multicinctus have been described in detail (Lauritis et al., 1983a, b; Orion and Bar-Eyal, 1995). However, only a few strictly ectoparasitic phytonematodes such as Criconemella xenoplax (Westcott and Hussey, 1992) and H. multicinctus (Orion and Bar-Eyal, 1995) have been successfully cultured on excised roots. This technique not only provided sterile nematode inocula for well controlled host-nematode relationship studies but also allowed direct observation of the nematode behavior without the interference of soil flora and fauna (Huang and Becker, 1997). Results must be interpreted with caution however, since the metabolic response of the host might be quite from that of an intact host. However, no obvious changes were observed during the course of this study in terms of behavior and parasitism.
REFERENCES
- Han, H.R., W. Dickson and D.P. Weingartner, 2006. Biological characterization of five isolates of Belonolaimus longicaudatus. Nematropica, 36: 25-35.
Direct Link - Huang, X. and O. Becker, 1997. In vitro culture and feeding behavior of Belonolaimus longicaudatus on excised Zea mays roots. J. Nematol., 29: 411-415.
Direct Link - Johannes, H., H. Detlef, B. Jorg and K. Michael, 2005. Plant parasitic nematodes in soil-less culture systems. Nematology, 7: 1-4.
Direct Link - Jordan, K.S. and N.A. Mitkowski, 2006. Population dynamics of plant parasitic nematodes in golf course greens turf in Southern New England. Plant Dis., 90: 501-505.
Direct Link - Lauritis, J.A., R.V. Rebois and L.S. Graney, 1983. Life cycle of Heterodera zeae Koshy, Swarup and Sethi on Zea mays L. axenic roots explants. J. Nematol., 15: 115-119.
PubMedDirect Link - Moody, E.M., B.F. Lownsbery and J.M. Ahmed, 1973. Culture of the root-lesion nematode Pratylenchus vulnus on carrot discs. J. Nematol, 5: 225-226.
PubMedDirect Link - Orion, D. and M. Bar-Eyal, 1995. Observation on the life cycle and behavior of Helicotylenchus multicictus in monoxenic culture on Arabidopsis thaliana. Nematropica, 25: 67-70.
Direct Link - Todd, T.C., 1989. Population dynamics and damage potential of Belonolaimus sp. on corn. Suppl. J. Nematol., 21: 697-702.
PubMedDirect Link - Verdejo, S., B.A. Jaffee and R. Mankau, 1988. Reproduction of Meloidogyne javaniva on plant roots genetically transformed by Agrobacterium rhizogenes. J. Nematol., 20: 599-604.
PubMedDirect Link - Verdejo, S. and J. Pinochet, 1991. Reproduction of the nematode Zygotylenchus guevarai (Nemata: Pratylenchidae) in monoxenic cultures. Revue Nematol., 14: 291-297.
Direct Link - Westcott, S.W. and R.S. Hussey, 1992. Feeding behavior of Criconemella xenoplax in monoxenic culture. Phytopathol., 82: 936-940.
Direct Link - Zuckerman, B.M. and D. Strich-Harari, 1963. The life stages of Helicotylenchus multicinctus (Cobb) in banana roots. Nematologica, 9: 347-353.
CrossRefDirect Link