
Ibtissem Hamrouni-Sellami
Unity of Aromatic and Medicinal Plants, Biotechnological Center, Technopark of Borj-Cedria 2050 Hammam-Lif, Tunisia
Hamadi Ben Salah
Laboratoire des Cultures Industrielles, INRAT, 45 Rue Hedy Karray, 2080 Ariana, Tunisia
Mohamed Elyes Kchouk
Unity of Aromatic and Medicinal Plants, Biotechnological Center, Technopark of Borj-Cedria 2050 Hammam-Lif, Tunisia
Brahim Marzouk
Unity of Aromatic and Medicinal Plants, Biotechnological Center, Technopark of Borj-Cedria 2050 Hammam-Lif, Tunisia
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 21 | Page No.: 3829-3834







ABSTRACT
Phytosterols are bioactive components of all vegetable foods. Their most studied and outstanding properties being their cholesterol-lowering activity. This property has led to the development of functional foods enriched with plant sterols. In the present study, total and conjugated sterols of Tunisian safflower seeds were investigated and their kinetic of accumulation was surveyed during the ripening stages. The results obtained revealed that total and individual phytosterols were actively accumulated during the first stages of seed development then their levels decreased until full maturity. In the other hand, the sterol profile was marked by the predominance of β-sitosterol during all ripening stages. As for Free Sterols (FS) and Esterified Sterols (ES), they were the major components and were actively accumulated from the beginning of seed development. Nevertheless, their amounts decreased at the end of maturity while Steryl Glycosides (SG) and acylated sterylglycosides (ASG) were practically stable during the ripening of seeds.
PDF Abstract XML References Citation
How to cite this article
Ibtissem Hamrouni-Sellami, Hamadi Ben Salah, Mohamed Elyes Kchouk and Brahim Marzouk, 2007. Variations in Phytosterol Composition During the Ripening of Tunisian Safflower (Carthamus tinctorius L.) Seeds. Pakistan Journal of Biological Sciences, 10: 3829-3834.
DOI: 10.3923/pjbs.2007.3829.3834
URL: https://scialert.net/abstract/?doi=pjbs.2007.3829.3834
DOI: 10.3923/pjbs.2007.3829.3834
URL: https://scialert.net/abstract/?doi=pjbs.2007.3829.3834
INTRODUCTION
Plant sterols (Phytosterols) are bioactive components of all vegetable foods. Their structure is similar to that of cholesterol but with some modifications involving the side chain and including the addition of a double bond and/or methyl or ethyl group. These compounds include over 200 different sterols and related compounds in various plant materials (Lagarda et al., 2006), but the more abundant are sitosterol, stigmasterol and campesterol (Moreau et al., 2002; Kritchevsky and Chen, 2005). Sterols in plants exist in the form of Free Alcohols (FS), fatty acid esters (SE), Steryl Glycosides (SG) and Acylated Steryl Glycosides (ASG). In edible oils, phytosterols are mainly present in the free and esterified forms (Phillips et al., 2002; Cunha et al., 2006).
Phytosterols play major roles in several areas, namely in pharmaceuticals (production of therapeutic steroids), nutrition (anti-cholesterol additives in functional foods, anti-cancer properties) and cosmetics (creams, leapstik). These utilisations have enhanced the search of new edible sources of phytosterols. Among the various seed oils, safflower (Carthamus tinctorius L.) could be of great interest since it was mainly studied for its high content of linoleic acid. In this way, proximate analysis of safflower seeds showed that crud protein ranged from 14.9 to 17%, total sugars from 3.2 to 9.2% and extractable lipids from 25 to 40% (Pavlov and Todorov, 1996). Oil content of safflower seeds is similar to the one of olive, sunflower and peanut oils (40% of dry matter weight). Besides, this linoleic oil is composed of 5.5% palmitic (C16:0), 1.6% stearic (C18:0), 11.0% oleic (C18:1), 81.5% linoleic (C18:2) and 0.4% linolenic (C18:3) acids (Lee et al., 2004). In a previous study carried out on safflower seeds from Tunisia, Hamrouni et al. (2004) reported comparable results.
Data on phytosterol composition of safflower seed oil were rather limited in spite of its introduction to the market relatively recently and more particularly, investigation on total sterols and their repartition in free and conjugated forms (ES, SG and ASG) has not yet been done. The objectives of the present study were to develop a simple and validated method for the extraction of total phytosterols (free and conjugated forms) from the seeds of Tunisian safflower. Estimation of total sterol content and their distribution between the free and conjugated forms allowed us to develop these seeds as new possible phytosterol sources. The survey of the kinetic of accumulation of phytosterols during the seed formation and ripening could be of great interest in order to determine the best period for optimal production of these bioactive compounds. The results will be important as an indication of the potentially economical utility of these seeds as a new source of phytosterols.
MATERIALS AND METHODS
Plant material, standards and reagents: Safflower seeds used in this study belonged to the lineage 99B (companion 2002) and were harvested at different ripening stages from the Experimental Station of the National Institute of Agronomic Research of Tunisia located at Oued Béjà in the north-west of the country. Harvest began 2 weeks after anthesis and continued until complete maturity of the seeds marked by the senescence of plants. After harvest, the seeds were stored at 4°C until extraction.
Silica gel plates used for Thin Layer Chromatography (TLC) were 20x20 cm x0.2 mm and were purchased from Merck (G60, Merck, Darmstadt, Germany). BSTFA + TMCS (99:1) used for sylilation of sterols and berberine used for sterol identification were purchased from Fluka (Buchs, Switzerland). Betulin (Lup-20[29]-en-3β, 28diol) used as internal standard for sterol quantification was obtained from Aldrich (St. Louis, Mo., USA). Stigmasterol used for the preparation of standard solution was from Supelco (Bellefonte, PA, USA). Reagents and chemicals used were of analytical grade.
Determination of fresh matter and dry matter weights: After every sampling, 30 seeds were used to determine the Fresh Matter Weight (FMW). Thereafter, these same seeds were dried at 103°C until a constant weight was reached in order to determine their Dry Matter Weight (DMW).
Total sterol extraction: Total sterols of C. tinctorius L. seeds were extracted by soxhlet apparatus according to the method described by Brenac and Sauvaire (1996). For this 10 g of finely ground seed samples were putted in cellulosic extraction cartridges of 30x80 mm. Two successive extractions were realized using soxhlet extractor: the first one for 12 h using hexane and permitting to recover majority of free and esterified sterols, the second one using a mixture of isopropanol-water (70:30; v/v) for 18 h allowing to recover majority of steryl glycosides and acylated steryl glycosides (Brenac and Sauvaire, 1996).
Saponification and unsaponifiable extraction: Two milliliter from each extract (hexane and isopropanol) were transferred into a hydrolize ball and 1 mL of betulinol (Lup-20[29]-en-3β, 28diol) used as internal standard in solution at 0.5 mg mL-1 in the absolute ethanol was added. After evaporation under vacuum, 8 mL of H2SO4 (0.18 N in ethanol 95%) were added and the ball content was heated under ebb during 12 h in order to hydrolize sterol-sugar bond of steryl glycosides and acyl steryl glycosides. To saponify the sterol-fatty acid bond of esterified sterols, the extract was heated under ebb for 90 min after addition of 16 mL of KOH (10% in ethanol 95%). After these two steps (hydrolysis and saponification), the unsaponifiable obtained contained only free sterols coming from the conjugated forms.
After decreasing the alcoholic title of the extract by addition of 100 mL of distilled water, the unsaponifiable was extracted three times with 20 mL of ethylic ether. The extracts obtained were mixed and washed with distilled water until neutrality detected by the addition of some drops of phenolphthalein to the solution recupered after washing.
Preparative TLC and unsaponifiable fractionation: The unsaponifiable was concentrated under vacuum, diluted with 0.5 mL of chloroform and chromatographed on silica gel plates (G60, Merck, Darmstadt, Germany) using a developmental mixture of chloroform-ethylic ether (9:1, v/v). After migration, total sterols and betulinol spots were visualized under UV light at 365 nm after spraying with berberin (0.1% in ethanol 95%). Sterol spots were then scrapped and eluted from silica gel with 2 mL of chloroform.
Derivatization of sterols: Before analysis with GC, all samples were dried under nitrogen stream and derivatized to their trimethylsilylethers (TMS). For this, samples were subjected to 1 h silylation by a BSTFA/TMCS (99:1, v/v) reagent in pyridine in the propotions 1:1 (v/v) at 45°C.
Before GC analysis, samples were evaporated and the residues were dissolved in hexane.
Analysis and identification of total sterols by GC/MS: Sterol samples were analysed by GC/MS using a FID HP 5890 chromatograph and a HP5 capillary column (HP5; Agilent) (25 m, 0.32 nm i.d., 0.52 μm film thickness) with stationary phase made of 5% diphenyl and 95% dimethylpolysiloxane. Helium was used as carrier gas at the flow of 1 mL min-1.
Analyses were performed at split mode under temperature program: isotherm at 150°C during 1 min, from 150 to 300°C at the rate of 6°C min-1, isotherm at 300°C during 12 min. Injector and detector temperatures were respectively of 250 and 280°C.
The GC was coupled to a HP 5972 mass spectrometer operating in the electron-impact mode. The electron energy was set to 70 eV; the source temperature was set to 150°C. Spectra were obtained by electron impact ionization within a mass range of 50-550 m/z (threshold = 150). The scan speed was 1 sec decade-1. Peaks were identified by comparing their mass spectra with those of authentic compounds.
The GC was connected to HP Chemstation (Rev.A. 0401) software for peaks areas and percentages of sterol determination.
Separation of free and conjugated phytosterols by TLC: Free sterols, esterified sterols, sterylglycosides and acyl sterylglycosides were separated by thin layer chromatography according to the method described by Hartmann and Benveniste (1987). For this, 100 μL from hexanic and isopropanolic extracts were mixed and evaporated to dryness. The residue obtained was then diluted into 100 μL of a mixture composed of chloroform/methanol (2:1; v/v) and fractionated using silica gel plates with alumina support (Silicagel G60, Merck) activated during 1 h at 110°C. Plates were submitted to a double development using two solvent combinations: a first one using a mixture of hexane/ethyl acetate (92:8; v/v) and the migration was stopped at 18 cm followed by a second one using a mixture composed of dichloromethane/methanol/water (90:10:0.5; v/v/v) and stopped at 11.7 cm. After spraying with berberin (0.1% in ethanol 95%), sterolic forms were visualized under UV light at 365 nm. Identification of sterolic forms was achieved by co-chromatography of authentic standards in the same conditions.
Quantitative estimation of sterols in the free and in the conjugated forms by spectrophotometry: The method utilized for the quantification of sterols in each sterolic form was inspired from the study of Sabir et al. (2003) based on the estimation of sterols by Liberman-Burchard method: After identification, free sterols and sterol conjugates were scrapped and eluted from silica gel with 2 mL of chloroform. The eluates obtained were then reduced to dryness and additioned of 8 mL of chloroform and they were stirred well until complete dissolution and their absorbance was determined on spectrophotometer after adding 2 mL of reagents. Liberman-Burchard reagent reacts with sterols to produce a characteristic green colour whose absorbance is determined on spectrophotometer (Sherwood 259) at 640 nm.
In order to quantify sterols in each sample, a standard range was prepared from a standard solution of stigmasterol at 1 mg mL-1 in the chloroform. For this, into six tubes marked from 1 to 6 we putted successively 0.5, 1, 1.5, 2 and 2.5 mL of the standard solution whereas tube 6 was kept blank. Then, 2 mL of the Liberman-Burchard reagent was added to all six tubes and the final volume was made equal in all tubes by adding chloroform. The tubes were covered with black carbon paper and kept in dark for 15 min. Then, the absorbance of all standards was determined at 640 nm and standard graph was plotted. Finally, the absorbance of sample solutions was determined after addition of Liberman-Burchard reagent in the same conditions.
Statistical analysis: All analyses were carried out on triplicate and the results reported in figures and tables were mean values with their SDs at 5%.
RESULTS AND DISCUSSION
Phytosterol composition of fully ripe seeds: Sterols comprise the bulk of the unsaponifiable matter in many oils. They are of interest due to their antioxidant activity and impact on health (Dutta et al., 1994). Moreover, analysis of sterols provides a powerful tool for the quality control of vegetable oils and for the detection of oil and mixtures not recognized by the fatty acids profile (Ramadan and Mörsel, 2002).
In fully ripened seeds of C. tinctorius L., TS account for 2.4 mg g-1 of DMW. These concentrations are comparable to those obtained from pumpkin seeds and estimated for 2.15 mg g-1 of DMW (Breinhölder et al., 2002). The results obtained showed that β-sitosterol (4.626 mg g-1 of oil is 51.4% of the TS content) represents the main component followed by Δ5-avenasterol (1.764 mg g-1 of oil = 19.6% of TS content) and campesterol (1.134 mg g-1 of oil = 12.6% of TS content). Stigmasterol, Ergost-7-en-3β-ol, Δ7-avenasterol and a non-identified sterol were detected in small amounts, comprising together 1.476 mg g-1 of oil (16% of TS content). Comparing to safflower seed oil, there are numerous published data on sterols of some other oils of the compositae family, e.g., sunflower (Dutta et al., 1994) and niger (Ramadan and Mörsel, 2002) seeds oils. These oils had higher β-sitosterol content in the range of 50-75% of the TS (Dutta et al., 1994).
| Table 1: | Sterol pattern of safflower seed oils |
 | |
| Data are expressed by mean values±SD of three independent experiments | |
In the other hand, numerous studies carried out on oleaginous seeds reported rates of β-sitosterol similar to present results; this was the case of soybean (Navari-Izzo et al., 1986; Matevienko et al., 2002) with 50% of TSs and lupin (Abdel-Nabey and Shehata, 1991) with 53% of TS.
The fractionation of TS by TLC showed that they were present as free and as conjugated sterols. The later are present in the form of Esterified Sterols (ES), Steryl Glycosides (SG) and Acylated Steryl Glycosides (ASG). As shown in Table 1, C. tinctorius L. sterols were mainly present in the esterified form with 47.8% of TSs (1.099 mg g-1 of DMW) whereas Free Sterols (FS) account for 38.7% of TSs (0.89 mg g-1 of DMW). In the other hand, a minority of sterols were distributed between the SG and the ASG forms representing 7.8% (0.179 mg g-1 of DMW) and 5.7% (0.131 mg g-1 DMW) of TSs respectively. The estimation of total sterol contents in each sterol class of safflower seeds has not yet been done, but other researches carried out on other plant species reported similar results, this was the case of soybean (Katayama and Katoh, 1973) and corn (Davis and Poneleit, 1974) seeds. In addition, in a recent study carried out on niger seed oil, an other plant from the Compositae family, Ramadan and Mörsel (2002) showed that ASG contents exceeded those of SG (ASG/AG = 1.3). However, the same authors found that AG levels were higher than ASG ones in the seeds of coriander, a plant from the Umbelliferae family. These results showed that in some plants SG contents exceeded ASG ones and the contrary is found in some others, but more generally, in all plant species these two moieties (SG and ASG) were detected in few amounts compared to FS and ES.
Sterol evolution during seed ripening: In order to determine the date corresponding to a better qualitative and quantitative yield of phytosterols, we proceeded to the survey of the kinetic of accumulation of these bioactive compounds during the ripening of seeds. The kinetic of TS accumulation presents three stages: in the first stage, comprised between the 18th and the 24th DAA (Days After Anthesis), TSs were actively accumulated in the seeds and reached a maximum of 4.46 mg g-1 DMW on the 24th DAA. Beyond this date, TSs contents decreased significantly and reached a minimum of 2.08 mg g-1 DMW on the 34th DAA. Finally the third stage is characterized by a relative stability of TS levels until the end of maturity (47th DAA) where TS content is of 2.4 mg g-1 DMW (Fig. 1).
Similar kinetics of TS accumulation in maturing seeds was also described.
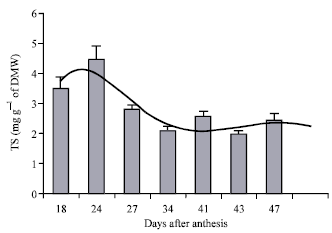 | |
| Fig. 1: | Evolution of the total sterol content during the ripening of safflower seeds. Error bars show the variations of three determinations in terms of SD |
In this way, studying the maturation of soybean seeds, Katayama and Katoh (1973) noticed a decrease of TS content from 4 mg g-1 DMW at the beginning of seed replenishment to 1 mg g-1 DMW at the end of maturity. In the same way, Davis and Poneleit (1974) reported similar results during the ripening of corn seeds. In another investigation on the ripening of pea, Baisted (1971) noticed that sterol biosynthesis is very important during the earlier stages and decreased during seed development. In the other hand, studying the germination of Phaseolus aureus, Geuns (1973) noticed that although all plant tissues were able to biosynthesise sterols, the younger tissues were richer in sterols than the older ones. This author confirm his results by working on hypochotyle sections of Phaseolus aureus (Geuns, 1975) and confirmed that the sterol content of younger sections (1 cm from the apex) is four times greater than the one of older sections (5 cm from the apex). This author suggested that growing tissues biosynthetise continuously biomembranes, which were necessary to cell formation and growth and phytosterols, were very important compounds for the integrity of these biomembranes.
We noticed that during all ripening stages β-sitosterol was the major sterol. Nevertheless, its rates, which were of 78.4 g/100 g TSs on the 18th DAA increased until the 34th DAA and reached 80 g/100 g TSs. After this date, its content decreased significantly until full maturity (47th DAA) where it reached 51 g/100 g TSs. As for the other sterols, the main observations concerned the Δ5-avenasterol whose rates, which were of 2.9 g/100 g TSs on the 18th, DAA, increased extensively and reached 19.6 g/100 g TSs at complete maturity. In the other hand, the contents of campesterol, stigmasterol and Δ7-avenasterol, which were of 6.5, 3.7 and 0.5 g/100 g TSs respectively, increased slightly during the ripening of seeds and reached 13, 6 and 1.3 g/100 g TSs, respectively at the end of maturity.
| Table 2: | Evolution of individual sterols and sterol classes during the ripening of safflower seeds |
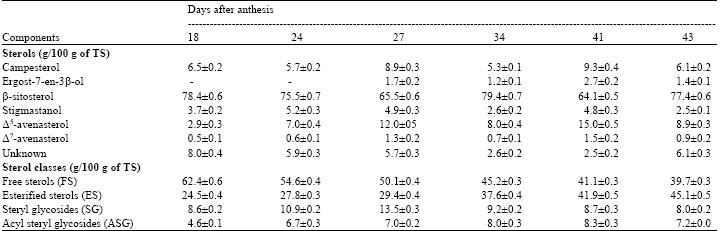 | |
| Data are expressed by mean values±SD of three independent experiments | |
As for the ergost-7-en-3β-ol and the unknown sterol their rates were almost stable around 3 and 5.5 g/100 g TSs respectively until the full maturity of seeds.
The kinetic of sterol evolution during the ripening of safflower seeds is similar to that of soybean seeds (Navari-Izzo et al., 1986).
On the other hand, the distribution of sterols between the free and the conjugated forms during the ripening of safflower seeds was studied and results expressed as gram of sterols in each sterol class per 100 g of total sterols were resumed in Table 2. We noticed that during all development stages there is a predominance of the free and esterified forms (FS and ES). Nevertheless, the level of FS decreased from 62.3 g/100 g TSs in the 18th DAA to 38.7 g/100 g TSs in the end of maturity (47th DAA). The content of ES increased during seed ripening (from 25.4 g/100 g TSs on the 18th DAA to 47.8 g/100 g TSs on the 47th DAA). As for the glycosilated forms (SG and ASG), their contents decreased slightly after the 34th DAA and reached 7.8 g/100 g TSs for SG and 5.7 g/100 g TSs for ASG in fully ripened seeds.
These results are in agreement with those obtained by Katayama and Katoh (1973) in soybean seeds. Bush and Grunwald (1972) suggested that FS may stabilize the structure of biomembranes by interaction with phospholipid. These authors indicated that tissues in active growth must accumulate FS required to the biosynthesis of new biomembranes. Finally, the decrease in the level of glycosilated sterols (SG and ASG) during the ripening of seeds may be attributed to the fact that these forms are precursors for the biosynthesis of many steroids. In this way, Eichenberger (1982) have demonstrated the intervention of cholesterol glycosides during the biosynthesis of steroidal saponins in oat (Avena sativa L.) plants.
CONCLUSIONS
Based on present results, it can be concluded that safflower seeds are an important source of phytosterols mainly of β-sitosterol and Δ5-avenasterol. These results may contribute to the biochemical characterization of safflower seeds and may valorise them as vegetable crops rich in phytosterols that may be utilized for the formulation of functional foods. On the other hand, the survey of the glycosylated forms of sterols (SG and ASG) during the ripening of seeds revealed that the maximum of these compounds corresponded to the earlier stages of seed development. Indeed, these compounds are of great importance for pharmaceutical industries since they could be used as precursors for the synthesis of many hydro-soluble steroids.
ACKNOWLEDGMENTS
The authors are grateful to Prof. Mohamed Hammami and for Imed Cherif from the Unity of Mass Spectrometry of the Faculty of Medicine in Monastir for their help and their cooperation.
REFERENCES
- Abdel-Nabey, A.A. and A.A. Shehata, 1991. Fatty acids, glycerides and unsaponifiables of lupinseed oil. Rivista Italiana delle Sostanze Grasse, 68: 481-485.
Direct Link - Baisted, D.J., 1971. Sterol and triterpene synthesis in the developing and germinating pea seed. Biochem. J., 124: 375-383.
CrossRefDirect Link - Breinholder, P., L. Mosca and W. Lindner, 2002. Concept of sequential analysis of free and conjugated phytosterols in different plant matrices. J. Chromatogr. B, 777: 67-82.
Direct Link - Brenac, P. and Y. Sauvaire, 1996. Accumulation of sterols and steroidal sapogenins in developing fenugreek pods: Possible biosynthesis in situ. Phytochemistry, 41: 415-422.
CrossRefDirect Link - Bush, P.B. and C. Grunwald, 1972. Sterol changes during germination of Nicotiana tabacum seeds. Plant Physiol., 50: 69-72.
CrossRefDirect Link - Cunha, S.S., J.O. Fernandes and M.B.P.P. Oliveira, 2006. Quantification of free and esterified sterols in Portuguese olive oils by solid-phase extraction and gas chromatography-mass spectrometry. J. Chromatogr. A, 1128: 220-227.
Direct Link - Davis, D.L. and C.G. Poneleit, 1974. Sterol accumulation and composition in developing Zea mays L. kernels. Plant Physiol., 54: 794-796.
Direct Link - Dutta, P.C., S. Helmersson, E. Kebedu, G. Alema and L.A. Appelqvist, 1994. Variation in lipid composition of niger seed (Guizotia abyssinica Cass.) samples collected from different regions in Ethiopia. J. Am. Oil Chem. Soc., 71: 839-843.
CrossRefDirect Link - Eichenberger, W., 1982. Incorporation of [4-14C] cholesterol into steryl derivatives and saponins of oat (Avena sativa L.) plants. Plant Cell Rep., 1: 253-256.
CrossRefDirect Link - Geuns, J.M.C., 1973. Variations in sterol composition in etiolated mung bean seedlings. Phytochemistry, 12: 103-106.
CrossRefDirect Link - Geuns, J.M.C., 1975. Regulation of sterol biosynthesis in etiolated mung bean hypocotyl sections. Phytochemistry, 14: 975-978.
CrossRefDirect Link - Hamrouni, I., S. Touati, W. Dhifi, T. Chahed, S. Ayachi, H. Ben Salah and B. Marzouk, 2004. Glycerolipid evolution during safflower seed formation and ripening. J. Food Lipids, 11: 297-311.
CrossRef - Kritchevsky, D. and S.C. Chen, 2005. Phytosterols-health benefits and potential concerns: A review. Nutr. Res., 25: 413-428.
CrossRefDirect Link - Lagarda, M.J., G. Garc�a-Llatas and R. Farre, 2006. Analysis of phytosterols in foods. J. Pharmaceut. Biomed. Anal., 41: 1486-1496.
CrossRefDirect Link - Matevienko, O.A., D.S. Lewis, M. Swanson, B. Arndt, D.L. Rainwater, J. Stewart and L. Alekel, 2002. A single daily dose of soybean phytosterols in ground beef decreases serum total cholesterol and LDL cholesterol in young mildly hypercholesterolemic men. Am. J. Clin. Nutr., 76: 57-64.
Direct Link - Moreau, R.A., B.D. Whitaker and K.B. Hicks, 2002. Phytosterols, phytostanols and their conjugates in foods: Structural diversity, quantitative analysis and health-promoting uses. Prog. Lipid Res., 41: 457-500.
CrossRefPubMedDirect Link - Phillips, K.M., D.M. Ruggio, J.I. Toivo, M.A. Swank and A.H. Simpkins, 2002. Free and esterified sterol composition of edible oils and fats. J. Food Compos. Anal., 15: 123-142.
Direct Link - Ramadan, M.F. and J.T. Moersel, 2002. Proximate neutral lipid composition of niger (Guizotia abyssinica Cass.) seed. Czech J. Food Sci., 20: 98-104.
Direct Link - Sabir, S.M., I. Hayat and S.D.A. Gardezi, 2003. Estimation of sterols in edible fats and oils. Pak. J. Nutr., 2: 178-181.
CrossRefDirect Link