
Mohammad Reza Nassiry
Department of Animal Science, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, P.O. Box 91775-1163, Iran
Alireza Heravi Moussavi
Department of Animal Science, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, P.O. Box 91775-1163, Iran
Abdul Raoof Alashawkany
Department of Animal Science, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, P.O. Box 91775-1163, Iran
Shahrokh Ghovati
Department of Animal Science, College of Agriculture, Ferdowsi University of Mashhad, Mashhad, P.O. Box 91775-1163, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3738-3741







ABSTRACT
The objective of the present investigation was to study genetic variations in the exon II of leptin gene in Iranian Golpayegani and Taleshi cows. Blood samples were collected from 76 Golpayegani and 64 Taleshi cows randomly selected from Delijan city (Esfahan province) and Talesh city (Guilan province) in Iran, respectively. Genomic DNA was extracted from 100 micro litters of blood using guanidium thiocyanate-silica gel method. A 94 bp fragment from exon II of the bovine leptin gene was amplified using the polymerase chain reaction. Digestion of PCR products with Bsp13I restriction enzyme differentiated C and T alleles. The T allele frequencies were 0.29 and 0.45 in Golpayegani and Taleshi breeds, respectively. The frequencies of the genotypes CC, CT and TT were 0.42, 0.58 and 0 in Golpayegani and 0.36, 0.36 and 0.27 in Taleshi cows. The average heterozygosities were 0.41 and 0.50 in Golpayegani and Taleshi breeds, respectively. The χ2 test did not confirm the Hardy-Weinberg equilibrium in both populations.
PDF Abstract XML References Citation
How to cite this article
Mohammad Reza Nassiry, Alireza Heravi Moussavi, Abdul Raoof Alashawkany and Shahrokh Ghovati, 2007. Leptin Gene Polymorphism in Iranian Native Golpayegani and Taleshi Cows. Pakistan Journal of Biological Sciences, 10: 3738-3741.
DOI: 10.3923/pjbs.2007.3738.3741
URL: https://scialert.net/abstract/?doi=pjbs.2007.3738.3741
DOI: 10.3923/pjbs.2007.3738.3741
URL: https://scialert.net/abstract/?doi=pjbs.2007.3738.3741
INTRODUCTION
Economical traits are among quantitative traits that are controlled by many genes each having a small effect (Gelderman, 1997). The major gene model suggests that only a few genes may account for relatively large proportion of the genetic variation (Lande, 1981), such major genes being the genes usually involved in the biology of a trait and are the candidate genes for marker identification. There is also the possibility that major genes may be linked with some Quantitative Trait Loci (QTL) contributing to a major part of the variation in traits (Fruhbeck et al., 1998). One of the candidate genes for marker assistant selection is leptin gene. Leptin is a 16-kDa protein that is synthesized by adipose tissue and is involved in regulation of feed intake, energy balance, fertility and immune functions (Fruhbeck et al., 1998). Plasma leptin levels in cattle and sheep increase linearly with increased body fat mass and with increased energy balance (Blache et al., 2002; Ehrhardt et al., 2000). Leptin gene expressed in a variety of tissues including adipose tissue, placenta, mammary glands, skeletal muscles, gastric mucosa, brain and pituitary glands. It seems that leptin has a large effect in coordinating whole body energy metabolism and may be classified as a metabolism modifier (Houseknecht et al., 1998). It has been shown that leptin gene influences milk performance in cattle (Liefers et al., 2002) and reproduction in beef cattle (Almeida et al., 2003).
In cattle, the leptin gene is located on chromosome 4 and consists of three exons (Pomp et al., 1997). Genetic differences in the leptin gene were first observed in mice ob/ob mice lack functional leptin and are hyperphagic, obese and infertile (Hamann and Matthaei, 1996). Several polymorphisms in this gene have been found in cattle. Restriction fragment length polymorphism of the bovine leptin gene was reported for first time by Lein et al. (1997).
Two RFLP were identified in exon 3: NruI (a C/T substitution resulting in a change from valine to alanine) (Lagonigro et al., 2003) and HphI (a C/T substitution resulting in a change from alanine to valine) (Haegeman et al., 2000). In exon 2, three RFLP were described: ClaI (an A/T substitution resulting in an amino acid change from tyrosine to phenylalanine) (Lagonigro et al., 2003), Kpn2I (Bsp13I) (a C/T substitution resulting in an amino acid change from arginine to cysteine) (Buchanan et al., 2002) and Sau3AI (Pomp et al., 1997). The cysteine/arginine change in exon 2 (Kpn2I) is a non-conserved substitution and is more likely to alter the functioning of the leptin hormone (Buchanan et al., 2002). In livestock, such polymorphisms may be associated with, or linked to, economic traits, which are controlled by many genes each having a small effect (Gelderman, 1997). Leptin is related to both energy metabolism and reproduction and it was shown that leptin polymorphisms had significant effect on calving interval and weight at first calving in beef cows (Almeida et al., 2003) These observations suggest that leptin may be a candidate gene for some of the economically important production traits in dairy cattle. Therefore, the objective of the present investigation was to study the genetic variations in the exon 2 of leptin gene of Iranian native Golpayegani and Taleshi cows.
MATERIALS AND METHODS
Animals and DNA extraction: Blood samples were randomly collected from 76 Golpayegani and 64 Taleshi cattle breeds from Deligan city (Esfahan province) and Talesh city (Guilan province) in Iran, respectively. Genomic DNA was extracted from 100 μL of blood using guanidium thiocyanate-silica gel method (Boom et al., 1990). Quality and quantity of DNA were measured by spectrophotometer by taking the optical density at wavelength of 260 and 280 nm, respectively.
PCR reaction: One microliter of DNA was amplified in a total volume of 25 μL PCR mix using the Biometra T Personal Ver: 1.11 thermocycler. The PCR mix contained: 2.5 μL PCR buffer 10-X (200 mM (NH4)2SO4, 0.1 mM Tween 20%, 750 mM Tris-HCl pH = 8.8), 2.5 mM MgCl2, 200 μM dNTPs and 10 pM from each primer, 1 U Taq DNA polymerase and 11 μL ddH2O. The thermal cycling profile consist of an initial denaturation step of 5 min at 94°C, followed by 35 cycles 45sec at 94°C, 45 sec at 58°C, 45 sec at 72°C and final extension step of 10 min at 72°C. A 94 bp fragment from exon II of the bovine leptin gene was amplified using the following primers described by :
LeptF: 5'ATGCGCTGTGGACCCCTGTATC 3'
LeptR: 5'- TGGTGTCATCCTGGACCTTCC-3'
Products of amplification were recognized by electrophoresis on 1.5% agarose gel stained with etithium bromide.
RFLP: Five micro litters of PCR products were incubated for 5 h at 50°C with three units of Bsp13I restriction enzyme. The C allele was cleaved into two fragments of 75 and 19 bp, while the T allele remained uncut at 94 bp (Buchanan et al., 2002). Digestion products were separated by electrophoresis on non-denaturing 8% polyacrylamide gel stained with AgNO3.
Statistical analysis: The frequencies of genotypes, alleles, mean expected and observed heterozygosities and χ2 test were calculated using PopGene32 (ver 1.31) program [http://cc.oulu.fi/~jaspi/popgen/popdown.htm].
RESULTS AND DISCUSSION
Iranian cattle (Bos indicus and Bos taurus) have unique features like adaptability to extreme climatic conditions, subsistence on poor feed and fodder and better resistance capabilities to withstand environmental stress and tropical disease. There are several diverse cattle breeds in Iran that are primarily being used for drought milk and meat. Although, cattle in Iranian is the most important livestock species and plays a major role in agricultural economy, yet population of some of the important cattle breeds is either declining or breed characters are being diluted under the present production system. To avoid further loss of important gene/gene-pool and preserve maximum amount of genetic diversity, an objective breed classification based on genetic uniqueness is of priority.
This study was done to investigate leptin polymorphism in two Iranian cattle breeds. Present results in leptin polymorphism showed two alleles (C and T) and three genotypes (CC, CT and TT) (Fig. 1) for leptin exon II gene. The CC and CT genotypes were observed in both Golpayegani and Taleshi breeds whereas the TT genotype was observed only in the Taleshi cattle breed. The higher observed frequency for CT genotype was 0.58 in Golpayegani breed (Table 1).
Although the TT genotype was not observed in Golpayegani breed, its frequency was relatively high (0.27) in Taleshi cattle breed. Similar results were reported by Choudhary et al. (2005), who did not detected TT genotype in the Hariana, Sahiwal, Gir and Nimari cattle breeds, but reported comparatively high TT genotype frequency (0.30) in Jersey cattle.
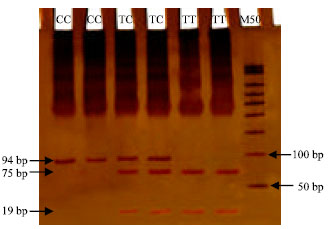 | |
| Fig. 1: | Acrylamide electrophoresis of different leptin genotypes. Molecular marker is M50 (From top to bottom 500, 450, 400, 350, 300, 250, 200, 150, 100 and 50 bp) |
| Table 1: | Observed, expected, average heterozygosities and χ2 test and genotype frequencies of leptin exon 2 for the Iranian native cattle breed |
 | |
| *Significant (p<0.01); **Significant (p<0.05) | |
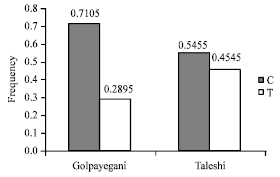 | |
| Fig. 2: | Allelic frequencies of the leptin exon 2 for two Iranian native cattle breed |
For the allelic frequencies, the frequency of T allele was lower than C allele in both cattle breeds (Fig. 2). Present results agree with Buchanan et al. (2002) who had reported C allele frequencies of 0.66 in charolias cattle and 0.68 in Simmental cattle and Madeja et al. (2004) who found frequency of 0.66 in Polish Black and White cattle. Buchanan et al. (2003) reported 0.55 for C allele frequency in Brown Swiss cattle. Present findings for Taleshi cattle (B. Indicus) breed is in contrast to Choudhary et al. (2005), who reported that the T allele found only in B. Taurus cattle breeds. The polymorphisms of leptin gene were reported by several studies. Heravi et al. (2006) found two genotypes, AA and AB, in Irania Holstein cows, which had the frequencies of 0.89 and 0.11, respectively. Madeja et al. (2004) investigated three restriction fragment length polymorphisms (RFLP): HphI, Kpn2I (Bsp13I) and Sau3AI in the leptin gene in Polish Black and White cattle. They reported the allelic frequencies of 0.54 (C) and 0.46 (T) for Kpn2I; 0.66 (C) and 0.34 (T) for HphI and 0.86 (A), 0.11 (B) and 0.03 (C) for Sau3AI. Konfortov et al. (1999) using sequence analysis detected 20 polymorphisms in both introns and exons of leptin gene in a diverse panel of cattle. Liefers et al. (2003) studied three microsatellite markers (R4C, RFLP1, A59V and BM1500) in leptin gene. Lagonigro et al. (2003) reported a new non-conservative mutation in exon II of the leptin gene. They used five different breeds and the frequency of the rare allele ranged from 0 to 15.2%. Pomp et al. (1997) using PCR-based polymorphism showed a rare polymorphism in the leptin gene. They reported that the Sau3AI restriction enzyme digested the PCR product four times instead of three times. These cattles are excellent sources of biological information for studies on genetic characterization in Iran. These breeds showed a middle degree of genetic variability and deviation from H-W equilibrium for the leptin locus (Table 1); this might be explained by breeding method, which is used in these stations. With respect to low number of population, the inbreeding rate is high so heterozygosity and genetic variability is not high in the studied populations.
REFERENCES
- Almeida, S.E., E.A. Almeida, J.C.F. Moraes and T. Weimer, 2003. Molecular markers in the LEP gene and reproductive performance of beef cattle. J. Anim. Breed. Genet., 120: 106-113.
CrossRefDirect Link - Blache, D., R.L. Tellam, L.M. Chagas, M.A. Blackberry, P.E. Vercoe and G.B. Martin, 2000. Level of nutrition affects leptin concentrations in plasma and cerebrospinal fluid in sheep. J. Endocrinol., 165: 625-637.
CrossRefDirect Link - Boom, R., C.J. Sol, M.M. Salimans, C.L. Jansen, P.M.E. Wertheim-van Dillen and J. van der Noordaa, 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol., 28: 495-503.
PubMedDirect Link - Buchanan, F.C., C.J. Fitzsimmons, A.G. van Kessel, T.D. Thue, D.C.W. Sim and S.M. Schmutz, 2002. Association of a missense mutation in the bovine leptin gene with carcass fat content and leptin mRNA levels. Genet. Sel. Evol., 34: 105-116.
CrossRefPubMedDirect Link - Buchanan, F.C., A.G. van Kessel, C. Waldner, D.A. Christensen, B. Laarveld and S.M. Schmutz, 2003. An association between a leptin single nucleotide polymorphism and milk and protein yield. J. Dairy Sci., 86: 3164-3166.
CrossRefDirect Link - Choudhary, V., P. Kumar, T.K. Bhattacharya, B. Bhushan and A. Sharma, 2005. DNA polymorphism of leptin gene in Bos indicus and Bos taurus cattle. J. Genet. Mol. Biol., 28: 740-742.
Direct Link - Ehrhardt, R.A., R.M. Slepetis, J. Siegal-Willott, M.E. van Amburgh, A.W. Bell and Y.R. Boisclair, 2000. Development of a specific radioimmunoassay to measure physiological changes of circulating leptin in cattle and sheep. J. Endocrinol., 166: 519-528.
CrossRefDirect Link - Fruhbeck, G., S.A. Jebb and A.M. Prentice, 1998. Leptin: Physiology and pathophysiology. J. Clin. Physiol., 18: 399-419.
Direct Link - Haegeman, A., A. van Zeveren and L.J. Peelman, 2000. New mutation in exon 2 of bovine leptin gene. Anim. Genet., 31: 79-79.
CrossRefDirect Link - Hamann, A. and S. Matthaei, 1996. Regulation of energy balance by leptin. Exp. Clin. Endocrinol. Diabetes, 104: 293-300.
Direct Link - Moussavi, A.H., M. Ahouei, M.R., Nassiry and A. Javadmanesh, 2006. Association of leptin polymorphism with production, reproduction and plasma glucose level in Iranian holstein cows. Asian-Aust. J. Anim. Sci., 19: 627-631.
Direct Link - Houseknecht, K.L., C.A. Baile, R.L. Matteri and M.E. Spurlock, 1998. The biology of leptin: A review. J. Anim. Sci., 76: 1405-1420.
PubMedDirect Link - Konfortov, B.A., V.E. Licence and J.R. Miller, 1999. Re-sequencing of DNA from a diverse panel of cattle reveals a high level of polymorphism in both intron and exon. Mamm. Genome, 10: 1142-1145.
Direct Link - Lagonigro, R., P. Wiener, F. Pilla, J.A. Woolliams and J.L. Williams, 2003. A new mutation in the coding region of the bovine leptin gene associated with feed intake. Anim. Genet., 34: 371-374.
CrossRefDirect Link - Liefers, S.C., M.F.W. te Pas, R.F. Veerkamp and T. van der Lende, 2002. Associations between leptin gene polymorphisms and production, live weight, energy balance, feed intake and fertility in holstein heifers. J. Dairy Sci., 85: 1633-1638.
CrossRefDirect Link - Liefers, S.C., R.F. Veerkamp, M.F.W. te Pas, C. Delavaud, Y. Chilliard and T. van der lende, 2003. Leptin concentrations in relation to energy balance, milk yield, intake, live weight and estrus in dairy cows. J. Dairy Sci., 86: 799-807.
CrossRefDirect Link - Madeja, Z., T. Adamowicz, A. Chmurzynska, T. Jankowski, J. Melonek, M. Switonski and T. Strabel, 2004. Short communication effect of leptin gene polymorphisms on breeding value for milk production traits. J. Dairy Sci., 87: 3925-3927.
Direct Link - Pomp, D., T. Zou, A.C. Clutter and W. Barendse, 1997. Rapid communication mapping of leptin to bovine chromosome 4 by linkage analysis of a PCR-based polymorphism. J. Anim. Sci., 75: 1427-1427.
PubMedDirect Link