
Intan Ahmad
Physiology, Developmental Biology and Biomedical Research Group, School of Life Sciences and Technology, Bandung Institute of Technology, Jalan Ganesha No. 10, Bandung 40132, Indonesia
Sita Astari
Physiology, Developmental Biology and Biomedical Research Group, School of Life Sciences and Technology, Bandung Institute of Technology, Jalan Ganesha No. 10, Bandung 40132, Indonesia
Marselina Tan
Physiology, Developmental Biology and Biomedical Research Group, School of Life Sciences and Technology, Bandung Institute of Technology, Jalan Ganesha No. 10, Bandung 40132, Indonesia
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3688-3692







ABSTRACT
Three strains of Aedes aegypti collected in 2006 from three major cities in Indonesia, i.e., Bandung, Palembang and Surabaya, were tested to determine their resistance status to pyrethroid (permethrin and deltamethrin) and also the resistance mechanisms regarding three detoxifying enzymes, i.e., oxidase, esterase A and esterase B. The resistance level was expressed as Resistance Ratio (RR) compared to the susceptible VCRU strain. Results of this study showed that Ae. aegypti in Indonesia has developed tolerance to permethrin and deltamethrin, except in the Bandung strain, which was resistant to permethrin and deltametherin with RR90 79.3 and 23.7, respectively. The study also suggested that detoxifying enzymes (oxidase, esterase A and esterase B) apparently were involved in the development of resistance in Bandung strain as indicated by the high level of activity of those enzymes in Bandung strain compared to other more susceptible strains. Although there is a possibility that other mechanisms, such as a target-site resistance mechanism, in Bandung strain were also involved.
PDF Abstract XML References Citation
How to cite this article
Intan Ahmad, Sita Astari and Marselina Tan, 2007. Resistance of Aedes aegypti (Diptera: Culicidae) in 2006 to Pyrethroid Insecticides in Indonesia and its Association with Oxidase and Esterase Levels. Pakistan Journal of Biological Sciences, 10: 3688-3692.
DOI: 10.3923/pjbs.2007.3688.3692
URL: https://scialert.net/abstract/?doi=pjbs.2007.3688.3692
DOI: 10.3923/pjbs.2007.3688.3692
URL: https://scialert.net/abstract/?doi=pjbs.2007.3688.3692
INTRODUCTION
Aedes aegypti is a primary vector for Dengue Fever/Dengue Haemorrhagic Fever (DF/DHF) virus, one of the major public health problems in many subtropical and tropical countries. For example, in Indonesia, a tropical country, several outbreaks of dengue have occurred since its first occurrence in 1968 and recently in 2006, 11173 cases with 1152 deaths were reported by the Ministry of Health Indonesia. In an effort to prevent or reduce dengue transmission, insecticides are used regularly to control Ae. aegypti. However the intensive and continuous use of insecticides to control this mosquito has caused the development of resistance of Ae. aegypti to many insecticides (Ponlawat et al., 2005; Macoris et al., 2003) and hinders the control programs (WHO, 1986). In Indonesia, organophosphates (temephos and malathion) and pyrethroids (permethrin and deltamethrin) have been widely used to control mosquito vectors, respectively since 1970s and 1980s. It is unfortunate that information on the susceptibility or resistance of Ae. aegypti to insecticides in Indonesia is limited. Among the few studies, Arief (2000) and Astari and Ahmad (2005) showed that Ae. aegypti collected in Bandung and several other cities in West Java, Indonesia (Ciamis, Purwakarta and Bogor) have become resistant to organophosphate and pyrethroids (deltamethrin and permethrin), unfortunately, however, their studies did not use a standard susceptible strain as control, resulting in the uncertainty of the resistant status. Whilst Brengues et al. (2003) reported a very high level of resistance to permethrin in Ae. aegypti collected from Semarang West Java Indonesia.
There are many reports detailing the role of metabolism in pyrethroid resistance in insects. For example, Vulule et al. (1999) reported elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae, whereas Brengues et al. (2003) found that Indonesian strain Ae. aegypti collected from Semarang and Salatiga had elevated carboxyl esterase and monooxygenase levels associated with permethrin resistance.
This report describes the current resistance status of Ae. aegypti collected from three cities in Indonesia, i.e., Bandung (West Java), Palembang (South Sumatera) and Surabaya (East Java), to pyrethroids (permethrin and deltamethrin), as well as the possible mechanisms, which include the detoxifying enzymes, i.e., oxidase and esterase. However, different than previous workers, including Vulule et al. (1999) and Brengues et al. (2003), in the study reported here, we compare the activity levels of oxidase and esterase in four strains of Ae. aegypti before and after exposure to permethrin and deltamethrin.
MATERIALS AND METHODS
Mosquitoes: Three mosquito strains were tested in this study. They were collected from three cities located in three provinces in Indonesia in 2006, i.e., Bandung-West Java (BDG), Palembang -South Sumatera (PAL) and Surabaya-East Java (SBY). A pure susceptible Vector Control Research Unit (VCRU) strain was generously provided by Vector Control Research Unit, Universiti Sains Malaysia and was used as standard strain. All strains were reared in the Laboratory of Entomology, School of Life Sciences and Technology, Bandung Institute of Technology, before being used.
Insecticides: Insecticides used were permethrin 0.75% and deltamethrin 0.05%. Permethrin and deltamethrin were obtained from WHO in the form of insecticide impregnated paper.
Chemicals: Chemical substances used for biochemical assay were, among others, cytochrome-C, 3,3’,5,5’-tetramethylbenzidine, α-naphthyl acetate, β-naphthyl acetate and O-Dianisidine (Fast Blue B) [Sigma-Aldrich, St. Louis, MO].
Resistance assay: Assay to determine resistance status of Ae. aegypti to permethrin and deltamethrin was conducted using method from WHO. Six groups of 15-25 adult male and female mosquitoes (2 days old) were exposed to insecticides for 5, 10, 15, 30, 45 and 60 min. The mosquitoes then were allowed to recover and after 24 h the number of dead mosquitoes was counted. The result was analyzed using probit analysis to determine LT90. The resistance level was determined by comparing the LT90 values of mosquito tested to the LT90 values of VCRU strain.
Biochemical assay: Biochemical assay was conducted using two samples of mosquito, i.e., mosquitoes not exposed (control) and exposed for 60 min to permethrin and deltamethrin before treatment. The mosquitoes used were adult male and female mosquitoes (2 days old). Exposure time was based on maximum exposure time in resistance assay.
Total protein content was determined using the Bradford method. Individual mosquitoes were homogenized in 500 μL potassium phosphate buffer (PPB) 0.1 M pH 7.2 using pestle in 1.5 mL microtube. The homogenate was centrifuged at 13,000 rpm in 4°C for 10 min. The supernatant was stored in -80°C before use. To determine the total protein content, 25 and 50 μL from each mosquito sample were assayed and the results were compared to a standard curve using bovine serum albumin as protein standard.
Assay to determine the activity levels of oxidase, esterase A and esterase B were conducted using a method as described on the Center for Disease Control website (http://www.cdc.gov/ncidod/wbt/resistance/ assay/microplate/index.htm) with modification. Samples containing 5 μg protein was used in each assay. In oxidase assay, TMBZ and H2O2 3% were added to the sample and the mixture was incubated for 5 min. Then, H2SO4 2 M was added as stop solution and the results were measured using Microplate Reader Spectrophotometer Model 550 (Biorad) at 450 nm. The results were compared to a standard curve of cytochrome-C. Activity level of oxidase was expressed in Unit/mg protein.
In esterase assay, α and β-naphthyl acetate were used as substrate for esterase A and esterase B, respectively. The mixture was incubated for 30 min and then O-dianisidine and SDS 1% were added as dye agent and stop solution, respectively. The resulted color can be measured using microplate reader spectrophotometer at 570 nm. The results were compared to standard curves of α and β-naphthol, the product of α and β-naphthyl acetate hydrolysis by esterase A and B, respectively. Activity levels of esterase A and esterase B were expressed in μmol-product/min.mg protein.
RESULTS AND DISCUSSION
Result of the assay showed that Bandung-West Java strain was the most resistant strain. It was resistant to permethrin and deltamethrin with RR90 79.3 and 23.7, respectively (Table 1). Palembang-South Sumatera strain was slightly resistant to permethrin (RR90 11.1) but still susceptible to deltamethrin (RR90 2.2). Surabaya-East Java strain was still susceptible to permethrin and deltamethrin (RR90 8.6 and 2.5, respectively). The definition of resistance was based on Valles et al. (1997), i.e., that a population is considered resistant if RR is more than 10. However, although Surabaya-East Java strain was considered susceptible to permethrin and deltamethrin and Palembang-South Sumatera strain was considered susceptible to deltamethrin, the higher RR compared to VCRU strain suggested that those strains have already developed tolerance to permethrin and deltamethrin and may develop resistance in the future. This is likely to happen if pyrethroids keep being used to control the population, thus allowing the resistant individuals to survive and reproduce.
| Table 1: | Resistance levels of Ae. aegypti to permethrin and deltamethrin |
 | |
| VCRU: Vector Control Research Unit, a susceptible pure strain of Ae. aegypti, PAL: Palembang, SBY: Surabaya, BDG: Bandung, LT90: Lethal Time 90%, min, RR90: Resistance Ratio at LT90 | |
Many studies have shown that resistance level of a population can increase in several generations when selection using insecticides was applied to the population (Chareonviriyaphap et al., 2002; Leksono, 1997).
The result of this study appears consonant with that of Brengues et al. (2003) interestingly however, reported a very high resistance ratio (296-fold) occurring in Semarang strain, whereas in the present study, the highest resistance ratio was only 79.3-fold, occurring in Bandung Strain. The reason for this difference resistance levels is probably due to the different frequency of insecticides use between those areas.
Resistance of Bandung-West Java strain to permethrin and deltamethrin may be cross resistance, which is resistance to different insecticides that have similar structure or mode of action. Permethrin and deltamethrin differ mainly by the α-cyano in deltamethrin that is the characteristics of type II pyrethroids. The α-cyano gives steric effect on deltamethrin that causes deltamethrin to be more stable and difficult to be hydrolyzed on its ester bond (Kerkut and Gilbert, 1985).
Cross resistance to several types of pyrethroid insecticides have been found in many mosquito species from many countries, for example An. stephensi and An. culicifacies from India (Ganesh et al., 2003). However, there were no indications of cross resistance in other strains, because of the low levels of resistance in those strains to permethrin as well as deltamethrin. Resistance of Bandung strain to permethrin and deltamethrin may be due partly to the increased level of activity of detoxifying enzymes. Many studies have found that elevated levels of activity of those enzymes (oxidases, esterase A and esterase B) can be related to an increase in tolerance to insecticides in mosquitoes, suggesting the role of detoxifying enzymes in the development of resistance to pyrethroids and organophosphate insecticides in mosquitoes (Chareonviriyaphap et al., 2003; Macoris et al., 2003; Yaicharoen et al., 2005). The results of biochemical assay in control mosquitoes (not exposed to insecticides) showed that activity levels of detoxifying enzymes (oxidases, esterase A and esterase B) before exposure to insecticides were significantly higher in Bandung strain compared to the susceptible VCRU strain (Table 2-4), suggesting that those enzymes might be involved in the development of resistance in Bandung strain.
| Table 2: | Activity levels of oxidase of four strains of Ae. aegypti before and after exposure to permethrin and deltamethrin |
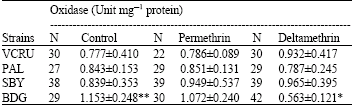 | |
| *: Significantly different (p<0.05) from control (not exposed) of the same strain, **: Significantly different (p<0.05) from VCRU in the same column, N: Number of mosquitoes tested | |
| Table 3: | Activity levels of esterase A of four strains of Ae. aegypti before and after exposure to permethrin and deltamethrin |
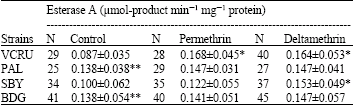 | |
| *: Significantly different (p<0.05) from control (not exposed) of the same strain, **: Significantly different (p<0.05) from VCRU in the same column, N: Number of mosquitoes tested | |
| Table 4: | Activity levels of esterase B of four strains of Ae. aegypti before and after exposure to permethrin and deltamethrin |
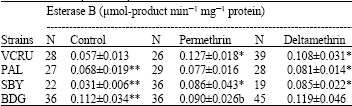 | |
| *: Significantly different (p<0.05) from control (not exposed) of the same strain, **: Significantly different (p<0.05) from VCRU in the same column, N: Number of mosquitoes tested | |
Both oxidase and esterase are capable of detoxifying pyrethroids through different mechanisms. Esterases detoxify pyrethroids through hydrolysis of ester bond, while oxidases detoxify pyrethroids through hydroxylation (adding -OH) or oxidation of ester bonds of pyrethroids (Kerkut and Gilbert, 1985). The high level of resistance in Bandung strain presumably was caused by a combination of various detoxifying enzymes available in the insect’s body, which together can readily detoxify the insecticides entering the insect’s body before reaching their target site.
Exposure to permethrin and deltamethrin caused a change in the activity levels of the enzymes, as can be seen in Table 2-4. Exposure to permethrin and deltamethrin increased the activity levels of the enzymes in all strains, except for oxidase in the Bandung strain, which decreased after exposure to deltamethrin. Overall, increase in the more resistant strain (Bandung) was lower than the more susceptible strain (VCRU, Surabaya and Palembang). It is hypothesized that some of the insecticides entering the insect’s body can be detoxified directly by the detoxifying enzymes, while the rest are bound by receptors and induce the production of more enzymes. In Bandung strain, it is probable that because the activity level of the enzymes were higher than in other strains, more insecticides could be detoxified and less insecticides were available to induce the production of the enzymes. A plausible explanation is that in Bandung strain, another mechanism of pyrethroid resistance is involved, i.e., changes in target site sensitivity. For example, Brengues et al. (2003) reported that in a highly resistant Ae aegypti strain from Semarang (269-fold levels of resistance to permethrin), the activity level of esterases and monooxygenases were not significantly higher than those of controls.
In conclusions the results of this research suggested that the resistance level of Ae. aegypti to pyrethroids (permethrin and deltamethrin) in Indonesia is still low, except for Bandung strain. However, the other strains tested already showed increased tolerance to both insecticides, indicating that a portion of the population has already developed resistance to the insecticides. If treatment using insecticides is still applied continuously, there is the possibility that those populations will become resistant in the future. Moreover, the involvement of oxidase and esterase and possibly changes in target sensitivity in the development of resistance, especially in Bandung strain, may pose a threat to the possibility of cross resistance to other classes of insecticides, such as temephos (organophosphate), which is widely used in Indonesia to control mosquitoes. Therefore, continuous monitoring of resistance levels of mosquito to insecticides is very important and must be done to prevent the development of resistance in the future that consequently might lead to more severe DF/DHF outbreaks.
ACKNOWLEDGMENTS
This research was funded by a Bandung Institute of Technology Research Grant and a Competitive Research Grant from Directorate General of Higher Education Given to Intan Ahmad. We also would like to thank Dr. Zairi Jaal of Vector Control Research Unit, University Sains Malaysia for providing susceptible mosquito eggs and to Dr. Amrul Munif for providing mosquito eggs collected from South Sumatera and East Java. We also thank Dr. Devi Choesin for helpful comments on the manuscript.
REFERENCES
- Astari, S. and I. Ahmad, 2005. Insecticide resistance and effect of piperonyl butoxide as a synergist in three strains of Aedes aegypti (Linn.) (Diptera: Culicidae) against insecticides permethrin, cypermethrin and D-allethri. Buletin Penelitian Kesehatan, 33: 73-79.
Direct Link - Brengues, C., N.J. Hawkes, F. Chandre, L. McCarroll and S. Duchon et al., 2003. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol., 17: 87-94.
CrossRefPubMedDirect Link - Chareonviriyaphap, T., P. Rongnoparut and P. Juntarumporn, 2002. Selection for pyrethroid resistance in a colony of Anopheles minimus species A, a malaria vector in Thailand. J. Vector Ecol., 27: 222-229.
Direct Link - Chareonviriyaphap, T., P. Rongnoparut, P. Chantarumporn and M.J. Bangs, 2003. Biochemical detection of pyrethroid resistance mechanisms in Anopheles minimus in Thailand. J. Vector Ecol., 28: 108-116.
Direct Link - Ganesh, K.N., J. Urmila, P. Guillet and L. Manga, 2003. Pyrethroid susceptibility and enzyme activity in two malaria vectors, Anopheles stephensi (Liston) and A. culicifacies (Giles) from Mysore, India. Ind. J. Med. Res., 117: 30-38.
Direct Link - Macoris, M.L.G., M.T.M. Andrighetti, L. Takaku, C.M. Glasser, V.V. Garbeloto and J.E. Bracco, 2003. Resistance of Aedes aegypti from Sao Paolo Brazil to organophosphate insecticides. Mem. Inst. Oswaldo Cruz., 98: 703-708.
Direct Link - Ponlawat, A., J.G. Scott and L.C. Harrington, 2005. Insecticides susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J. Med. Entomol., 42: 821-825.
Direct Link - Valles, S.M., P.G. Koehler and R.J. Brenner, 1997. Antagonism of fipronil toxicity by Piperonyl butoxide and S,S,S-tributyl phosphorotrithioate in the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol., 90: 1254-1258.
Direct Link - Vulule, J.M., R.F. Beach, F.K. Atieli, J.C. McAllister and W.G. Brogdon et al., 1999. Elevated oxidase and esterase levels associated with permethrin Tolerance in Anopheles gambiae from Kenyan villages using permethrin-impregnated nets. Med. Vet. Entomol., 13: 239-244.
Direct Link - Yaicharoen, R., R. Kiatfuengfoo, T. Chareonviriyaphap and P. Rongnoparut, 2005. Characterization of deltamethrin resistance in field populations of Aedes aegypti in Thailand. J. Vector Ecol., 30: 144-150.
Direct Link