
S. D. Siadat
Department of Bacterial Vaccines and Antigen Production, Pasteur Institute of Iran, Tehran, Iran
M. Kheirandish
Research Center of Iranian Blood Transfusion Organization, Tehran, Iran
D. Norouzian
Department of Bacterial Vaccines and Antigen Production, Pasteur Institute of Iran, Tehran, Iran
Q. Behzadiyannejad
Faculty of Medicine, Tarbiat Modarres University, Tehran, Iran
S. Najar Peerayeh
Faculty of Medicine, Tarbiat Modarres University, Tehran, Iran
M. Zangeneh
Faculty of Medicine, Islamic Azad University, Tehran, Iran
M. Nejati
Department of Bacterial Vaccines and Antigen Production, Pasteur Institute of Iran, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3578-3584







ABSTRACT
Opsonophagocytosis mediated by antibody and complement is the major defense mechanism for clearing Neisseria meningitidis from the host. Therefore, a newly developed phagocytosis assay based on flow cytometry (flow assay) was using sera obtained from rabbit postvaccination with outer membrane vesicle of N. meningitidis serogroup B, was done in order to evaluation of the potential efficacy of (experimental) meningococcal vaccines. The Outer Membrane Vesicles (OMVs) and control were injected intramuscularly into groups of five rabbit with boosters on 14, 28 and 42 days after the primary immunization. The serum on 0, 14, 28, 42 and 56 days were collected and stored at -20°C for next analysis. Phagocytic function of and intracellular oxidative burst generation by rabbit polymorphonuclear (PMN), against N. meningitidis serogroup B, was measured with flow cytometer, using dihydrorhodamine-123 as probes, respectively. We use a Coulter Epics XL-profile (USA) with an argon laser operating at 488 nm. The results of quantitative flow cytometric analysis of rabbit PMN function in hyperimmun sera with OMVs revealed a highly significant increase in opsonophagocytic responses against serogroup B meningococci after 56 day in comparison with the control group (p<0.05). Present results indicated that OMVs could be as a candidate for vaccine toward serogroup B meningococci and a new standard flow cytometric method to measure the opsonophagocytosis activity by rabbit PMNs was shown by this study.
PDF Abstract XML References Citation
How to cite this article
S. D. Siadat, M. Kheirandish, D. Norouzian, Q. Behzadiyannejad, S. Najar Peerayeh, M. Zangeneh and M. Nejati, 2007. A Flow Cytometric Opsonophagocytic Assay for Measurement of Functional Antibodies Elicited after Immunization with Outer Membrane Vesicle of Neisseria meningitidis serogroup B. Pakistan Journal of Biological Sciences, 10: 3578-3584.
DOI: 10.3923/pjbs.2007.3578.3584
URL: https://scialert.net/abstract/?doi=pjbs.2007.3578.3584
DOI: 10.3923/pjbs.2007.3578.3584
URL: https://scialert.net/abstract/?doi=pjbs.2007.3578.3584
INTRODUCTION
Meningococcal disease remains a global public health problem and continues to be a significant cause of morbidity and mortality in the worldwide (Rosenstin et al., 2001). Serogroup B causes the majority (50-80%) of all N. meningitidis cases in Europe and the United States. In order to develop effective vaccines against serogroup B meningococcal disease, effective methods to determine vaccine efficacy need to be established (Plested et al., 2001; Cartwright, 2001). Specific antibodies and a functional complement system are of crucial importance in the host defense against systemic meningococcal infections (Balmer and Borrow, 2004). N. meningitidis is efficiently phagocytosed by PMN cells and antibodies are bactericidal in the presence of peripheral blood polymorphonuclear leukocytes and complement. Consequently, patients with defects in the terminal complement pathway are highly susceptible to meningococcal disease, especially from meningococci of uncommon serogroups (Aase et al., 1998; Martinez et al., 2002). However, the presence of serum opsonins to facilitate phagocytic killing is also likely to be of great importance in the in vivo defense against this organism. The previous studies have shown that serogroup B meningococci are more resistant to bactericidal killing than serogroup A and C meningococci but are highly susceptible to killing by PMN after opsonization (Aase et al., 1995). During complement activation, C3 and C4 split products will be deposited on the bacterial surface and these fragments may be effective opsonins. IgG antibodies bound to bacteria are also excellent opsonins and a synergistic opsonic effect is achieved when the target is covered with both IgG and complement split products. PMN and macrophages constitutively express Fcγ receptors (FcγRs) and complement receptors. An immune response in which the effector function is biased against phagocytosis rather than bacteriolysis may be beneficial to the host, since intracellular destruction will minimize intravascular release of bacterial endotoxin and thus reduce the risk for septic shock (Jack et al., 1994; Aase et al., 1995). Therefore, the in vitro opsonophagocytic activity (OPA) of antibodies to meningococcal components is believed to be a measure of their functional activity in vivo (Väkeväinen et al., 2001). Thus, to determine the serological correlates or surrogates of protection from the samples of ongoing efficacy trials, both quantitative and qualitative characteristics of antibodies have to be measured reliably (Romero-Steiner et al., 1997). This report describes a flow cytometric assay to measure Respiratory Burst (RB) in PMN with serogroup A and B meningococci as the target cells. RB is a terminal effect of phagocytosis thought to reflect microbial killing. Postvaccination sera were collected in period times after immunization with a meningococcal conjugate vaccine.
We use complement-inactivated test sera and add a constant amount of an external complement source to each sample. By this means we may easily quantitate the opsonic activity in postvaccination serum samples. The aim of this study was to establish a modified flow cytometric method to measure the opsonophagocytosis activity and RB by rabbit PMNs and see if this assay might be used as a test to study the immune response after vaccination.
MATERIALS AND METHODS
Preparation of OMVs: OMVs were prepared as described previously (Siadat et al., 2006). In brief, N. meningitidis serogroup B strain (CSBPI, G-245) was grown under controlled-submerge cultural condition in fermentor containing modified Frantz medium at 36°C for 24 h up to early stationary phase. Outer membrane vesicles (OMVs) were extracted in 0.1 M Tris-HCl, pH 8.6, 10 mM EDTA and 0.5% w/v deoxycholate. Purification of OMVs was done by sequential centrifugation at 20,000 g for 30 min and finally followed by ultracentrifugation at 125,000 g for 2 h, the pelleted OMVs were homogenized in phosphate buffered saline(PBS) pH 7.2. Throughout the process thiomersal (100 mg L-1) was added as preservative (Arigita et al., 2004; Claassen et al., 1996; Norheim et al., 2005; Siadat et al., 2006).
Physicochemical analysis of OMVs
Protein determination: Protein content of OMVs was measured according to Peterson (Peterson, 1977).
Electron microscopy: OMV integrity was checked by electron microscopy. OMVs were ultrasonically treated to disperse the vesicles and were attached to Formvar/carbon-coated nickel grids. Grids were washed with a 0.01 M PBS supplement with 0.5% bovine serum albumin (BSA-Sigma) and 0.1% gelatin (PBG-Sigma) and vesicles on the grids were fixed briefly with 1% glutaraldehyde in PBS and negatively stained with potassium phosphotungstate pH 6.0. The grids were examined in a Zeiss CEA902A electron microscope at 80 kV (Siadat et al., 2006).
Biological activity: The biological activity of the endotoxin content of OMVs were estimated by the Limulus Amebocyte Lysate (LAL) gelatin test. The lipid-A part of the endotoxin molecule can activate the gelation of the limulus lysate. The OMVs were incubated in a fivefold dilution with a fixed concentration of limulus lysate at 37°C for 45 min. The endotoxin activity was visualized using Bromthymol Blue. Unknown activity of endotoxin was compared with Escherichia coli standard endotoxin (FDA, Bethesda, USA).
To determine the pyrogenicity of the OMVs a study in rabbits was performed (Pyrogen test, 1995). Samples of the OMVs final lot were injected I.V. in three healthy rabbits (1 mL kg-1), that have not been injected previously, at 1:100, 1:300 and 1:1000 dilution and the rise of body temperature was monitored. Furthermore a placebo which contains only Freund’s complete adjuvant and thiomersal, was injected at 1:100 dilution. For the 1:100 vaccine dilution a repeat of the test was necessary (World health organization).
Immunization procedures: For immunization of rabbit the OMVs were adsorbed to the adjuvant Al(OH)3 with a protein: Al(OH)3 ratio of 1:66(w/v) as previously described and diluted in saline. New Zealand white rabbit weighing 2-2.5 kg (n = 5 rabbit per group) were immunized intramuscularly with 40 μg protein of each of the vaccines on 0, 14, 28 and 42 days and bled on 0, 14, 28, 42 and 56 days. Sera were stored at -20°C.
PMN: Venous blood from healthy rabbit was drawn into heparinized vacuum tubes and the erythrocytes were lysed with a solution containing 8.3 mg mL-1 of NH4Cl, 1 mg mL-1 of NaHCO3 and 0.08 mg mL-1 of EDTA (pH 6.8). The leukocytes were washed twice with Hanks Balanced Salt Solution (HBSS) with 0.2% Bovine Serum Albumin (BSA) and the cell concentration was adjusted to 5x103 mL-1. No additional purification of PMN was required, because further analyses were done by flow cytometry that easily discriminates PMN from other cells within the suspension (Aase et al., 1995).
Complement source for RB assay: Normal rabbit serum with no detectable antibody against N. meningitidis serogroup B CSBPI, G245 was used as the source of complement for opsonization of meningococci. We found no antibodies against meningococcal antigens measured at serum dilutions starting at 1:20 in an ELISA with whole-cell N. meningitidis as the antigen and developed with an alkaline phosphatase-conjugated anti-IgG. The complement source also did not give any measurable RB to the CSBPI, G 243 and CSBPI, G 245 strains. The serum was aliquoted in small volumes, stored at -85°C and thawed immediately before use (Aase et al., 1995; Jodar et al., 2000).
Bacteria to be used in RB assay: N. meningitidis serogroup B CSBPI, G-245 were grown as described previously for Serum Bactericidal Activity (SBA), fixed in 70% ethanol overnight at 20°C and washed in HBSS. The concentration was adjusted to 103 CFU mL-1, aliquoted in 1 mL ampoules and stored at -85°C (Aase et al., 1995).
RB: The RB analysis was performed mainly as described by with some modification (Aase et al., 1995; Chung and Li, 2003; Fleck et al., 2003). All sera to be tested were heated to 56°C for 30 min to inactivate endogenous complement. Fifty microliters of a threefold dilution of sera was mixed with 5 mL of serogroup B meningococci (103 bacteria per mL) in U-bottomed microtiter plates and incubated for 30 min at 37°C with continuous agitation. Then, 5 μL of serum as the complement source was added and the incubation continued for 8 min at 37°C with agitation. Each dilution was tested in duplicate. As an indicator for RB, we used the nonfluorescent probe dihydrorhodamine 123 (DHR) that will be oxidized to fluorescent rhodamine 123 during RB. A stock solution of DHR was prepared by dissolving 10 mg in 1 mL of dimethyl sulfoxide. This solution was aliquoted and stored at -85°C until use. DHR was added to the effector cells to give a final concentration of 10 μg mL-1 just before mixing 50 μL of the effector cells with the opsonized bacteria and the incubation continued for 8 min at 37°C with agitation. Each sample was tested in duplicate. The reactions were stopped by placing the microtiter plates in an ice bath until RB was measured by flow cytometry.
The Phorbol Myristate Acetate (PMA) working solutions were freshly prepared before each assay of RB. The PMA working solution (100 ng mL-1) was prepared by 1:10 dilution of PMA stock (1 mg mL-1) in PBS. The following protocol was used for measuring naturally function of PMA (as positive control): the reaction was performed in 100 μl of whole-blood specimen. First, the DHR solution (50 mM) was added to each tube and incubated for 5 min at 37°C. Then, the PMA solution (100 mg mL-1) was added to each sample and the tube was further incubated for 10 min in the 37°C water bath. Finally, the reaction was stopped by washing the cells once with 4°C PBS (3 mL). The RBCs were lysed using the following ammonium chloride lysing method. Cells incubated with DHR only served as negative controls. All control samples were tested according to the above protocol for the evaluation of RB (Chung and Li, 2003).
Flow cytometry: The samples were run on a flow cytometer (Coulter EPICS-XL-Profile USA) with a 15 mW argon laser. The excitation wavelength was 488 nm and standard Coulter filters were used in all measurements. Neutrophils, monocytes and lymphocytes could be clearly defined and separated on the basis of forward scatter and sideward scatter characteristics. On the scatter histogram, a gate was set on the PMN and RB was measured as percent positive cells within the gate with three-decade logarithmic amplification on the fluorescence detector. By this method PMN could easily be discriminated from monocytes and lymphocytes (and bacteria) without any further purification of the leukocyte fraction. As a negative control, the test serum was omitted and replaced by HBSS containing BSA. This mixture was used to set the correct region on the fluorescent axis. About 10,000 effector cells were counted in each sample and all determinations were performed in duplicate. Phagocytic function and intracellular oxidative burst generation by rabbit PMN, against N. meningitidis serogroup B, were measured with flow cytometer, using dihydrorhodamine-123 as probes, respectively. In these experiments, viable meninigococci, CSBPI G-245 serogroup B, grown to log phase, was used as target cells and DHR-123 primed PMNs from a healthy donor rabbit were used as effector cells. The results are presented by summarizing the percent RB positive PMN at each sample. Activities below 10% at any dilution are defined as zero (Chung and Li, 2003; Lehmann et al., 2000).
RESULTS
OMVs characterization and identity test: Total protein yield of the OMV production was determined after extraction and purification. Total protein yield ranged from 350 to 950 mg in the final product.
Electron microscopy was used order to verify the integrity of the OMV after preparation. OMV size ranged from 70 to 120 nm in this process (Fig. 1). Intactness of the vesicles in this preparation ranged from 50 to 80% of the vesicles.
The biological activity of the endotoxin was determined in the Limulus Amoebocyte Lysate (LAL) assay. OMV final lot samples were tested in a fivefold dilution and contain 677 and 135 EU mL-1, respectively, when compared with E. coli standard endotoxin. The endotoxin activity is within the range of DPT/polio vaccines and is therefore regarded as safe. The endotoxin activity of LPS in vesicles is much lower than that of free purified LPS.
On the basis of the results of the pyrogenicity test, temperature rises of 1:300 and 1:1000 dilutions were comparable with those obtained with the placebo (data not shown). 1:1000 dilution of the vaccine corresponds with the requirements for 23-valent pneumococcal polysaccharide vaccine and mono-, bi-and tetravalent meningococcal polysaccharide vaccines.
Performance of the RB assay: The immune response of strain CSBPI G-245 serogroup B meningococci after vaccination was measured by PMN-mediated RB. We used a Coulter Epics XL-Profile USA with an argon laser operating at 488 nm. Lymphocytes and non-lymphocytes (monocytes and polymorphonuclear leukocytes, i.e., the potentially phagocytosing cells) can be discriminated and quantified by combined measurements of Forward Scatter (FS), which is related to the size of cells and Side Scatter (SS), which is related to granularity of cells. In present experiments, oxidative burst has mostly been assayed by the percentage of PMN with R-123 and by the mean that nonfluorescent DHR-123 is converted during the respiratory burst (major H2O2 release) into a fluorescent compound R-123. Sera taken from fifth different time points (prevaccination, 2 weeks after the first dose, 2 weeks after the second dose, 2 weeks after the third dose and 2 weeks after the fourth dose,) were analyzed. Each serum sample was heated to 56°C for 30 min to inactivate endogenous complement and each of sera was measured for RB activity. RB results were calculated from the area under the histogram bars where each sample is plotted on the abscissa and the percent fluorescence-positive PMN is plotted on the ordinate. The percent positive PMN at each test is summarized to give ΣRB% and have indicated opsonophagocytic responses.
OMVs induced a highly significant increase in opsonophagocytic responses (ΣRB%) against the serogroup B strain 2 weeks after the first dose (p<0.05).
 | |
| Fig. 1: | Electron microscopy of Outer Membrane Vesicle (OMV) stained with 1% potassium phosphotungstate |
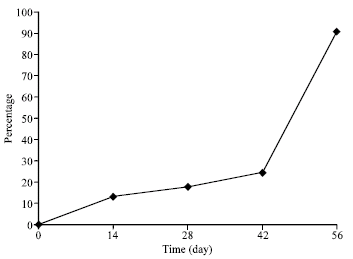 | |
| Fig. 2: | Opsonophagocytosis activity of different titer of rabbit sera against Neisseria meningitidis after immunization with four doses of OMVs |
The clear booster effects were seen after the second dose in 28 days and strongly after the fourth dose in 56 days (Fig. 2) (p<0.05).
A significant shift in the fluorescence histogram from a low fluorescence to a high fluorescence after PMA treatment was seen. This increase in cell fluorescence results from the fact that nonfluorescent DHR-123 is converted during the respiratory burst into a strong green fluorescent compound R-123 (data not shown).
All opsonophagocytic experiments were repeated at least twice. The difference between duplicates was regularly less than 5%.
DISCUSSION
In order to develop effective vaccines against serogroup B meningococci, the relative contributions of antibodies against serogroup B lipopolysaccharide and the various meningococcal outer membrane components in protection against meningococcal disease need to be established (Ruggeberg and Pollard, 2004; Plested et al., 2001; Katial et al., 2002). Among these components, the OMV was chosen instead of OMP, because of the poor immunogenicity of purified OMP, as compared to OMV and may be explained by the absence of lipooligosaccharide (LOS) in these preparations, which has been reported to have immune response stimulating properties. In principal, OMV has potent delivery adjuvant characteristics (Bethell and Pollard, 2002; Jensen et al., 2000; Fukasawa et al., 2004).
The opsonophagocytic activity of antibodies have been documented by others using bacteria that generate high levels of antibodies. Lortan et al. (1993) showed a significant correlation between phagocytosis and the serum levels of anti-pneumococcal polysaccharide antibodies of the IgG, although they did not test each subclass as purified preparations. Bredius et al. (1993) studied opsonophagocytic activity of purified IgG1 and IGg2 antibodies against Staphylococcus aureus and Haemophilus influenzae type b. Similar data were also supplied by Amir et al. (1990).
Flow cytometry has been increasingly used to evaluated the functional capabilities of PMN in host defense. Investigators interested in studying the function of immune system cells have utilized bacteria to elicit cellular responses (Falkow and Validivia, 1998; Lehmann et al., 2000). In such applications, the intracellular oxidation of compounds such as dihydrorhodamine-123 to its fluorescent derivative, rhodamine-123, is indicative of oxygen radical production; Therefore, in flow cytometric analysis, quantitative measurements of bacterial association and oxidative burst can be done on a per cell basis (Lun et al., 2000; Romero-Steiner et al., 1997; Väkeväinen et al., 2001). RB be related effector functions and be induced by antibodies and complement. Complement-mediated opsonization needs activation through C3 to cause deposits of C3b, iC3b and C4b that then ligate to the corresponding receptors on phagocytes. A synergistic opsonic effect is achieved when the phagocytes are triggered through both complement receptors and FcγRs. The potential for an antibody to bind FcR or activate the complement cascade is strongly dependent of the antibody isotype. IgM antibodies are powerful activators of the complement cascade, but they probably cannot induce RB by FcR binding, as there are few FcμRs on neutrophils. Also, the different IgG subclasses reveal individual patterns for these effector functions. Several protocols to measure opsonophagocytic activity against meningococci have been described. Several reports describe chemiluminescence methods to measure the production of reactive oxygen intermediates generated during phagocytosis(Aase et al., 1995, 1998). All these reports use the test serum itself as the complement source, which may be inconvenient for several reasons: all sera must be collected by standardized procedures to preserve complement activity, sera must be tested at high concentrations to avoid diluting the complement components that will consequently make it difficult to differentiate between stronger responses (as some responses may be more or less off scale) and there is high consumption of serum. Also, the complement activity and other serum factors may differ from one rabbit to another and the responses measured may thus not reflect specific antibody responses (Aase et al., 1998). In our RB assay we use an external complement source, ensuring identical complement activity among the different serum samples and the dilution effect is omitted, since a constant amount of complement source is added to all test serum dilutions. We prefer to measure RB rather than phagocytosis. This late event of phagocytosis is probably more related to bacterial killing than mere ingestion and is therefore more relevant to measure in vaccine response studies. Phagocytosis (ingestion) might also be measured in a similar flow cytometric technique, but then the bacteria have to be labelled with a fluorochrome such as FITC. Such surface labelling might modify or disguise relevant antigens and additional techniques must be introduced to discriminate between internalized and adherent bacteria (Lun et al., 2000; Jansen et al., 2001).
In this study we have analyzed the terminal effector functions and opsonophagocytic activity of opsonic antibody that induced against live group B meningococci in rabbit’s whole blood with flow cytometry.
Present results showed that anti-meningococcal opsonic antibodies may all effectively contribute to protection against disease caused by this microorganism. It is not clear which of these effector functions is the most important, but we may anticipate that both are involved: followed by antibody binding the complement cascade is immediately activated and the complement-mediated destruction will begin. Concurrently, PMNs and other phagocytic cells may start engulfing opsonized meningococci (viable, complement-killing and deberis). This may direct much of the microbial destruction to intracellular compartment and perhaps limit some of the damages induced by LPS. Whether one of these effector functions alone will be protective against group B meningococcal disease is highly uncertain.
This study confirms previous results that the OMV vaccine from a serogroup B meningococcal strain is highly immunogenic in rabbit, capable of inducing opsonophagocytic antibodies. It also was to establish a modified standard flow cytometric method to measure the opsonophagocytosis activity by rabbit.
REFERENCES
- Aase, A.,G. Bjune, E.A. Hoiby, E. Rosenqvist, A.K. Pedersen and T.E. Michaelsen, 1995. Comparison among opsonic activity antimeningococcal immunoglobulin response and serum bactericidal activity against meningococci in sera from vaccines after immunization with a serogroup b outr membrane vesicle vaccine. Infect. Immunol., 63: 3531-3536.
PubMedDirect Link - Aase, H�iby A. and E.A.T.E. Michaelsen, 1998. Opsonophagocytic and bactericidal activity mediated by purified igg subclass antibodies after vaccination with the norwegian group b meningococcal vaccine. Scand J. Immunol., 47: 388-396.
Direct Link - Amir, J., M.G. Scott, M.H. Nahm and D.M. Granoff, 1990. Bactricidal and opsonic activity of igg1 and igg2 anticapsular antibodies to haemaphilus influenza type B. J. Infect. Dis., 162: 163-171.
PubMedDirect Link - Arigita, C., W. Jiskoot, J. Westdijk, C. Ingen and W.E. Hennink et al., 2004. Stability of mono-and trivalent meningococcal outer membrane vesicle vaccines. Vaccine, 22: 629-643.
Direct Link - Balmer, P. and R. Borrow, 2004. Serologic correlates of protection for evaluating the response to meningococcal vaccine. Expert. Rev. Vaccines, 3: 77-87.
CrossRef - Bredius, R.G., C.E. Vrise and A. Torelstra, 1993. Phagocytosis of Staphylococcus aureus and Haemophilus influenzae type b opsonized with polyclonal igg1 and igg2 antibodies. J. Immunol., 151: 1463-1472.
Direct Link - Chung, S.C.S. and W. Li, 2003. Flow cytometric evaluation of leukocyte function in rat whole blood. In vitro Cell Dev. Biol. Anim., 39: 413-419.
Direct Link - Claassen, I., J. Meylis, P. Ley, C. Peeters and H. Brons et al., 1996. Production characterization and control of Neisseria meningitidis hexavalent class 1 outer membrane protein containing vesicles vaccine. Vaccine, 14: 1001-1008.
CrossRefDirect Link - Falkow, S. and R.H. Valdivia, 1998. Flow cytometry and bacterial pathogenesis. Curr. Opin. Microbiol., 1: 359-363.
CrossRef - Fleck, R.A., H. Athwal and J.A. Bygraves, 2003. Optimization of NB-4 and HL-60 differentiation for use in opsonophagocytosis assays In vitro. Cell Dev. Biol. Anim., 36: 235-242.
CrossRef - Fukasawa, L.O., W.O. Dias, R.P.F. Schenkman, I. Raw and M. Tanizaki, 2004. Adjuvant can improve protection induced by omv vaccine against neisseria meningitidis serogroup b/c in neonatal mice. Fems Immun. Med. Microbial., 41: 205-210.
Direct Link - Jack, R.M., B.A. Lowenstein and A. Nicholson-Waller, 1994. Regulation of c1q receptor expression on human polymorphonuclear leukocytes. J. Immunol., 153: 262-269.
Direct Link - Jensen, C., B. Kuipers, J. Van Der Bienzen, H. Cock, P. Van Der Ley and J. Tommassen, 2000. Immunogenicity of in vitro folded outer membrane protein PorA of Neisseria meningitidis. Fems Immunol. Med. Microbiol., 27: 227-233.
Direct Link - Jensen, W.T.M., M. Vakevainen-Anttila, H. Kayhty, M. Nahm and N. Bakker et al., 2001. Comparison of a classical phagocytosis assay and a flow cytometry assay for assessment of the phagocytic capacity of sera from adults vaccinated with pneumococcal conjugate vaccine. Clin. Lab. Immun., 8: 245-250.
CrossRef - Katial, R.K., B.L. Brandt, E.E. Moran, S. Marks, V. Agnello and W.D. Zollinger, 2002. Immunogenicity and safety testing of a group B intranasal meningococcal native outer membrane vesicle vaccine. Infect. Immunol., 70: 702-707.
CrossRefDirect Link - Lehmann, A.K., S. Sqrnes and A. Halstensen, 2000. Phagocytosis measurement by flow cytometry. J. Immunol. Meth., 243: 229-242.
CrossRef - Lortan, J.E., A.S. Kaniuk and M.A. Monteil, 1993. Relationship of in vitro phagocytosis of serotype 14 Streptococcus pneumoniae to specific class and IgG subclass antibody levels in healty adults. Clin. Exp. Immunol., 91: 54-57.
Direct Link - Lun, A., M. Schmitt and H. Renz, 2000. Phagocytosis and oxidative burst reference values for flow cytometric assay independent of age. Clin. Chem., 46: 1836-1839.
Direct Link - Martinez, J., T. Pilishvili, S. Barnard, J. Caba, W. Spear, S. Romero-Steiner and G.M. Carlone, 2002. Opsonophagocytosis of fluorescent polystyrene beads coupled to Neisseria meningitidis serogroup A, C, Y, or W135 polysaccharide correlates with serum bactericidial activity. Clin. Diag. Lab. Immunol., 9: 485-488.
CrossRefDirect Link - Norheim, G., A. Aase, D.A. Caugant, E.A. Hoiby and E. Eritzsonn et al., 2005. Development and characterization of outer membrane vesicle vaccines against serogroup A Neisseria meningitids. Vaccine, 23: 3762-3774.
CrossRef - Plested, J., B.L. Ferry, P.A. Coull, K. Makepeace and A.K. Lehmann et al., 2001. Functional opsonic activity of human serum antibodies to inner core Lipopolysaccharide (gale) of serogroup B meningococci measured by flow cytometry. Infect. Immun., 69: 3203-3213.
CrossRef - Peterson, G.L., 1977. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem., 83: 346-356.
CrossRefPubMedDirect Link - Romero-Steiner, S., D. Libutti L.B. Pais, J. Dykes and P. Anderson et al., 1997. Standardization of an opsonophagocytic assay for the measurement of functional antibody activity against streptococcus pneumoniae using different HL-60 cells. Clin. Diag. Lab. Immun., 4: 415-422.
Direct Link - Vakevainen, M., W. Jansen, E. Sadand, I. Jonsdottir and H. Snippe et al., 2001. Are the opsonophagocytic activities of antibodies in infant sera measured by different pneumococcal phagocytosis assay comparable? Clin. Diag. Lab. Immun., 8: 363-369.
CrossRefDirect Link - Cartwright, K., 2001. Meningococcal disease in europe epidemiology mortality and prevention with conjugate vaccines report of a european advisory board meeting vienna. Vaccine, 19: 4347-4356.
CrossRef