
M. Sheibani
Department of Horticulture, College of Agriculture, Ferdowsi University of Mashhad, P.O. Box 9177948978, Mashhad, Iran
A. V. Azghandi
Department of Tissue Culture and Gene Transformation, Agricultural Biotech. Res. InsI. of Iran (ABRIl),
P.O. Box 31535-1897, Karaj, Iran
S. H. Nemati
Department of Horticulture, College of Agriculture, Ferdowsi University of Mashhad, P.O. Box 9177948978, Mashhad, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3564-3570







ABSTRACT
In vitro propagation of saffron either through somatic embryogenesis or cormogenesis is considered to be an efficient alternative method for large-scale propagation of pathogen-free corms. In order to develop an efficient protocol for in vitro propagation of saffron, a factorial experiment was carried out based on completely randomized design to investigate the effects of various concentrations of TDZ (0, 0.1, 0.25 and 0.5 mg L-1) on somatic embryogenesis induction from 5 different types of corm explants (terminal or axillary buds, upper or lower parts of the corm tissue and terminal buds from pre-treated corms at 4°C for 2 weeks). The results revealed that TDZ concentrations affected the induction of somatic embryogenesis significantly while different types of corm explants showed no significant effect on this process. Among TDZ concentrations used, 0.5 mg L-1 was the most effective treatment for embryogenesis induction. Embryogenic calli (globular stage) proliferated well when subcultured into MS medium supplemented with 0.25 mg L-1 TDZ before transferring to hormone-free MS medium containing 6% sucrose for maturation (scutellar or horn-shape stage). Matured embryos were transferred to half strength MS medium without growth regulators for further development, from which microcorms were produced at the basal part after 3 months.
PDF Abstract XML References Citation
How to cite this article
M. Sheibani, A. V. Azghandi and S. H. Nemati, 2007. Induction of Somatic Embryogenesis in Saffron Using Thidiazuron (TDZ). Pakistan Journal of Biological Sciences, 10: 3564-3570.
DOI: 10.3923/pjbs.2007.3564.3570
URL: https://scialert.net/abstract/?doi=pjbs.2007.3564.3570
DOI: 10.3923/pjbs.2007.3564.3570
URL: https://scialert.net/abstract/?doi=pjbs.2007.3564.3570
INTRODUCTION
Saffron (Crocus sativus L.) is one of the most valuable industrial crops in Iran which is particularly important in exportation and income revenue. Iran has the first rank regarding saffron cultivation areas (54700 hectares) and its production (230 tons) in the world (Anonymous, 2004). Saffron is a sterile triploid plant which is naturally propagated vegetatively by daughter corms developing on each mother corm. Each year about 3-4 new corms are produced and therefore the propagation rate of saffron is relatively slow. Traditionally, saffron is cultivated by sowing the rest corms of previous growth season in late summer or early autumn.
Since different pathogens attack the saffron corms, they cause rotting and necrosis and reduce growth and flowering (Plessner et al., 1990; Plessner and Ziv, 1999) they may even stay alive after replanting. So, further expansion of pathogens and more reduction of flowering and saffron yield would be expected by this method each year. The average saffron stigma yield in the last ten years has been 4.4 kg ha-1 which shows a considerable reduction compared to the 2 previous decades (5.3 kg ha-1), while potentially it is about 10 kg ha-1 or even more (Kafi, 2002; Mollafilabi, 2002). One of the important factors that might cause this reduction is using infected corms in the new or re-planted farms. Finding such a method to produce and supply pathogen-free corms rapidly and efficiently can reduce this problem and cause the endurance and stability of production and increase the saffron yield considerably. Mass production of pathogen-free corms would be also needed for modernization of saffron cultivation (Plessner and Ziv, 1999).
Like many other cormous and bulbous plants, meristem culture and regeneration through tissue culture is the most effective method for mass production of healthy and pathogen-free saffron plant materials. In vitro corm production through somatic embryogenesis as an effective method for plant regeneration could be one of the most important aspects of tissue culture application in saffron.
There are several reports on the application of TDZ, as a very effective growth regulator for somatic embryogenesis induction (Zhang et al., 2001; Ipecki and Gozukirmizi, 2003; Panaia et al., 2004; Giridhar et al., 2004; Belokurova et al., 2004) and for induction of organogenesis in different plant species (Caramori et al., 2001; Mithila et al., 2003; Haensch, 2004; Sharma et al., 2005a; Mundhara and Rashid, 2006). In addition to the effects of medium addenda, the tissue or organ type, its physiological status and the genotype of the donor plant appear to be important in the induction of somatic embryogenesis (Finer, 1994). Various plant tissues have been used for regeneration and induction of somatic embryogenesis in saffron including terminal bud (Plessner et al., 1990; Piqueras et al., 1999), axillary bud (Aguero and Tizio, 1994), meristematic zone (Karamian, 2004), leaf segments (Huang, 1987) and ovary (Bhagyalakshmi, 1999).
There are a few investigations published on saffron somatic embryogenesis (Karamian and Ebrahimzadeh, 2003; Karamian, 2004; Blasquez et al., 2004a, b; Alvarez-Orti et al., 2004). Also there are some other researches carried out on in vitro regeneration of saffron through cormogenesis (Homes et al., 1987; Plessner et al., 1990; Aguero and Tizio, 1994; Piqueras et al., 1999; Blasquez et al., 2001) or via organogenesis (Bhagyalakshmi, 1999; Ebrahimzadeh et al., 1996; Ebrahimzadeh and Rajabian, 1998). However, in spite of these studies, so far there is not a defined and routine protocol, available for in vitro propagation of saffron corms. In the present study, effect of various concentrations of TDZ on saffron somatic embryogenesis induction from different types of corm explants was investigated. Further studies were also conducted on maturation of somatic embryos, plantlet regeneration and finally microcorm production.
MATERIALS AND METHODS
Plant materials and surface sterilization of explants: Rest corms were collected from a 4 year-old saffron farm in Torbat e Heydarieh (Khorasan province) in September 2004. After surface sterilization, different parts of the corm tissue (terminal bud, axillary bud, upper or lower parts of the corm tissue and the terminal buds from pretreated corms at 4°C for 2 weeks) were used as explants (Fig. 1). For surface sterilization, the scales were removed and the corms were sterilized using 70% ethanol for 45 sec and then 0.2 % (w/v) HgCl2 solution for 20 min and finally rinsing the corms with sterile distilled water 3 times for 15 min.
Culture media: MS basal medium (Murashige and Skoog, 1962) supplemented with 3% sucrose and 7 g L-1 plant agar was prepared and the pH was adjusted to 5.8 with 1N NaOH or 1 N HCl prior to autoclaving at 121°C for 15 min. TDZ was added after filter sterilization using 0.2 μm pore-size disposable filters. When the temperature of the media was about 50°C, sterile TDZ was added to the media in different concentrations (0, 0.1, 0.25, 0.5 mg L-1) and the media were distributed in Petri dishes and gelled.
Somatic embryogenesis induction: After preparing the media, different types of explant from the corm tissue (Fig. 1) were cultured in Petri dishes (90x15 mm). Cultures were maintained at 25±1°C under 16/8 h (light/darkness) photoperiod with a light intensity of approximately 4000 lux. These cultures were subcultured every 4 weeks. After 10 weeks, numbers of globular embryos induced on each explant were counted.
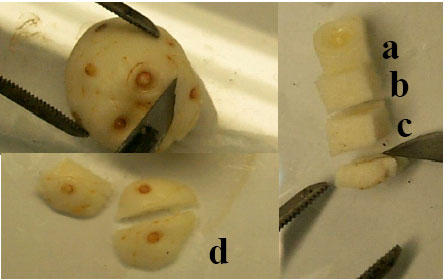 | |
| Fig. 1: | Different parts of corm tissue used as explant (a. terminal bud, b and c. upper and lower parts of the corm tissue, d. axillary buds) |
The experiment was carried out as a factorial based on completely randomized design with 20 treatments in 3 replications (Petri dishes) and 5 explants in each replication.
Maturation of embryos: After further proliferation on MS medium supplemented with 0.25 mg L-1 TDZ, globular somatic embryos were transferred to MS medium without TDZ but containing 2 sucrose concentration levels of 3 and 6% for maturation. These two media were combined with 3 further maturation treatments including incubation at 25±1°C in the light, keeping cultures in the cold room (at 4°C) in the dark and addition of 1 mg L-1 ABA into the medium plus incubation at 25±1°C in the light) for 2 months after then the number of matured embryos in each culture dish (containing 5 embryogenic masses) were counted. This study was also carried out as a factorial experiment based on completely randomized design with 6 treatments, in 5 replications and 5 explants in each replication. Other conditions were similar to the previous experiment.
Data collection and statistical analysis: Data were collected by counting the number of explants with embryogenic response, number of globular embryos induced on each explant, number of horn-shape (matured) embryos and finally the number of plantlets regenerated from each culture dish. These data were analyzed using the General Linear Models (GLM) procedure of SAS (Ver. 6.12) system and the means were compared using LSD test. Data were subjected to transformation and a radical conversion [(X+0.05)**0.5] was used. Non-transformed means have been brought in Tables and Figures.
RESULTS AND DISCUSSION
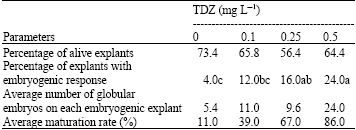
Somatic embryogenesis induction: Embryogenic callus was formed within 2 months, first on the tissues cultured on medium supplemented with 0.5 mg L-1 TDZ and later appeared in two other TDZ concentrations (0.1 and 0.25 mg L-1). The embryogenic calli were often nodular, compact and yellowish (Fig. 2). The induction of somatic embryogenesis was significantly affected by different concentrations of TDZ, while unexpectedly different types of explant did not show any significant effect on this process. Also the interactive effects between TDZ concentrations and explant types were not significant (Table 1). Maximum embryogenic callus was obtained on 0.5 mg L-1 TDZ and the highest number of globular embryos (12.04 embryos per each embryogenic explant) and maximum conversion of globular to horn-shape embryos (86%) were also obtained in this concentration (Table 2).
| Table 1: | Analysis of variances of the effect of various concentrations of TDZ and explant type on embryogenesis induction in saffron |
 | |
| **; Very significant (p<0.01), ns; not significant | |
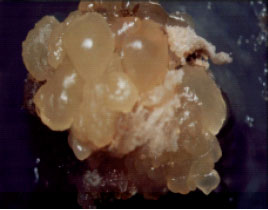 | |
| Fig. 2: | Embryogenesis induction on the lower part of corm tissue cultured on MS medium supplemented with 0.5 mg L-1 TDZ, 2 months after culture |
Since the percentage of explants showing embryogenic response increased with increasing TDZ concentrations, it might be possible to get more responses using higher concentrations of TDZ. Although there are some reports showing that high concentrations of TDZ promote shoot organogenesis and/or multiple shoot formation rather than somatic embryogenesis (Sharma et al., 2005b). Cheong and Pooler (2004) reported that somatic embryogenesis was inhibited by addition of higher concentrations of TDZ to the culture media.
All types of explants produced somatic embryogenesis without an obvious intermediate callus phase and the induction rates were not significantly different among different types of explant used in this experiment. The possible explanation is that all parts of the corm tissue used regardless of their types had embryogenic potential. Another possibility is that the explants, terminal and axillary buds and the upper or lower parts of the corm tissue which had nodular points contained meristemic cells and the existence of these meristemic cells might have been essential for somatic embryogenesis induction.
Thidiazuron has been used for induction of somatic embryogenesis in many plant species like soybean, tobacco, legumes, pelargonium and orchids (Hutchinson et al., 1996; Chen and Chang, 2003; Khan et al., 2005; Kiran et al., 2005).
| Table 2: | Influence of TDZ concentration on embryogenesis parameters |
 | |
 | |
| Fig. 3: | Effect of different maintaining treatments on maturation of somatic embryos |
In many cases, TDZ induced embryogenesis with a higher efficiency and frequency compared to other cytokinins or the combination treatments of auxins and cytokinins (Visser et al., 1992; Zhang et al., 2001; Belokurova et al., 2004; Panaia, 2004). TDZ is thought to affect the endogenous ratio of auxins to cytokinins which in turn influences the induction of somatic embryogenesis (Visser et al., 1992). In the present study, effective embryogenesis induction was observed using TDZ and higher concentrations were more effective. Blasquez et al. (2001) have previously applied TDZ for proliferation of saffron cormogenic calli, but to the best of our knowledge this is the first report on induction of somatic embryogenesis in saffron using TDZ.
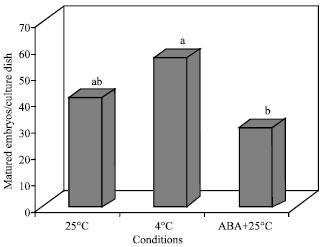
Maturation of embryos: Maturation of somatic embryos was affected by sucrose concentration in the culture medium significantly. The average number of matured embryos in each culture dish was 50 embryos for 6% sucrose compared with 35 embryos in 3% treatment. On the other hand, cultures on MS basal medium incubated in the darkness at 4°C had maximum maturation rate while ones cultured on ABA-containing medium and incubated at 25±1°C in a 16/8 h photoperiod had the minimum maturation rate (Fig. 3 and 4). Using a concentration of 6% sucrose in the culture medium accompanied with keeping the cultures at 4°C in the darkness for 2 months was the most effective treatment for maturation of globular embryos (Table 3 and 4).
| Table 3: | Analysis of variances of maturation treatments, A: sucrose concentration, B: different conditions of maintaining globular embryos (25°C, 4°C and 1 mg L-1 ABA+25°C) |
 | |
| Table 4: | Interactive effects of sucrose concentration and 3 different maintaining conditions on the number of matured embryos of saffron |
 | |
| Means with similar letter(s) are not significantly different (LSD 0.05 = 20.466) | |
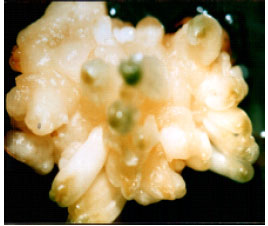 | |
| Fig. 4: | Conversion of globular embryos to horn-shape (matured) embryos on MS medium without growth regulator supplemented with 6% sucrose and conditions at 4°C for 2 months |
Using higher levels of sucrose in the culture medium promote the maturation of somatic embryos (Komatsuda and Ohyana, 1988). However, according to Pence (1992) somatic embryos of alfalfa matured on the medium with high sucrose levels (9%) lost chlorophyll and turned yellow. It was concluded that the synthesis of chlorophyll and lamellar development could be inhibited by high sucrose concentrations. In the present study only two levels of sucrose concentration (3 and 6%) were studied and a beneficial effect was observed when 6% sucrose was used in the culture medium.
Cold treatment (4°C) has also been used for maturation of somatic embryos (Deng and Cornu, 1992; Corredoria et al., 2003). According to Deng and Cornu (1992) pre-treatment of globular embryos in darkness at 4°C for 2 months increased the maturation percentage of walnut somatic embryos. The results obtained in this work are in consistence with these researchers.
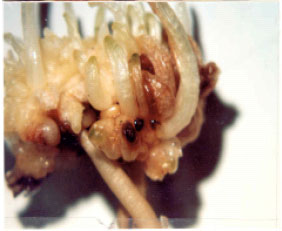 | |
| Fig. 5: | Contractile root formation at the base of an embryogenic mass cultured on ½ MS medium without growth regulator |
 | |
| Fig. 6: | Microcorm production on the embryogenic masses cultured on ½ MS medium without growth regulator after 3 months |
 | |
| Fig. 7: | Returning to globular stage when cultured on ½ MS medium supplemented with 1 mg L-1 NAA + 1 mg L-1 BA |
 | |
| Fig. 8: | Comparison of natural corms (a) with microcorms (b) |
Although ABA has been previously used for embryo maturation of saffron species (Karamian, 2000), in the present experiment no satisfactory results were obtained from ABA-containing culture medium and most of the globular embryos cultured in this medium got brown and died later.
Plantlet regeneration: Within 3 months and afterwards, most of the matured embryos cultured on ½ MS medium without growth regulators produced plantlets with microcorm and occasionally roots (Fig. 5 and 6). Whereas the ones cultured on medium containing 1.0 mg L-1 NAA+1.0 mg L-1 BA returned to globular stage and produced more globular embryos and callus (Fig. 7). Finally, Fig. 8 illustrates a comparison between field-grown natural corms (a) with the in vitro produced microcorms (b).
Although like Chen and Chang (2003) we obtained better results using basal medium devoid of growth regulators for plantlet regeneration from somatic embryos induced and proliferated by TDZ, but this seems to be different from the results reported by Karamian (2000) recommending supplementation of medium with 1 mg L-1 of both NAA and BA for conversion of saffron somatic embryos to plantlets.
CONCLUSIONS
An efficient procedure was developed for induction of somatic embryogenesis from different types of corm tissues of saffron (Crocus sativus L.) using TDZ and further development of somatic embryos, maturation, plantlet regeneration and finally microcorm production were successfully achieved. This procedure can be used both in saffron breeding and improvement projects as well as propagation of pathogen-free microcorms for saffron farmers.
ACKNOWLEDGMENTS
This study was a part of M.Sc. Thesis on Horticulture, Department of Horticultural Science, Faculty of Agriculture, Ferdowsi University of Mashhad, funded by Agricultural Biotechnology Research Institute of Iran (project number 2-013-140000-03-8303-83007). The authors wish to thank and appreciate all of the collaborations.
REFERENCES
- Belokurova, V.B., L.S. Glovach, N.L. Shcherbak and N.V. Kuchuk, 2004. In vitro regeneration of Nicotiana africana plants from explants of different type and mesophyll protoplasts. Tsitol Genet., 38: 9-15.
Direct Link - Bhagyalakshmi, N., 1999. Factors influencing direct shoot regeneration from ovary explants of saffron. Plant Cell Tissue Organ Cult., 58: 205-211.
CrossRefDirect Link - Chen, J.T. and W.C. Chang, 2004. Induction of repetitive embryogenesis from seed-derived protocorms of Phalaneopsis amabilis Var. Formosa shimadzu. In vitro Cell. Dev. Biol. Plant, 24: 290-293.
Direct Link - Cheong, E. and M.R. Pooler, 2004. Factors affecting somatic embryogenesis in Prunus incise cv. February pink. Plant Cell Rep., 22: 810-815.
Direct Link - Corredoria, E., A. Ballester and A.M. Vieitez, 2003. Proliferation, maturation and germination of Castanea sativa Mill. somatic embryos originated from leaf explants. Ann. Bot., 92: 129-136.
Direct Link - Deng, M.D. and D. Cornu, 1992. Maturation and germination of walnut somatic embryos. Plant Cell Tissue Org. Cult., 28: 195-202.
Direct Link - Haensch, K.T., 2004. Thidiazuron-induced morphogenetic response in petiole cultivars of Pelargonium H domesticum and its histological analysis. Plant Cell Rep., 23: 211-217.
Direct Link - Homes, J., M. Legros and M. Jaziri, 1987. In vitro multiplication of Crocus sativus L. Acta Hortic., 11: 675-676.
Direct Link - Hutchinson, M.J., S. Krishnaraj and P.K. Saxena, 1996. Morphological embryogenesis in Geranium (Plargonium H Hortorum Bailey) hypocotyl cultures. J. Plant Sci., 154: 440-446.
Direct Link - Karamian, R., 2004. Plantlet regeneration via somatic embryogenesis in four Crocus specie. Acta Hortic., 650: 253-259.
Direct Link - Khan, H., I. Siddique and M. Anis, 2006. Thidiazuron induced somatic embryogenesis and plant regeneration in Capsicum annum. Biol. Plant, 50: 789-792.
Direct Link - Kiran, G., C.D. Kaviraj, G. Jogeswar, P.B. Kavi Kishor and S. Rao, 2005. Direct and high frequency somatic embryogenesis and plant regeneration from hypocotyls of chickpea (Cicer arietinum L.), a grain legume. Curr. Sci., 89: 1012-1018.
Direct Link - Mithila, J., J.C. Hall, J.M. Victor and P.K. Saxena, 2003. Thidiazuron induces shoot organogenesis at low concentrations and somatic embryogenesis at high concentrations on leaf and petiole explants of African violet (Saintpaulia ionantha Wendl.). Plant Cell Rep., 21: 408-414.
CrossRefDirect Link - Mundhara, R. and A. Rashid, 2006. Recalcitrant grain legume Vigna radiata, mung bean, made to regenerate on change of hormonal and cultural conditions. Plant Cell Tissue Org. Cult., 85: 265-270.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Panaia, M., T. Senaratna, K.W. Dixon and Sivasithamparam, 2004. The role of cytokinins and thidiazuron in the stimulation of somatic embryogenesis in key members of the Restionaceae. Aust. J. Bot., 52: 257-265.
Direct Link - Pence, V.C., 1992. Abscisic acid and the maturation of cacao embryos in vitro. Plant Physiol., 98: 1391-1395.
Direct Link - Piqueras, A., B.H. Han, J. Escribano, C. Rubio, E. Hellin and J.A. Fernandez, 1999. Development of cormogenic nodules and microcorms by tissue culture, a new tool for the multiplication and genetic improvement of saffron. Agronomic, 19: 603-610.
CrossRefDirect Link - Plessner, O., M. Ziv and M. Negbi, 1990. In vitro corm production in the saffron Crocus (Crocus sativus L.). Plant Cell Tissue Org. Cult., 20: 89-94.
Direct Link - Sharma, V.K., R. Hansch, R. Mendel and J. Schulze, 2005. Mature embryo axis-based high frequency somatic embryogenesis and plant regeneration from multiple cultivars of barley (Hordeum vulgare L.). J. Exp. Bot., 56: 1913-1922.
Direct Link - Sharma, V.K., R. Hansch, R.R. Mendel and J. Schulze, 2005. Influence of Picloram and Thidiazuron on high frequency plant regeneration in elite cultivars of wheat with long-term retention of morphogenecity using meristematic shoot segments. Plant Breed., 124: 242-246.
CrossRefDirect Link - Visser, C., J.A. Qureshi, G. Ravinder and P.K. Saxena, 1992. Morphology role of Thidiazuron: Substitution of auxin and cytokinin requirements for the induction of somatic embryogenesis in geranium hypocotyl cultures. Plant Physiol., 99: 1704-1707.
Direct Link - Zhang, C.L., D.F. Chen, M.C. Elliott and A. Slater, 2001. Thidiazuron-induced organogenesis and somatic embryogenesis in sugar beet (Beta vulgaris L.). In vitro Cell. Dev. Biol.-Plant, 37: 305-310.
CrossRefDirect Link