
M. A. Al-Fredan
College of Science, King Faisal University, Al-Hassa, Saudi Arabia
A. A. Fathi
College of Science, King Faisal University, Al-Hassa, Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3210-3214







ABSTRACT
Five soil samples were taken from different cultivated sites from the North Villages of Al-Hasa. Three algal groups were recorded throughout this study, Bacillariophyta, Cyanophyta and Chlorophyta. Fifty-two genera of algae were identified in all the soils sampled during this investigation. Out of these, 11 genera belonged to the Chlorophyta; 16 genera belong to Bacillariophyta and 25 genera to Cyanophyta. Penniate Bacillariophyta (Nitzschia, Gyrosigma), oscillatorid Cyanophyta (Osillatoria, Phormidium, Nostoc and Anabaena) and coccoid Chlorophyta (Ankistrodesmus falacatus, Chlorella ellipsoidea, Chlorella vulgaris, Gleocystis bcillus, Scenedesmus acuminatus, Scenedesmus arvensis and Scenedesmus bijuga) were the predominant in all investigated sites.
PDF Abstract XML References Citation
How to cite this article
M. A. Al-Fredan and A. A. Fathi, 2007. Preliminary Survey of Edaphic algae in Al-Hasa Region, Saudi Arabia. Pakistan Journal of Biological Sciences, 10: 3210-3214.
DOI: 10.3923/pjbs.2007.3210.3214
URL: https://scialert.net/abstract/?doi=pjbs.2007.3210.3214
DOI: 10.3923/pjbs.2007.3210.3214
URL: https://scialert.net/abstract/?doi=pjbs.2007.3210.3214
INTRODUCTION
Algae are generally found on the surface of moist soils, where there is sufficient light for their photosynthetic reactions. The major types present are green algae (Chlorophyceae) and the diatoms (Bacillariophyceae). Their total numbers vary from several hundreds to several thousands. In some situations their number is quite large and brings about beneficial changes, especially desert soils. Soil habitats are the most important non-aqueous ecosystems for algae (Zenova et al., 1995; El-Sheekh et al., 1998; Fathi and Zaki, 2003; Santina, 2006). Algae represent the first community to colonise bare soil, including volcanic deposits and unfarmed soils from various origins, thereby enabling the subsequent establishment of higher plant communities (Starks et al., 1981).
The nature of algal flora in different localities is the result of a complex influence of the local type of vegetation, soil properties and climatic conditions, but it often also depends on the input of airborne algal Diasporas (Lukešova´, 1993; El-Sheekh et al., 1998; Cavacini, 2001; Fathi and Zaki, 2003; Santina, 2006). The interaction of soil biota, e.g. micro-organisms, roots, animals and plants is affected, sometimes significantly, by agricultural practices and human activities. Soil labour, pesticide residues, chemical fertilisers and agronomic practices affect plants, animal life and soil community structure (Paoletti et al., 1988).
The role of algae as pioneer organisms is particularly important in bare soils liable to erosion. Soil algae, especially cyanobacteria, are known to aggregate soil particles by producing extracellular polysaccharides (Lynch and Bragg, 1985) and forming water-stable aggregates that reduce the impact of wind erosion Johansen, 1993). Green and blue-green populations in upper topsoil are large and diverse and they can perform valuable services for the soil ecosystems (Starks et al., 1981) and for agriculture too (Ruble and Davis, 1988).
Al-Hassa is one of the largest oases of the world and located in the southern part of the eastern region of Saudi Arabia. The region of Al-Hassa, with its center at the town of Al-Hofuf, is about 70 km towards the interior of the country from the Gulf coast near Al-Uqayr at elevations of 130 to 160 m above sea level. No efforts have so far been made to study the soil algae in eastern region of Saudi Arabia. This research is a contribution to the knowledge of soil algae from the North Villages of Al-Hasa. The present study may represent the first attempt to survey the algal populations inhibiting cultivated soils in Al-Hasa.
MATERIALS AND METHODS
The studied area and climate: Al-Hasa lies in the south of the Kingdom's Eastern region and is bounded by the Al-Dahna and the Al-Daman deserts. It is situated between 25° 05' and 25° 40' northern latitude and 49° 55' eastern longitude. The Al-Hasa oasis is the largest oasis in the Kingdom of Saudi Arabia and the municipality of Al-Hasa constitutes the largest administrative area in the Kingdom. Al-Hasa has a dry, tropical climate, with a five month summer and a relatively cold winter. It enjoys the benefit of copious reserves of underground water which has allowed the area to develop its agricultural potential. Nevertheless, Al-Hasa has to deal with tons of sand which the wind carries and deposits over the land. To counter this problem, the Kingdom has planted large barriers of trees to prevent the wind-borne sand from damaging inhabited and agricultural areas.
Soil sampling: Five samples were taken from different cultivated sites from the North Villages of Al-Hasa. The samples were collected only once during April, 2006. Selection of these samples was based on variabilities in the cultivated plants and habitat features (Site I was collected by Hibiscus esculentus L., site II was collected by Solanum lycopersicum, site III was collected by Cucurbita pepo, site IV was collected by Spinacia oleracea L. and site V was uncultivated). Soil samples were taken by cores (5 cm diameter) from representative habitats at a depth of zero to two cm. Each soil samples was composed of four random samples from each site.
Laboratory analysis: Soil texture analysis was carried out using the sieving method by where the percentage of gravel, coarse and fine sand, silt and clay were estimated. The hygroscopic moisture content of the soil was estimated by oven drying air-dry samples. Digital pH- meter (Lutron, pH 204) measured the pH value. Conductivity was measured by a direct indicating conductivity bridge. organic matter was determined according to Jackson (1960).
Soil algae: Moist plate method recommended by Jurgensen and Davey (1968) was applied. One gram of each soil sample was placed in 99 mL of sterile water and then placed in a shaker for 15 min. Five replicate petri-dishes were inoculated each with one mL of the appropriate dilution and 25 mL of nutrient agar medium (45°C) were added. Allen and Arnon's (1955) medium incubated at 35°C was used for preferential isolation of blue-green algae. The eukaryotic algae were isolated in Chu’10 (Nichols, 1973) at 25°C. Both were incubated on 16/8 h. Light-dark cycle, with a light intensity (80 μmol m-2 sec-1). This method was recommended by Bold (1970). The numbers of colonies were proportional to dry weight soil.
Diatom frustules were cleaned by soaking in hydrogen peroxide (H2O2) overnight. They were then rinsed and centrifuged repeatedly. All non diatom algae were identified by direct microscopic examination of wet mounts. Algae were identified according to Fott (1972), Bourelly (1981) and Prescott (1987). The appropriate statistic in Brillouin’s index (Pilou, 1966) was used for quantitative analysis of species diversity of the phytoplankton, using the following equation:-
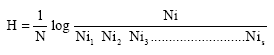 |
Where, H is the diversity index and N is the total number of individuals of Sth species.
RESULTS AND DISCUSSION
It is well known that changes in physico-chemical characteristics and edaphic factors of any soil, with some specific features within the algal organisms themselves constitute the main factors responsible for the existence of soil algae (Ahmed, 1994; El-Attar, 1999; Fathi and Zaki, 2003; Santina, 2006).
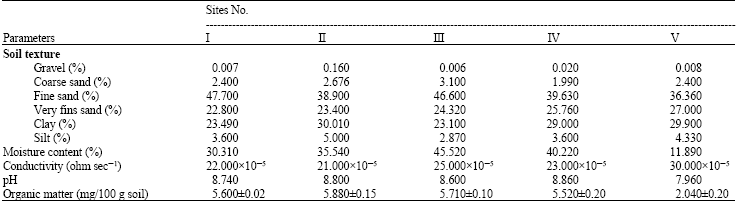
The granulometric analysis of soil showed that the percentage of gravel, coarse sand, fine sand, clay and slit fractions varied remarkably from one site to another (Table 1). Higher percentage of coarse and fine sand was confined to sample (III), while the maximum percentage clay and silt were represented by the soil of site II. On the other hand, higher very fine sand percentage was a distinct character of the non-cultivated site (V). In the present study, the response of algal biomass at the different investigated sites to soil texture was not reflected in retarded or activated algal growth. Similarly the results obtained by Hifney (1998) and Fathi and Zaki, (2003).
The results in Table 1 show that the maximum moisture content of soil samples (45.52%) was recorded at site III, which was cultivated with Cucurbita pepo, whereas the lowest moisture content was recorded at site V (non-cultivated). In general, the moisture content of the study sites ranged from 11.98-45%. The higher values of moisture content performed an elevation in algal biomass. Soil algae grow better in partially dry soil (40-60%) of water holding capacity (Round, 1981 and Darley, 1982). Further, the data of Table 1 revealed that pH values of all sampled soils were always on the alkaline side (7.96-8.86) except site (IV). Shields and Durrell (1964) accepted that neutral and alkaline soils are more favorable to development of blue-green algae. Starks et al. (1981) found that members of Cyanophyta dominated in alkaline soils (pH 8.5-9.5), while the Chlorophyta were better represented in acidic soils (pH 6.0-4.6).
Electric conductivity of sample (V) represented the highest value (30x105 ohm sec-1) followed by site (III), while the lowest value was recorded at site (II) being cultivated with Solanum lycopersicum. The data also show that the collected soil samples were characterized by higher contents of organic matter, which ranged between (2.04-5.88 mg/100 g soils). The maximum value was recorded in site (II), which cultivated by Solanum lycopersicum, whereas the lower value was recorded for site (V) non-cultivated soil (Table 1).
| Table 1: | Average values of some physico-chemical characters of soils collected from different sites of El-Hasa North Villages in April 2006. Mean of three replicates±SE |
 | |
The data of this investigation and those obtained by other investigators (Hifney, 1998), clearly indicated that the inhibited and stimulated growth of certain algal groups or species may be one aspect of the role played by organic matter of soils in modifying algal population and activities in various soil. On the other hand Ketchum (1951) speculated that the organic nutrients might have a great effect on the algae in terms of fertilization than inorganic nutrients. Eutrophication may delimit the diversity of species (Sheath and Munawar, 1974).
Soil algae are present in all kinds of soils, both arable and virgin, in large amounts and in great diversity. These photosynthetic microorganisms, which are concentrated in the top few centimeters of the soil profile, are organized in a community structure that varies depending on soil type, farming method and pesticide application (Berard et al., 2004). The data of this study shows that there are marked differences in the quantitative and qualitative composition of the algal flora in all the soil sampled. (Table 2-4). Cells counts were calculated on the basis of 1.0 g dry soil. Filamentous and colonial forms were counted as one cell. In terms of total cell number for each species in all the algal groups, the total algal counts ranged from a minimum of 15.00x106 cell L-1 at the non-cultivated site (V) to a maximum of 141.5x106 cell L-1 at site II cultivated with Solanum lycopersicu (Fig. 1).
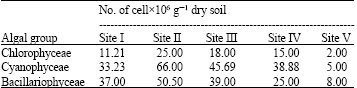
Three algal groups namely, Bacillariophyta, Chlorophyta and Cyanophyta were recorded throughout this study (Table 2). Representative taxa from all these algal groups were recorded in all soil sampled except fewer groups were represented in the uncultivated site (V). The data in Table 3 shows that except the non-cultivated site Cyanophyta (40-49%) dominated the algal taxa in other four soil sampled. Bacillariophyta ranked second (31-45%), Chlorophyta ranked the third (13-19%) in order of dominance. As regards to the total cell number of each algal groups (Table 2); Chlorophyta, the highest cell number was recorded in site II (25.0x106 cell L-1), whereas site V harbored the lowest cell count (2.0x106 cell L-1); Bacillariopohyta counts ranged from 8.0x106 cell L-1 at the non-cultivated site to 50.50x106 cell L-1 at site II cultivated with Barley; The highest cell yield of Cyanophyta was also contained in the soil samples of site II (66.0x106 cell L-1).
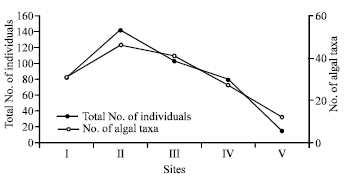 | |
| Fig. 1: | Species richness (total number of algal taxa encounted) and algal abundance (cell number x 106/g dry soil) of soil algae at Al-Hasa, (April, 2006) |
| Table 2: | Cell numbers average of the three main algal groups recorded at Al-Hasa, Saudi Arabia (April, 2006) |
 | |
| Table 3: | The percentage composition of the main algal groups recorded at Al-Hasa, Saudi Arabia (April, 2006) |
 | |
Fifty-two genera of algae were identified in all the soils sampled during this investigation. Out of these, 11 genera belonged to the Chlorophyta; 16 genera belong to Bacillariophyta and 25 genera to Cyanophyta. The maximum abundance of algal taxa (46 species) was observed in the site II cultivated with Solanum lycopersicum, whereas the minimum abundance was observed in the non-cultivated area (12 species).
| Table 4: | Relative abundance of soil algal species of soil samples collected from, different lands at Al-Hasa, Saudi Arabia |
 | |
| + = Recorded | |
As shown in Table 4 the ranking of algal taxa abundance by habitats were as follows: site II > site III > site I > site IV and site V > site IV. The Blue-green algae frequently encountered from 33-50% of the total algal taxa of all soils sampled and there were predominantly the filamentous, Oscillatorid forms, Anabaena, Arthrospira, Calothrix, Oscillatoria, Phormidium, Lyngbya, Schizothrix, Nostoc species. On the other hand green algae were represented mostly by unicellular and colonial/or coccoid forms. The predominant species of Chlorophyta were, Chlorella vulgaris, Gleocystis bcillus, Scenedesmus acuminatus, Scenedesmus arvensis and Scenedesmus bijuga. The dominant taxa of diatoms were, Gyrosigma, Navicula and Nitzschia Ranking the habitats by diversity index (H), there were: site II (2.60) > site III (2.00) > site I (1.88) > site VI (1.56) > site V (0.25).
The presence of numerous Cyanophyta species as compared with other types of soil algae is a matter of tolerance and adaptability (Ahmed, 1994; Hinfey, 1989; El-Attar, 1999). The pH value at all sites under investigation was more than 7 and this might partially explain the wide distribution of Cyanobactertial species, being represented at most sites compared with eukaryotic algal assemblage. However, another additive explanations for the widespread of Cyanophyta members in the different studied sites, may be due to presence of lower amount of carbonate and higher amount of calcium (Gorham et al., 1974) or due to the properties of prokaryotic cells (Fay and Fog, 1962; Trainor, 1978). These results are in accordance with the data of some previous studies at Egypt (Ahmed, 1994; Hifney, 1998; El-Sheekh et al., 1998; El-Attar, 1999; Fathi and Zaki, 2003).
Regarding the common species, penniate Bacillariophyta (Nitzschia, Gyrosigma) and oscillatorid Cyanophyta (Osillatoria, Phormidium, Nostoc and Anabaena) represented the highest abundance of all diatoms recovered in all sites (Table 4). As regards to the Chlorophyta the coccoid forms (Ankistrodesmus falacatus, Chlorella ellipsoidea, Chlorella vulgaris, Gleocystis bcillus, Scenedesmus acuminatus, Scenedesmus arvensis and Scenedesmus bijuga) were the predominant in all investigated sites. Also, Chlorophyta have predilection for organic matter and monovalent cations. These results are in accordance with that of Ahmed (1994), Hinfney, (1998), El-Sheekh et al. (1998) and El-Attar (1999).
CONCLUSIONS
This study may represent the first attempt to survey the algal populations inhibiting cultivated soils in Al-Hasa. In general, the data herein obtained; firstly the appearance of algal species from various soils seemed to be governed by the physico-chemical characters of the soil. Secondly, oscillatorid Cyanophyta, coccoid Chlorophyta and penniate Bacillariophyta represented the predominant forms in almost cultivated soils.
ACKNOWLEDGMENT
We thank the Deanship of Scientific Research, King Faisal University for financial support (Grant No. 7113).
REFERENCES
- Ahmed, A.Z., 1994. Preliminary survey of soil algal flora in Upper Egypt. Egypt. J. Bot., 34: 17-36.
Direct Link - Allen, M.B. and D.I. Arnon, 1955. Studies on nitrogen-fixing blue-green algae. I. Growth and nitrogen fixation by Anabaena cylindrica Lemm. Plant Physiol., 30: 366-372.
Direct Link - Berard, A., F. Rimet, Y. Capowiez and C. Leboulanger, 2004. Procedures for determining the pesticide sensitivity of indigenous soil algae: A possible bioindicator of soil contamination?. Arch. Environ. Contam. Toxicol., 46: 24-31.
PubMedDirect Link - Cavacini, P., 2001. Soil algae from northern Victoria Land (Antarctica). Polar Biosci., 14: 45-60.
Direct Link - Johansen, J.R., 1993. Cryptogamic crusts of semiarid and arid lands of North America. J. Phycol., 29: 140-147.
CrossRefDirect Link - Jurgensen, M.F. and C.B. Davey, 1968. Nitrogen fixating blue-green algae in acid forest and nursery soils. Can. J. Microb., 14: 1179-1179.
PubMed - Pielou, E.C., 1966. The measurement of diversity in different types of biological collections. J. Theor. Biol., 13: 131-144.
CrossRefDirect Link - Zancan, S., R. Trevisan and M.G. Paoletti, 2006. Soil algae composition under different agro-ecosystem in North-Eastern Italy. Agric. Ecosyst. Environ., 112: 1-12.
CrossRef - Sheath, R. and M. Munawar, 1974. Phytoplankton composition of a small subarctic lake in the Northwest Territories, Canada. Phycologia, 13: 149-161.
CrossRefDirect Link - Starks, T.L., L.E. Shubert and F.R. Trainor, 1981. Ecology of soil algae: A review. Phycologia, 20: 65-80.
CrossRefDirect Link - Zenova, G.M., E.A. Shtina, S.N. Dedysh, O.B. Glagoleva, A.A. Likhacheva and T.A. Gracheva, 1995. Ecological relations of algae in biocenoses. Mikrobiologiya, 64: 121-133.
Direct Link