
Rubina Arshad
Nuclear Institute for Agriculture and Biology (NIAB), P.O. Box 128, Jhang Road, Faisalabad, Pakistan
Shafqat Farooq
Nuclear Institute for Agriculture and Biology (NIAB), P.O. Box 128, Jhang Road, Faisalabad, Pakistan
Syed Shahid Ali
Department of Zoology, University of the Punjab, Quaid-e-Azam Campus, Canal Bank Road, Lahore, Pakistan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3190-3194







ABSTRACT
The aim of present study was to optimize conditions for conversion of penicillin G into 6-APA using intact crude cells of locally collected PGA producing bacterial strains as biocatalyst. Corn steep liquor medium supplemented with phenylacetic acid was used for PGA production. For enzymatic conversion of penicillin G into 6-APA by PGA impregnated bacterial cells, a maximum reaction time of 4 h was found adequate. The procedure for extraction and crystallization of 6-APA from the enzyme reaction mixture was standardized. Isolation process was carried out under controlled pH conditions and 6-APA crystals were recovered from the reaction mixture via filtration, concentration and drying. The maximum PGA activity was observed in Escherichia coli strain BDCS-N-FMu12 (6.4 mg 6-APA h-1 mg-1 wet cells) whereas Bacillus megaterium (ATCC 14945 used as check) exhibited only 2.4 mg 6-APA h-1 mg-1 wet cells. The overall yield of 6-APA crystals obtained after enzymatic conversion of penicillin G ranged between 37-55 and 47-68% in foreign and local strains, respectively. BDCS-N-FMu12 was identified as the best PGA producer with 68% 6-APA conversion whereas ATCC 14945 showed the lowest conversion (37%). The recovery of 6-APA (68%) obtained by using crude intact cells as cheap biocatalyst appeared promising. The process of enzyme fermentation and 6-APA crystallization optimized during this study seems cost-effective and environment-friendly. However, further studies are required to scale up the 6-APA biosynthesis reaction for achieving 80-90% conversion of penicillin G into 6-APA by PGA hyper-producing locally collected strains of E. coli.
PDF Abstract XML References Citation
How to cite this article
Rubina Arshad, Shafqat Farooq and Syed Shahid Ali, 2007. 6-Aminopenicillanic Acid Production by Intact Cells of E. coli Containing Penicillin G Acylase (PGA). Pakistan Journal of Biological Sciences, 10: 3190-3194.
DOI: 10.3923/pjbs.2007.3190.3194
URL: https://scialert.net/abstract/?doi=pjbs.2007.3190.3194
DOI: 10.3923/pjbs.2007.3190.3194
URL: https://scialert.net/abstract/?doi=pjbs.2007.3190.3194
INTRODUCTION
Penicillin G acylase (PGA): A microbial enzyme catalyzes the deacylation of penicillin with formation of side chain acid (Shewale et al., 1990) and 6-Aminopenicillanic acid (6-APA), which is an important commercial intermediate in the industrial production of β-lactam antibiotics such as ampicillin and amoxicillin (Shewale and SivaRaman, 1989). More than 60% of 6-APA is produced enzymatically (Shewale and SivaRaman, 1989). Immobilized enzymes and cells have been used in the pharmaceutical industry to produce 6-APA (Wang et al., 1982; Zhang, 1982; Ishimura and Suga, 1992; Stambolieva et al., 1992; Giordano et al., 2006). Penicillin G acylase can be immobilized either in the form of isolated enzyme or whole cell enzyme by different techniques (Norouzian et al., 2002). Production of 6-APA by enzymatic hydrolysis of penicillin G using immobilized PGA is an environment-friendly, sustainable and efficient process (Wang et al., 2006). In batch reaction, the enzymatic conversion yield of penicillin G to 6-APA using whole cells immobilized system is about 75% (Cheng et al., 2006). Water two-phase system for 6-APA production is more beneficial than aqueous batch systems used widely in industrial production of 6-APA (Zhang et al., 2006). The aims of present study are therefore, to explore the feasibility of producing 6-APA using intact cells of different locally collected PGA producing E. coli strains as cheaper biocatalyst and to optimize conditions for cost-effective production of 6-APA.
MATERIALS AND METHODS
Bacterial strains and culture conditions: Penicillin G acylase producing bacterial strains (Table 1) obtained from BDCS-NIAB (Arshad et al., 2006) and foreign culture collection (ATCC) were cultivated into 250 mL Erlenmeyer flask containing 100 mL fermentation medium of the following composition: Corn steep liquor (CSL) 1.25%, ammonium sulphate 0.1%, phenylacetic acid (PAA) 0.15%.
| Table 1: | Bacterial strains used |
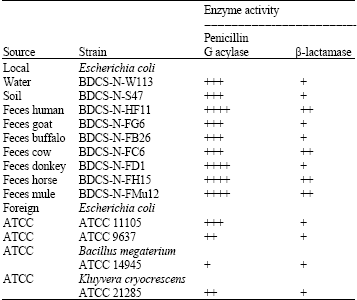 | |
| + fair, ++ moderate, +++ good, ++++ excellent, ATCC American Type Culture Collection | |
The final pH of medium was adjusted to 6.5 with I M NaOH. Fermentation medium was incubated at 28°C for 18 h on a rotary shaker (150 rpm). Bacterial cells from overnight grown culture were separated by centrifugation at 5,000 x g at 4°C for 10 min., washed and suspended in 0.05 M phosphate buffer, pH 7.5 (50 mg wet cells mL-1).
Enzymatic hydrolysis of penicillin G: One millilitre cell suspension of each bacterial culture was mixed with 5 mL of 0.05 M phosphate buffer (pH 7.5) containing 25 mg penicillin G. The reaction mixture was incubated at 40°C for 1-4 h, samples were taken hourly up to 4 h to monitor the bioconversion of penicillin G into 6-APA by p-dimethylaminobenzaldehyde (PDAB) method (Saba et al., 1998). The amount of 6-APA produced was estimated by spectrophotometer using 415 nm wavelength.
Isolation process of 6-APA: One gram wet cells were suspended in 20 mL of potassium phosphate buffer, pH 7.5. Penicillin G solution was prepared by dissolving 0.5 g of penicillin G in 100 mL of phosphate buffer, pH 7.5 and was added to cell suspension in a 250 mL Erlenmeyer flask. The reaction mixture (120 mL) was kept shaking (50 rpm) at 40°C for 4 h. Isolation of 6-APA from the reaction mixture: a multi-step process was carried out after a reaction time of 4 h. The reaction mixture was centrifuged, cells were discarded and supernatant was processed for 6-APA isolation. The pH of supernatant was adjusted to 2.0 with 6 M H2SO4. The PAA and residual penicillin G were extracted from the reaction mixture at pH 2.0 with same volume (120 mL) of n-butylacetate. After phase separation, the pH of aqueous phase was adjusted to 7.0 with Ba(OH)2 and centrifuged. After centrifugation, precipitates were discarded, supernatant was concentrated to 20 mL by evaporation under vacuum (< 30°C), 40 mL of methanol was added before it was filtered after keeping the mixture overnight in refrigerator. Filtrate was re-concentrated (5 mL) under vacuum and precipitated by adjusting the pH to 4.3 with 6 M HCl. The 6-APA crystals were washed with cold water and dried up under vacuum.
RESULTS AND DISCUSSION
Penicillin G acylase activity in locally collected E. coli strains ranged from 3.1-6.4 mg 6-APA h-1 mg-1 wet cells, whereas comparatively low levels of enzyme activity (2.4-4.7 mg 6-APA h-1 mg-1 wet cells) were recorded in foreign strains (Table 2). The maximum PGA activity was observed in BDCS-N-FMu12 (6.4 mg 6-APA h-1 mg-1 wet cells) followed by BDCS-N-FD1 (5.7 mg 6-APA h-1 mg-1 wet cells). Among the foreign strain B. megaterium (ATCC 14945) exhibited the minimum enzyme activity (2.4 mg 6-APA h-1 mg-1 wet cells), while remaining three locally collected E. coli strains BDCS-N-FMu12, BDCS-N-HF11 and BDCS-N-FH15 showed enzyme activity in between the two. The PGA producing fermentation medium used in the present study was supplemented with 1.25% corn steep liquor and 0.15% phenylacetic acid because this enhances the production of PGA as reported earlier (Arshad et al., 2000). We carried out acylation reaction at pH 7.5 which is slightly lower than (pH 7.8) reported by Lagerlof et al. (1976).
The steps involved in the enzymatic conversion of penicillin G into 6-APA include isolation process comprising enzyme reaction mixture incubation in a rotary shaker set for 50 rpm for 4 h at 28°C, centrifugation of reaction mixture, acidic pH adjustment of supernatant, supernatant extraction for phase separation, pH adjustment of aqueous phase containing 6-APA, supernatant concentration under vacuum, concentrate treatment with methanol and finally 6-APA crystallization by pH adjustment to 4.3. White crystals of 6-APA were obtained at the completion of this process (Fig. 1). The production of 6-APA is a multi-step process involving fermentation, acylation, extraction, concentration, crystallization, filtration, washing, and drying in which penicillin acylases may be used as purified enzymes (Balasingham et al., 1972; Schömer et al., 1984) or in immobilized form (Nabais and Cardoso, 2000; Giordano et al., 2006).
| Table 2: | Penicillin G acylase activity in various bacterial strains and enzymatic conversion of penicillin G into 6-aminopenicillanic acid (6-APA) |
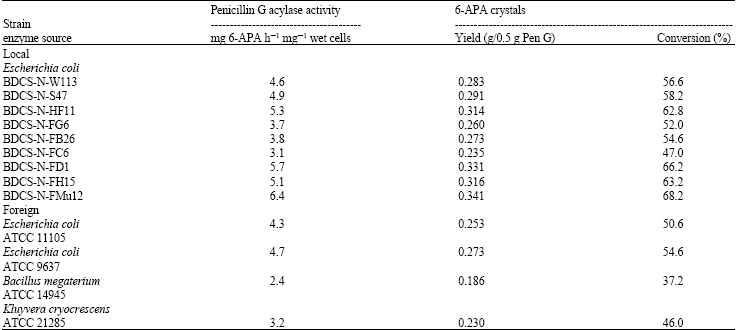 | |
| 6-APA: 6-aminopenicillanic acid; Pen G: penicillin G | |
 | |
| Fig. 1: | Extracted 6-APA in crystalline form |
The process optimized during the present study involved penicillin G hydrolysis by flask culture using cell-bound PGA of locally collected E. coli strains followed by extraction of 6-APA as a crystalline product. Since cell-bound enzyme lowers operational cost as compared to purified free/immobilized enzyme therefore, crude cell suspensions were used in this study. Phenylacetic acid (PAA): a strong acid, formed during enzymatic hydrolysis of penicillin G to 6-APA, is required to be removed prior to crystallization of 6-APA by extraction with organic solvent. For this purpose we used n-butylacetate at pH 2.0. Acetone (Boehringer Mannheim, 1985), methylene chloride (Rohm Pharma, 1987), butanol (Shewale and SivaRaman, 1989) and methyl isobutyl ketone (Harrison and Gibson, 1984) have also been used for PAA extraction. Among these solvents, methyl isobutyl ketone is considered a hazardous air pollutant (Traister, 2000) whereas n-butylacetate is nearly three times less toxic than methyl isobutyl ketone. Using n-butylacetate as the extraction solvent rather than methyl isobutyl ketone is therefore, pollution free.
The filtrate, containing 6-APA crystals, was concentrated in vacuum at low temperature (> 30°C) and a pH of 7.0 which is necessary to avoid degradation of β-lactam ring. The concentrated aqueous solution was finally precipitated at 2-5°C, pH 4.2-4.3 in the presence of methanol and 6-APA was then isolated by crystallization under acidic conditions. Recently, Gong et al. (2005), have reported an iso-electric reactive crystallization to obtain 6-APA crystals at different physical conditions of the solutions however, in the present study, crystallization was carried out by a stepwise adjustment of the pH of 6-APA-solution within a range of 2.5-4.5 using hydrochloric acid. This is necessary to avoid contamination in 6-APA with high side chain molecules. Methanol is also added because under low concentrations in the reaction medium it exerts complex and interesting changes in the catalytic properties of immobilized PGA (Fernandez-Lafuente et al., 1998).
The overall yield of 6-APA obtained using the procedure standardized in the present study was around 37-68%. The maximum conversion of penicillin G into 6-APA was observed in E. coli strain BDCS-N-FMu12 in which one gram of wet cells converted 0.5 g of penicillin G into 0.341 g of 6-APA. BDCS-N-FMu12 was identified as the best PGA producer with 68% conversion of penicillin G into 6-APA whereas B. megaterium ATCC 14945 showed the lowest conversion (37%). The recovery percentages achieved in foreign E. coli strains ATCC 11105 and ATCC 9637 were 51 and 55%, respectively. The yield of 6-APA crystals obtained in local and foreign strains ranged from 47-68 and 37-55%, respectively. Only one local strain BDCS-N-FC6 showed less than 50% conversion. The results indicated that local strains were relatively better for biosynthesis of 6-APA.
In the present study, extraction and crystallization, were carried out in shake-flask scale therefore, 6-APA crystals were isolated by filtration and drying under vacuum. It was observed that maximum amount of 6-APA and PAA was removed by a washing which together with reaction solution increase product losses in mother liquor (about 4% 6-APA remains in the mother liquor). Nevertheless, we have obtained approximately 37-68% yield of 6-APA using intact cells of locally collected E. coli strains (Arshad et al., 2006) as the enzyme source.
We have observed that if all stages of extraction procedure are well controlled, time and temperature of process kept low and pH is adjusted carefully, a yield of about 80-90% can then be achieved which is close to 6-APA production (90% conversion of penicillin G to 6-APA) using cross-linked enzyme aggregates of Bacillus badius PGA (Rajendhran and Gunasekaran, 2007).
The data reported in the present study suggested that locally collected E. coli strains are good producers of PGA. Although, the expression of the intracellular PGA activity occurs under several regulatory controls such as induction, catabolic repression, temperature, pH, oxygen levels etc. (Babu and Panda, 1991; Liu et al., 2000) yet it is free from hazardous chemicals, is environmental safe and using intact whole cells as PGA biocatalyst could be an efficient approach for low cost production of 6-APA. The process can be further enhanced by removing the coexistence of β-lactamase with PGA (Arshad and Saba, 2001), the major factor underlying the low production of 6-APA. In the present study, the recovery percentage of 68% using crude cell suspension (having coexistence of both PGA and β-lactamase) is quite promising which can reach to 80-90% if β-lactamase is removed by mutagenesis.
REFERENCES
- Cheng, S., D. Wei, Q. Song and X. Zhao, 2006. Immobilization of permeabilized whole cell penicillin G acylase from Alcaligenes faecalis using pore matrix crosslinked with glutaraldehyde. Biotechnol. Lett., 28: 1129-1133.
CrossRefDirect Link - Fernandez-Lafuente, R., C.M. Rosell and J.M. Guisan, 1998. The presence of methanol exerts a strong and complex modulation of the synthesis of different antibiotics by immobilized penicillin G acylase. Enz. Microb. Technol., 23: 305-310.
CrossRefDirect Link - Giordano, R.C., M.P. Ribeiro and R.L. Giordano, 2006. Kinetics of betalactam antibiotics synthesis by penicillin G acylase (PGA) from the viewpoint of the industrial enzymatic reactor optimization. Biotechnol. Adv., 24: 27-41.
PubMedDirect Link - Liu, Y.C., L.C. Liao and W.T. Wu, 2000. Cultivation of recombinant Escherichia coli to achieve high cell density with a high level of penicillin G acylase activity. Proc. Natl. Sci. Counc. Repub. China B., 24: 156-160.
PubMed - Nabais, A.M.A. and J.P. Cardoso, 2000. Enzymatic conversion of benzylpenicillin to 6-aminopenicillanic acid in concentrated ultrafiltered broths. Bioproc. Biosyst. Eng., 23: 191-197.
Direct Link - Norouzian, D., S. Javadpour, N. Moazami and A. Akbarzadeh, 2002. Immobilization of whole cell penicillin G acylase in open pore gelatin matrix. Enz. Microb. Technol., 30: 26-29.
CrossRefDirect Link - Wang, L., W. Zhilong, J.H. Xu, D. Bao and H. Qi, 2006. An eco-friendly and sustainable process for enzymatic hydrolysis of penicillin G in cloud point system. Bioproc. Biosyst. Eng., 29: 157-162.
CrossRefDirect Link - Zhang, W.G., D.Z. Wei, X.P. Yang and Q.X. Song, 2006. Penicillin acylase catalysis in the presence of ionic liquids. Bioproc. Biosyst. Eng., 29: 379-383.
CrossRefDirect Link