
M. Badri
Departrnent of Biology, Faculty of Sciences, Razi University, Kermanshah, Islamic Republic of Iran
M. R. Zamani
National Institute for Genetic Engineering and Biotechnology (NIGEB), Tehran, Islamic Republic of Iran
M. Motallebi
National Institute for Genetic Engineering and Biotechnology (NIGEB), Tehran, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 17 | Page No.: 2850-2855







ABSTRACT
In this study the effect of two plant growth regulators (indolacetic acid, IAA and gibberellic acid, GA3) and also Trichoderma harzianum (T8) on the phytopathogen fungus Fusarium oxysporium (F15) was investigated. IAA and GA3 with 15 and 30 ppm concentration have no significant effect on T. harzianum (T8) growth. The biocontrol activity of T. harzianum on F. oxysporum was slightly decreased by the presence of IAA and/or GA3. Addition of 40 ppm of GA3 to the culture medium of F. oxsporum increased polygalacturonase activity about 100%. A strong increasing effect on chitinase activity (60%) by T. harzianum (T8) was observed in the presence of phytopathogenic fungus F. oxysporum, but 40 ppm IAA and/or GA3 decreased about 47% of chitinase activity of T. harzianum.
PDF Abstract XML References Citation
How to cite this article
M. Badri, M. R. Zamani and M. Motallebi, 2007. Effect of Plant Growth Regulators on in vitro Biological Control of Fusarium oxysporum by Trichoderma harzianum (T8). Pakistan Journal of Biological Sciences, 10: 2850-2855.
DOI: 10.3923/pjbs.2007.2850.2855
URL: https://scialert.net/abstract/?doi=pjbs.2007.2850.2855
DOI: 10.3923/pjbs.2007.2850.2855
URL: https://scialert.net/abstract/?doi=pjbs.2007.2850.2855
INTRODUCTION
Fusarium wilt caused by Fusarium oxysporum is regarded as the most important disease of chickpea (Cicer arietinum) in Iran (Zamani et al., 2001). The disease is characterized by two syndromes, namely vascular wilt and yellowing, which can be distinguished by both symptomatology and chronological development. The wilt syndrome results in a rapid flaccidity and dessication of leaves and stems by 20 days after inoculation, whereas the yellowing syndrome results in a progressive foliar yellowing followed by necrosis 35-40 days after inoculation (Trapero-Casas and Jimi Nez-Diaz, 1985). The pathogenic mechanisms of F. oxsporum are complex and varied and may be due to the action of different pectolytic enzymes that produce modification of cell wall structure, increasing accessibility of cell wall components for degradation by other enzymes, cell lysis and plant tissue maceration (Perez-Artez and Tena, 1989).
Control of plant disease is vital in plant agriculture (Ekelund et al., 2000). Commercially produced, registered products, such as fungicides, are frequently recommended for plant disease management. These chemicals have an established history of controlling many economically important diseases, despite the development of tolerance or resistance in pest organisms (Godet and Limpert, 1998; Gossen and Rimmer, 2001). In most countries, products used for disease management are subject to tests of efficacy and assessments of risks to non-target organisms. This risk can be defined as the combination of the agent’s toxicity and the probability of exposure of non-target organisms (Cook et al., 1996; Hintz et al., 2001).
Intensified use of fungicides has resulted in the accumulation of toxic compounds potentially hazardous to humans and environments. In order to tackle these national and global problems, effective alternatives to chemical control are being investigated and the use of antagonistic microbes seems to be one of the promising approaches (Lorito et al., 1998). Antagonism may be accomplished by competition, parasitism, or by a combination of these modes of action (Lorito et al., 1998; Samuels, 1996).
Chitin and β-1, 3 glucan are the two major structural components of the cell wall of many plant pathogenic fungi. Therefore, it is expected that the 1, 3 glucanases, chitainases and proteases produced extracellulary by soil fungus Trichoderma play an important role in biocontrol (Harighi et al., 2006, 2007; Chye et al., 2005; Geremia et al., 1993). Some species of Trichoderma sp. have been shown to act as mycoparasites against a range of economically important aerial and soil-borne plant pathogens, being successfully used in the field and greenhouse (Chet, 1987; Dubos, 1987; Papavizas, 1985).
The interaction between Trichoderma sp. and its host is first detectable as chemotropic growth of hyphae of the mycoparasite toward its host (Carsolio et al., 1994).
After interaction the mycoparasite penetrates the host mycelium, apparently by partially degrading its cell wall by producing mycolytic enzymes (Carsolio et al., 1994).
Biocontrol activity of Trichoderma sp. may be affected by environmental conditions such as the presence of plant nutrients at the field level, which contain growth regulators. The presence of growth regulators in the soil could come from various ways including treatments of foliar portion of plants or fruits and tubers or from soil microorganisms (Agrios, 1997). Some fungal pathogens and bacteria such as Agrobacterium tumefaciens and Pseudomonas savastanoi code for indolacetic acid (IAA) synthesis. Therefore, its seemed important to analyze if plant growth regulators could affect the biocontrol activity of Trichoderma sp. The activity of endopolygalacturonase of fungi is known to be affected by fungicides, environmental conditions and foliar nutrients (Aub et al., 1993) which may contain growth regulators that could also affect the activity of this enzyme.
In this study it was decided to use F. oxysporum as the target microorganism to test the biocontrol activity of T. harizianum (T8) in the presence or absence of phytohormones which would allow us to improve biocontrol conditions.
MATERIAL AND METHODS
Fungal isolates and growth conditions: A highly virulent isolate of F. oxyspurum (F15) from North West of Iran (Zamani et al., 2001; Motallebi et al., 2002) and T. harizianum (T8) isolate were used throughout this study. The isolates were maintained in Potato Dextrose Agar (PDA) medium at 4<°C.
Enzyme production: For polygalacturonase production, F. oxysporum, was grown in shake culture on Pectic Zymogram (PZ) medium containing 2.64 g (NH4)2 SO4, 0.34 g KH2PO4, 0.14 g MgSO4. 7H2O, 10 g Citrus pectin, 1 Lit. dH2O. pH adjusted to 4.5 (Sweetingham et al., 1986). After 6 days growth at 26<°C, the liquid culture filtrate was obtained (as crude enzyme), by Whatman filter paper No. 1 and stored at -20<°C until using for enzyme assay or protein measurement.
For chitinase production, T. harizianum was grown on SM medium containing 0.6 g Urea, 4 g KH2PO4, 2.8 g (NH4)2 SO4, 0.6 g CaCl2.2H2O, 0.2 g MgSO4, 0.01 g FeSO4.7H2O, 2.8 mg ZnSO4.H2O and 3.2 mg CoCl2.6H2O supplemented with 0.5% of crab shell chitin (Sigma) as carbon source. The medium was inoculated with a spore suspension to give a final concentration of 5x106 conidia mL-1 and placed on rotary shaker at 150 rpm at 25<°C for 4 days and the liquid culture filtrate was obtained (as crude enzyme), by Whatman filter paper No. 1 and stored at -20<°C until using for enzyme assay.
Enzyme assay: Polygalacturonase (PG) activity was assayed by measuring the release of the reducing groups using the Somogi assay with Nelson’s arsenomolibdate reagent (Collmer et al., 1988). The reaction mixture, containing 0.9 mL of 2.5% polygalacturonic acid in 25 mm citrate-phosphate buffer pH 4.5 and 0.1 mL of enzyme solution, was incubated at 40<°C for 20 min. One unit of polygalacturonase activity was defined as the amount of enzyme that releases 1 μmol of galacturonic acid per minute.
Chitinase activity was assayed using the colorimetric method described by Rojas-Avelipaza et al. (1999) with minor modifications. The assay mixture contained 200 μL of 0.5% pure chitin (sigma) in 50 mm acetate buffer pH = 6 and 200 μL of enzyme solution. The reaction mixture was incubated for 60 min at 40<°C and was stopped by centrifugation (6000 rpm) for 5 min and the addition of 1 mL of dinitrosalicylate (DNS) reagent. One unit of chitinase activity was defined as the amount of enzyme that releases 1 μmol of N-acetylglucosamine per hour.
Mycelium growth assessment: Disks (5 mm diameter) from culture of F. oxysporum or T. harizianum were seeded in a PDA medium with or without the addition of 15, 30 and 40 ppm indolacetic acid (IAA) or gibberellic acid (GA3). Fungi were grown up to one week at 28<°C. Colony diameter was recorded every two days. Results correspond to the mean of seven different experiments run in duplicates. Data was analyzed through the test of student at p<0.05.
Conidia germination assessment: Conidia from F. oxysporum or T. harizianum were obtained as described by Sneh (1981). One hundred conidia from each fungus were grown on 2% (w/v) water agar (WA) for 11 h in the absence or presence of 40 ppm GA3 or IAA at 25<°C. Results correspond to the mean of seven different experiments run in duplicates. Data was analyzed through the test of student at p<0.05.
Production of PG and chitinase in submerged culture: The secretion of the enzymes was evaluated in the presence of 40 ppm IAA and/or GA3 in submerged cultures with the addition of pectin or chitin for production of polygalacturonase or chitinase, respectively. Flasks, containing 200 mL of any of the above media were inoculated with 1x106 conidia of either F. oxysporum or T. harizianum. To test the effect of T. harizianum on the ability of F. oxysporum to secrete polygalacturonase, conidia of each fungus were inoculated in the pectin-containing medium. To test the effect of F. oxysporum on the ability of T. harizianum to secret chitainase, conidia of each fungus were inoculated in the chitin-containing medium. Flasks were incubated at 28<°C up to 2 weeks with shaking at 150 rpm. The whole medium was then centrifuged at 7000 rpm for 10 min to remove mycelia and the supernatant was used to test polygalacturonase or chitinase activity. Results correspond to the mean of three independent experiments run in triplicates. Controls were performed with heat inactivated conidia. This study was carried out in Plant Biotechnology Laboratory on National Institute for Genetic Engineering and Biotechnology (NIGEB), Tehran, Islamic Republic of Iran.
RESULTS
Evaluation of mycelial growth and conidia germination: Out of 30 Trichoderma sp. isolates T. harzianum, the best chitinase producer, (T8) (Seyed Asli et al., 2004) selected and used for study of biocontrolling effect on a phytopathogenic isolate of F. oxysporum. This fungal isolate of F. oxysporum (F15) had been classified among highly virulent isolates, on the basis of bioassay test (Zamani et al., 2001).
In this study the effect of plant growth regulators IAA and GA3 at concentration of 15, 30 and 40 ppm on fungal growth of F. oxysporum (F15) and T. harzianum (T8) was investigated. The results showed that the 32.5 mm growth of (colony diameter) F. oxysporum on PDA after three days, was not altered by the presence of IAA and/or GA3 at the concentrations tested (Table 1). On the other hand, 15 and 30 ppm of IAA and GA3 either alone or in combination have no significant effect on T. harzianum growth, but 40 ppm of IAA and/or GA3 slightly decreased the T. harzianum colony diameter (Table 1).
| Table 1: | Effect of IAA and GA3 on mycelial growth of F. oxysporum and T. harzianum in independent culture |
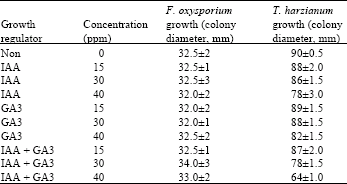 | |
The in vitro biocontrol activity of T. harzianum on F. oxysporum (about 31%) was slightly decreased by the presence of IAA and/or GA3 in 15, 30 and 40 ppm (Fig. 1). To demonstrate the effect of IAA and/or GA3 on conidial germination of F. oxysporum (F15) and T. harzianum (T8), 40 ppm concentration of these two growth regulators was added to their independent culture medium. The results indicated that in the absence of the plant growth regulators, germination of conidia after 11 h in F. oxysporum and T. harzianum were 49% (Fig. 2) and 83% (Fig. 3), respectively. Addition of 40 ppm of IAA reduced germination of conidia in F. oxysporum and T. harzianum by 76 and 48%, respectively (Fig. 2 and 3). The same concentration of GA3 did not affect significantly the germination of conidia in these fungi (Fig. 2 and 3), while concentration of 40 ppm of GA3 and IAA in combination reduced conidial germination by 35 and 22% in F. oxysporum (F15) and T. harzianum (T8), respectively.
Evaluation of polygalacturonase and chitinase in submerged culture: To evaluate the effect of the presence of T. harzianum (T8) in submerged culture on polygalacturonase activity of F. oxysporum, an equal number of conidia of these two fungi were used. The results showed 55% inhibitory effect on polygalacturonase activity in the presence of T. harzianum (T8) (Fig. 4) which may be explained as a consequence of the growth inhibitory effect of the biocontrol agent on F. oxysporum (F15).
The effect of 40 ppm GA3 and/or IAA on the secretion of polygalacturonase from F. oxysporum (F15) in the presence of T. harzianum was investigated in submerged cultures. The obtaining results indicate that while the addition of 40 ppm of GA3 to the culture medium of F. oxysporum increased polygalacturonase activity about 100% (Fig. 4), the addition of 40 ppm of IAA either alone or in combination with 40 ppm of GA3 did not affect significantly polygalacturonase activity of this fungus (Fig. 4).
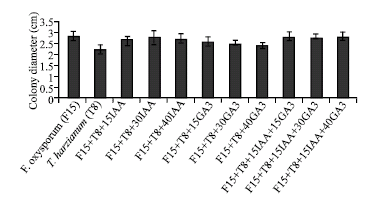 | |
| Fig. 1: | Effect of different concentration of IAA and/or GA3 in the presence of T. harzianum (T8) on mycelia growth of F. oxysporum (F15) |
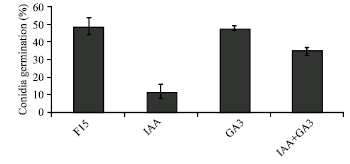 | |
| Fig. 2: | Effect of 40 ppm of IAA and/or GA3 on conidial germination of F. oxysporum (F15) |
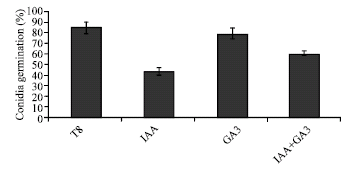 | |
| Fig. 3: | Effect of 40 ppm of IAA and/or GA3 on conidial germination of T. harzianum (T8) |
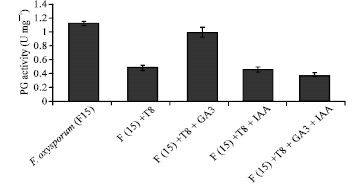 | |
| Fig. 4: | Effect of 40 ppm IAA and/or GA3 and T. harzianum (T8) on production of polygalacturonase from F. oxysporum (F15) |
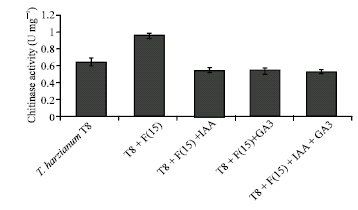 | |
| Fig. 5: | Effect of 40 ppm IAA and/or GA3 and F. oxysporum (F15) on production of chitinase enzymes from T. harzianum (T8) |
To investigate the effect of the presence of F. oxysporum on chitinase activity of T. harzianum, the chitinase activity was assayed in submerged culture medium. A strong increasing effect on chitinase activity (60%) was observed in the presence of phytopathogenic fungus, F. oxysporum (F15) (Fig. 5), which may be explained as a consequence of inducing effect of the F. oxysporum on chitinase production of T. harzianum (T8).
To evaluate the effect of plant growth regulators on chitinase activity of T. harzianum (T8), 40 ppm IAA and/or GA3 was added to the culture medium of this fungus in the presence of F. oxysporum. The chitinase activity was assayed after 14 days of incubation. The results demonstrated that 40 ppm IAA and GA3 either alone or in combination decreased about 47% of chitinase activity of T. harzianum (T8) (Fig. 5).
DISCUSSION
The mycoparasitic Trichoderma spp. are important biocontrol agents against a variety of phytopathogenic fungi (Howell, 1998; Harighi et al., 2006, 2007). One of the major mechanisms involved in the biocontrol activity of members of the Trichoderma is mycoparasitism (Haran et al., 1996; Lorito et al., 1998; Wang et al., 2003). In order to be able to degrade phytopathogen cell walls, the Trichoderma induces the production of extracellular hydrolytic enzymes, responsible for the direct attack against the pathogen. These enzymes have been reported mainly in isolates of T. harzianum (Lorito et al., 1993, 1994; Haran et al., 1996; Lorito et al., 1998; Chye et al., 2005). Nevertheless, the biocontrol activity of T. harzianum could be affected by environmental cues, that include among others, the presence of phytopathogenic fungi and plant growth regulators such as IAA and GA3. The presence of growth regulators in the soil could come from applications done to the foliar portion of plants or to fruits, where part of applications fall either directly or from plants into the soil (Roco and Perez, 2001). It has been described that some fungal pathogens are able not only to induce increased levels of IAA in their respective hosts, but are themselves capable of producing IAA which is directly released into the soil (Agrios, 1997).
In this study the in vitro biocontrol activity of T. harzianum (T8) on the phytopathogenic fungi F. oxysporum was investigated in the presence and absence of the growth regulator IAA and GA3. While these regulators showed to have no significant effect on mycelium growth rate of F. oxysporum, the mycelium growth of T. harzianum was decreased in the presence of these regulators.
The maximal secretion of polygalacturonase from F. oxysporum into submerged cultures was reduced about 56% when this phytopathogen was grown in the presence of T. harzianum (T8). Although the presence of GA3 in this medium increased the polygalacturonase activity, but IAA showed to have no significant effect on the polygalacturonase secretion in the medium. These results suggest that GA3 may interfere the secretion process of the enzyme or its levels, which is confirmed by the finding of Al-Masri et al. (2002).
The addition of 40 ppm of GA3 and/or IAA in the presence of F. oxysporum did not significantly affect the ability of T. harzianum (T8) to secrete chitinase enzymes. The addition of F. oxysporum in the absence of growth regulators into submerged culture of T. harzianum (T8) increased the level of chitinase activity about 58%, suggesting that the presence of the pathogen could serve as an inducer of this fungal cell wall degrading enzymes. Then, the presence of this pathogen would stimulate one of the mechanisms which is used by T. harzianum for its biocontrol activity. The different expression of isoenzymes of chitinase by Trichoderma has been described during the mycoparasitism of this fungus on pathogens (Haran et al., 1996), thus accounting for its antagonism against several pathogens.
CONCLUSION
In conclusion, the presence of phytopathogen fungus (F. oxysporum) in submerged culture of T. harzianum (T8) in the absence of growth regulators increases the ability of T. harzianum to control F. oxysporum by increasing the chitinase production. On the other hand, the presence of plant growth regulators in T. harzianum culture medium decrease both the chitinase production and mycelia growth of T. harzianum which could decrease the mycoparasitim of this fungus on phytopathogens.
REFERENCES
- Al-Masri, M.I., M.S. Ali-Shtayeh, Y. Elad, A. Sharon, P. Tudzynski and R. Barakat, 2002. Effect of plant growth regulators on white mould (Sclerotinia sclerotiorum) on bean and cucumber. J. Phytopathol., 150: 481-487.
CrossRefDirect Link - Chye, M.L., K.J. Zhao, Z.M. He, S. Ramalingam and K.L. Fung, 2005. An agglutinating chitinase with two chitin-binding domains confers fungal protection in transgenic potato. Planta, 220: 717-730.
CrossRefPubMedDirect Link - Cook, R.J., W.L. Bruckart, J.R. Coulson, M.S. Goettel and R.A. Humber et al., 1996. Safety of microorganisms intended for pest and plant disease control: A framework for scientific evaluation. Biol. Control, 7: 333-351.
CrossRef - Ekelund, F., K. Westergaard and D. Soe, 2000. The toxicity of the fungicide propiconazole to soil flagellates. Biol. Fertil. Soil, 31: 70-77.
Direct Link - Harighi, M.J., M.R. Zamani and M. Motallebi, 2007. Evaluation of antifungal activity of purified chitinase 42 from Trichoderma atroviride PTCC5220. Biotechnology, 6: 28-33.
CrossRefDirect Link - Papavizas, G.C., 1985. Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Ann. Rev. Phytopathol., 23: 23-54.
Direct Link - Perez-Artez, E. and M. Tena, 1989. Pectic enzymes from two races of Fusarium oxysporum f. sp. Ciceri, enzyme production in culture and enzymatic activity on isolated chickpea cell wall. J. Phytopathol., 124: 39-51.
Direct Link - Wang, Y., A.P. Kausch, J.M. Chandlee, H. Luo and B.A. Ruemmele et al., 2003. Co-transfer and expression of chitinase, glucanase and bar genes in creeping bentgrass for conferring fungal disease resistance. Plant Sci., 165: 497-506.
Direct Link