
Azra Saboora
Department of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Khadijeh Kiarostami
Department of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Fatemeh Behroozbayati
Department of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Shokoofeh Hajihashemi
Department of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 11 | Page No.: 2009-2021







ABSTRACT
Salinity tolerance during germination and early seedling growth was evaluated for nine wheat cultivar (Triticum aestivum L.) in six treatments of salinity including 0 (control), 75, 150, 225, 300 and 375 mM NaCl in a 3 replicated RCBD. The results showed that different treatments of salinity had considerable effect on the germination percentage, germination rate, elongation of the first leaf and root, total dry weight and root/shoot dry weight. Germination percentage in all cultivar showed considerable decrease with increasing salinity up to 375 mM NaCl. This reduction was more in Pishtaz and Ghods as compared to Tajan and Karchia. The seedling growth of nine genotypes was significantly inhibited by all salinity levels. During early seedling growth, applied NaCl salinity significantly affected the RGR of all the genotypes, particularly at 150-375 mM NaCl but there was no significantly difference at the low salinity treatments (p<0.05). The effect of salinity on RGR values varied according to the salt tolerance of the genotypes. First leaf and root length of all genotypes were considerably reduced. Elongation of the first leaf was more affected as compared to root growth at all salinity levels. Also, the rate of reducing of total dry matter was slower than reducing of leaf elongation. Cluster analysis with multiple parameters simultaneously to evaluate the salt tolerance revealed three groups. Karchia was used as a standard for the salt tolerance test. Among 9 genotypes, Tajan was the most tolerant and Pishtaz and Ghods were the most sensitive genotypes.
PDF Abstract XML References Citation
How to cite this article
Azra Saboora, Khadijeh Kiarostami, Fatemeh Behroozbayati and Shokoofeh Hajihashemi, 2006. Salinity (NaCl) Tolerance of Wheat Genotypes at Germination and Early Seedling Growth. Pakistan Journal of Biological Sciences, 9: 2009-2021.
DOI: 10.3923/pjbs.2006.2009.2021
URL: https://scialert.net/abstract/?doi=pjbs.2006.2009.2021
DOI: 10.3923/pjbs.2006.2009.2021
URL: https://scialert.net/abstract/?doi=pjbs.2006.2009.2021
INTRODUCTION
Soil salinity is one of the most important factors that limit crop production in arid and semi arid regions (Neumann, 1995). Salinity affects about 7% of the world’s total lands area (Flowers et al., 1997). The percentage of cultivated land affected by salt is even greater, comprises 19% of the 2.8 billion hectares of arable land on earth (Ponnamierumo, 1984; Pessarakli and Szabolcs, 1999; El-Hendawy et al., 2004). Furthermore there is also a dangerous trend of a 10% per year increase in the saline area throughout the world. Soil salinity may be robbing the country of about 25% of its crop production. A major part of the salt-affected soils, about 3.5 million hectares is under rice, wheat, cotton, sugarcane and rapeseed cultivation (Raza, 2005). Wheat is a moderately salt tolerant crop and serves as a staple food in 43 countries (Pervaiz et al., 2002; Raza, 2005); including Iran, where it is grown on a large area. On the other hand, Iran is one of the countries that suffer from severe salinity problems. For example 18 M ha or 10% of total land area in Iran is salinity or sodicity soil.
Most crop plants are glycophytes, which have evolved under low salt condition. The mechanisms they have evolved for uptake, transport, recirculation and utilization of minerals may not function optimally under saline conditions. Salinity decreases germination (Murillo-Amador et al., 2000a; Gehlot et al., 2005; Sharma et al., 2004), dry matter accumulation, the rate of net CO2 assimilation, relative growth, leaf cell expansion and ultimate leaf growth (Murillo-Amador et al., 2000b; Cramer et al., 2001; Saqib et al., 2004; Mansour et al., 2005). The plant growth is ultimately reduced by salinity stress but plant species have different responses to salinity. Salt tolerance of crops may vary with their growth stage (Munns and Termaat, 1986; Mass et al., 1994; Rogers et al, 1995; Flowers et al., 1997).
The establishment stage of the crop consists of three parts: germination, emergence and early seedling growth; that are particularly sensitive to substrate salinity (Mariko et al., 1992; Baldwin et al., 1996; Grieve and Suarez, 1997; Jamil et al., 2005; Raza, 2005). Successful seedling establishment depends on the frequency and the amount of precipitation as well as on the ability of the seed species to germinate and grow while soil moisture and osmotic potentials decrease (Welbaum et al., 1990; Roundy, 1985). Much information is available in literature about the effects of water quality, soil texture and soil salinity on germination and emergence (Grillot, 1957; Maas, 1986; Jamil and Rha, 2004; Jamil et al., 2005). Retardation and reduction in seed germination have been reported under NaCl treatments in the literatures (Sharma et al., 2004; Gill et al., 2003, Garrciarrubio et al., 2003). The decrease in germination rate particularly under drought and salt stress conditions may be due to the fact that seeds seemingly develop an osmotically enforced dormancy under water stress conditions. This may be an adaptive strategy of seeds to prevent germination under stressful environment thus ensuring proper establishment of the seedlings (Gill et al., 2003). During early seedling growth, salinity and soil texture affected the development of the seedlings that showed symptoms of water stress. The consequence of water stress could already be observed some days after emergence, the higher water stress, expressed by lower leaf water potential, stomatal conductance and evapotranspiration and the lower the leaf area and dry matter production (Katerji et al., 1994; El-Hendawy et al., 2005).
Salinity stress is an important characteristic when selecting a variety for salinity tolerance (Konak et al., 1999). Numerous traits related to salt tolerance that have been used to screen germplasm include germination percentage, seedling root and shoot attributes, rates of Na+ or Cl¯ accumulation in leaves (Munns et al., 1995, 2006), ion concentration in root cells (Flowers and Hajibagheri, 2001; Munns et al., 2006). Munns et al. (2006) discussed physiological mechanisms and selectable indicators of gene action, with the aim of promoting new screening methods to identify genetic variation for increasing the salt tolerance of cereal crops, particularly with respect to wheat. They reported that precise phenotyping is the key to finding and introducing new genes for salt tolerance into crop plants.
Furthermore, Growth rate is a key parameter, but the Relative Growth Rate of plants (RGR) under saline condition has been considered to allow more appropriate comparisons of growth among species or genotypes than absolute growth rate (Cramer et al., 1994; Jaradat et al., 2004). The RGR is a function of the Net Assimilation Rate (NAR), which is an index of the photosynthetic-assimilatory capacity of the plant per unit leaf area. At the level of the whole plant, therefore, this parameter may make it possible to clarify whether genotypic variation in salt tolerance can be attributed to morphological changes or photosynthetic response (Ishikawa et al., 1991; El-Hendawy et al., 2005).
Studies on a range of grass species showed that leaf elongation rate reduced by salinity. The relative elemental growth rate of leaf and the length of elongation zone respond in various ways to environmental constraints on the plant (Delane et al., 1982; Bernstein et al. 1993a, b; Hu et al., 2000; Neves-Piestun and Bernstein, 2005). Leaf growth of grasses is of central importance to their development, not only is the expansion of leaves crucial to early seedling establishment by providing a continuous supply of energy and carbon through photosynthesis, it also facilitates development of other organs such as tillers, ears and grains (Hu et al., 2005).
Genetic variation for desirable plant traits is fundamental to any plant improvement program and dictates potential progress. Researchers are trying to get the salt resistant crop on which human’s food depend. Thus, screening for salt-tolerant wheat germplasm is important to determine whether there is a genetic basis for selection and breeding purposes and to whether there are useful genotypes or new genes for tolerance to salt stress. Although there are extensive studies of salinity effects on wheat, research about the effects of salinity on wheat and critical thresholds of responses is still limited at early seedling growth. The present study was undertaken to study the responses of nine wheat cultivars to different levels of salinity (75-375 mM NaCl) and to determine the genotypic variability in their tolerance to salinity both at the germination and seedling stage. For screen the different wheat genotypes, we used multiple physiological parameters.
MATERIALS AND METHODS
Plant material and growth conditions: Nine cultivars of wheat (Triticum aestivum L.), were used in this study. All seeds (Chamran, Ghods, Karchia, Pishtaz, Shahryar, Shahpasand, Shiraz, Tabasi and Tajan variety) were obtained from the Seed and Plant Improvement Institute in Iran. The experiment was carried out in Azzahra University, Tehran, Iran in September 2004. Karchia is the most tolerant wheat genotype and is used as a standard for the salt resistance test of wheat worldwide (Sharma et al., 1994; Ashraf, 2002).
Similar seed size and weight was selected to exclude effect of that on the seedling establishment. Seeds were surface sterilized in 1.5% (v/v) sodium hypochloride for 10 min and thoroughly washed with sterile tap water. Although field screening for salt tolerance has the advantage of testing germplasm under natural conditions, it is less efficient at germination and early growth stages and also it is more expensive than screening under controlled conditions (Shannon and Noble, 1990). For this reasons, fifteen seeds were placed in 10 cm sterile petri dishes on filter paper supplemented with salt solutions. Salt stress was applied by subjecting the seeds to 5 mL of control (no added NaCl), 75, 150, 225, 300 and 375 mM NaCl solutions. Plates were sealed with parafilm and placed under controlled conditions (25±2°C during the day; 16/8 h Light/ dark; irradiance 4500 Lux) Seeds were considered to have germinated when the radicle measured in excess of 2 mm.
Germination Rate and germination percentage: A germination index was calculated for each subpopulation as GR:
Germination Rate =X1/Y1+ (X2-X1)/Y2 + …+ (Xn-Xn-1)/Yn |
Where Xn is the germination percentage on the nth day and Yn is the number of day from first day experiment (Maguire, 1962). The mean germination percentage was calculated from number of seedlings, at 7th day of growth, having axes at least 5 mm long derived from each Petri dish.
Growth parameters: The fresh and dry weight of the shoots and roots, the length of shoots and roots, length of first leaf and coleoptile were measured immediately after the end experiment of stress treatment. The dry weights were measured by drying the shoot and root at 75°C for 48 h, to give a constant weight.
Relative Growth Rate (RGR): Plant growth was evaluated in terms of RGR. Root and shoot growth of the nine genotypes was assessed by measuring dry weights at the 3 day of start and end of the treatments and calculating Relative Growth Rates (RGR). These were estimated as mean values over the time interval T1 to T2 according to Hunt (1990) using the following equations:
where, T and W represent time (days) and total plant dry weight (g) respectively. Subscripts 1 and 2 refer to the values of the variable measured at two successive harvests (1, initial and, 2, final). Relative growth rate is expressed in g g-1/day.
Standard errors (SE) were calculated for each variable.
Statistical analysis: All the experiments were conducted by using a Randomized Complete Block Design (RCBD) with three replications. For isolation of intrinsic growth potential of genotypes from their tolerance, all data from every experiment divided to its control value of every genotype. As data were corrected, data related to controls are equal and have no effect in total variance thus they deleted in statistical analysis. Then remaining data transformed into arcsin form and were subjected to analysis of variance (ANOVA) to examine the effects of cultivar, salinity treatment and their interactions after testing for normal distribution and homogenous variance. Significant differences between treatments were determined using Duncans multiple range test at the 0.05 level. Analysis of variance was performed by using the SPSS software version 9 and Microsoft Excel 2002.
For grouping the lines that show similar growth characteristic, different clustering methods as well as ordination based on principal components analysis was performed (Chatfield and Collins, 1995).
RESULTS
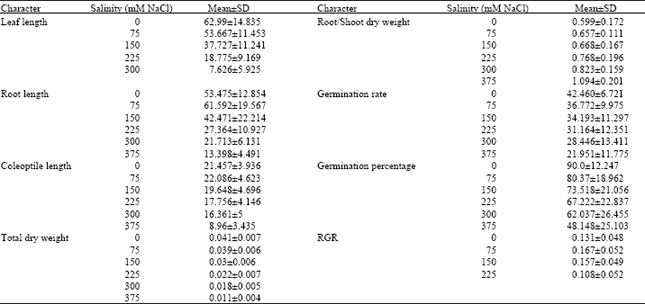
At early growth stage of seedling, the relative salt tolerance indicates for all the measured parameters varied among genotypes. The 9 genotypes used in this study were classified into 3 groups: salt-tolerant (Karchia, Tajan, Shiraz), moderately tolerant (Chamran, Shahryar, Tabasi) and salt-sensitive genotypes (Shahpasand, Pishtaz, Ghods) based on the ranking of 7 characters of these genotypes (Fig. 1). This study revealed a remarkable reduction in the germination rate, germination percentage, total dry weight and length of root, coleoptile and first leaf at early stage of seedling growth (Table 1). The low salinity treatment (75 mM) reduced these parameters to a lesser degree than high salinity treatment and even related to some parameters, such as root/shoot dry weight ratio, they increased (Table 2).
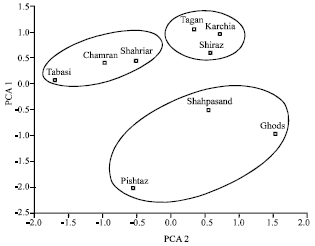 | |
| Fig. 1: | Cluster analysis and ordination of lines that treatment by different concentration of NaCl, PCA represented Principal Component Analysis |
| Table 1: | Means of some studied characters of 9 wheat genotypes at early stage of growth seedling |
 | |
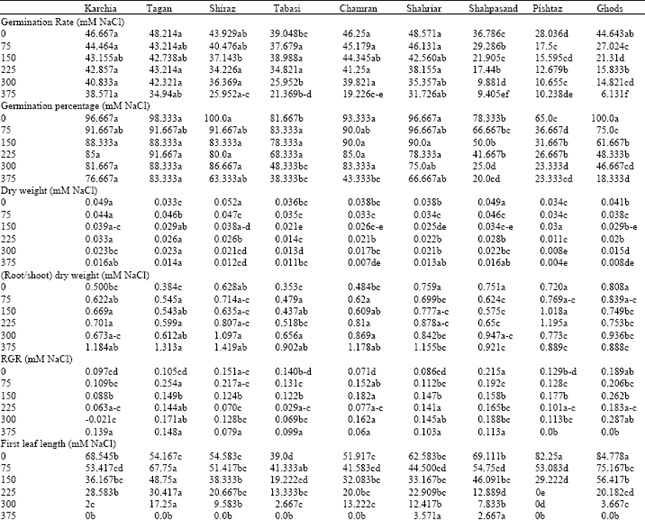
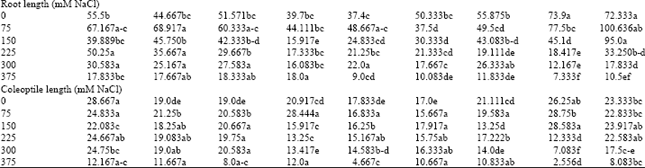
| Table 2: | Salt tolerance indices of physiological parameters in wheat genotypes under different salinity levels at germination and early growth stage |
 | |
 | |
| Value followed by different letter(s) differ significantly at p>0.05 | |
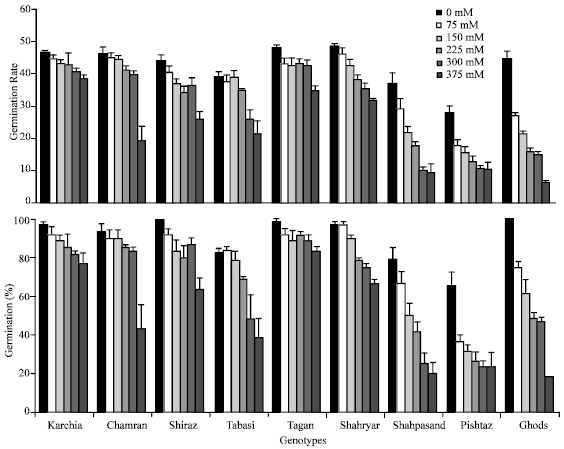 | |
| Fig. 2: | Effect of different salinity levels on the germination percentage (A) and germination rate (B) for different wheat genotypes. Error bars represent standard deviations |
Karchia, Tajan, Shiraz and chamran genotypes displayed the greatest germination percentage, dry weight and leaf area and leaf length in high salinity treatment. This indicates that salinity affected on the germination, emergence and early seedling growth and there was different response to salt stress among genotypes.
GENOTYPIC VARIATION IN GERMINATION
Although genotypes indicated a reduction in the final seed germination under increased salinity, analysis of variance revealed that significant differences were evident among the wheat genotypes for germination percentage and germination rate under different salinity treatments (p<0.01) germination percentage was high level among tolerant and then moderate genotypes (Fig. 2) and salinity treatments, with the exception of 375 mM NaCl, show no significant effect on it. Karchia and Tajan locates in grouping a, because they showed good germination response at all salinity levels among all lines. Also rate and percentage of germination of Karchia, Tajan, Chamran and Shiraz varied very low by increased salinity treatment until 300 mM NaCl.
Percentage of germination in the Pishtaz, Shahpasand, Ghods and Tabasi genotypes was strongly affected by all salinity treatments. Germination response to NaCl was significantly different in case of Pishtaz; its final germination was lesser than 70% at the control (0 mM NaCl) and by increased concentration of salt reduced to half at 150 mM. The reduction being strongest particularly at the highest level of salt concentration compared to control. Germination percentage of Tabasi, Pishtaz, Shahpasand and Ghods reached to 39, 36, 25 and 18%, respectively, as compared with the controls at 375 mM NaCl treatment. While final germination in Karchia, Tajan and Shiraz at 375 mM NaCl was 79, 84 and 63% of the control, respectively.
In view of germination rate, there was considerable reduction in this character in Pishtaz, Shahpasand and Ghods genotypes at all salinity levels compared to others, this parameter among 3 lines mentioned above restricted to 6-10% at 375 mM NaCl (Table 2). Although Ghods cultivar shows suitable germination at control level (percentage and rate germination was 100 and 44.6% respectively) but this line considered as sensitive cultivar because by increasing of salt concentration induced a significant decrease in these parameters. For example, percentage and rate of germination of Ghods genotype reduced to 18.3 and 13.7% at 375 mM NaCl than control.
GENOTYPIC VARIATION IN GROWTH
Dry matter production is an important criterion to evaluate salt tolerance in plants since it permits direct estimations of economic returns under specified saline conditions (Maas, 1986). Much less information is available about effect of salt stress on growth of leaf area and dry matter during early seedling growth, because young seedlings are rather delicate material for such measurements.
Effects of different salinity levels on dry weight of total plant and root/shoot dry weight ratios of different genotypes show in Fig. 3. Analysis of dry weight of the varieties showed that all the genotypes had decreased levels of dry matter production and concomitantly increased root/shoot ratio with increased substrate salinity (Fig. 3). The difference among the genotypes and salinity levels and their interaction was significant (p<0.01). Regarding to applied different salinity treatment in each genotype, there was no significantly different between 0-75 mM NaCl treatments however by increased salinity from 150 to 375 mM NaCl revealed that significant differences among the salinity levels (p<0.05). There were obvious differences among the 3 genotype groups in total dry matter, root/shoot biomass and RGR values. The salt tolerant group had higher dry matter at moderate and high salinity levels, reduced dry matter per plant for the salt tolerant genotypes were about 21% at 150 mM NaCl, 54% at 300 mM NaCl and 63% at 375 mM NaCl lesser than the controls, respectively, whereas in sensitive genotypes this value reduced about 58% at 150 mM NaCl, 68% at 300 mM NaCl and 81% at 375 mM NaCl compared with the controls, respectively.
Shiraz, Karchia and Shahpasand genotypes produced more dry matter than other genotypes under control condition but at high concentration of NaCl, ratio of root/shoot dry matter in Shahpasand was lower while Karchia and Tajan genotypes had significantly higher dry matter compared with other genotypes (Table 2). The effects of different salinity levels on the chlorophyll content revealed that total chlorophyll in the leaf of wheat seedlings was not significantly different from 0 to 150 mM NaCl in the tolerant and moderate groups but it was plunged with increased salinity levels from 150 to 300 mM NaCl. Among sensitive genotypes, chlorophyll content of Shahpasand was higher near to tolerant genotypes (data don’t show) therefore, it seems that higher dry matter and higher RGR in this variety may be caused for this reason. Response of biomass production in Pishtaz genotype to salinity was completely depended on salinity level and this genotype located in latest class among other lines at all salinity treatment.
Applied NaCl salinity significantly affected the RGR of all the genotypes, but there was no significantly difference among the salinity treatments at p<0.05 levels, interaction of the different genotype and salinity was significant at p<0.1 levels. The effect of salinity on RGR values varied according to the salt tolerance of the genotypes. RGR values was increased at low salt concentration (75 mM NaCl) in the tolerant and moderate groups, whereas the opposite was found in the salt sensitive group. In this group, RGR values not changed or reduced. For example, compared with the control, RGR value at 75 mM NaCl was increased by about 2.4-fold in Tajan, 2.1 fold in Chamran, 1.4 fold in Shiraz, 1.1 in Karchia and that decreased about 0.9 fold in Tabasi and Shahpasand lines. At higher salinity levels, RGR values were decreased in three genotypic groups, reduce of RGR was obvious especially in Shiraz and Tabasi at 225 mM NaCl level.
At early seedling growth, RGR value in sensitive genotypes of Ghods and Shahpasand was determined higher under non-saline condition, as compared with Karchia. In spite of this, Ghods genotype was reported as a salt sensitive line in Iran in previous studies (Kafi et al., 2003).
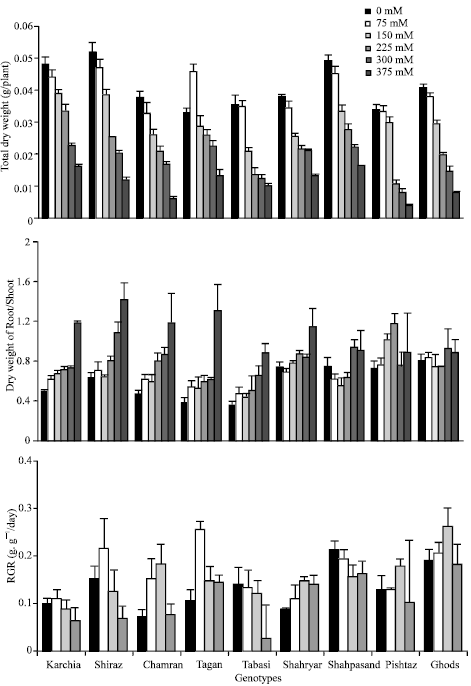 | |
| Fig. 3: | Effect of different salinity levels on the total dry weight (A), root/shoot dry weight (B) and RGR (C) for different wheat genotypes. Error bars represent standard deviation |
It seems that higher relative growth rate in both lines which mentioned above was depended on genotypic traits and caused harmful ions accumulated at toxic levels during future growth stage and so that decreased growth of these genotypes.
Concerning R/S ratio, there was a highly significant increase at Tajan, Karchia, Shiraz, Chamran and there was a non-significant change at sensitive genotypes such as Shahpasand, Shahryar and Ghods. The increase in this ratio results from relatively greater decrease in shoot than in root growth under salt stress (Fig. 4). The results in Table 2 and Fig. 4 show that there was remarkably increased in R/S ratio of tolerant and moderates groups at 375 mM NaCl treatment. The R/S ratio were increased by an average of 2.8 fold in tolerant genotypes, as compared with the control. Genotype Tajan indicated the highest (3.4 fold) while Ghods showed the lowest (1.1 fold) R/S ratio at the highest salinity treatment. These results are in agreement with the reports by Malik et al. (1979). The increase in root/shoot dry matter ratio often implies the development of a larger ratio of root length density to leaf area, which translate into a better capacity for sustaining plant water status under a given evapotranspirational demand (Malik et al., 1979).
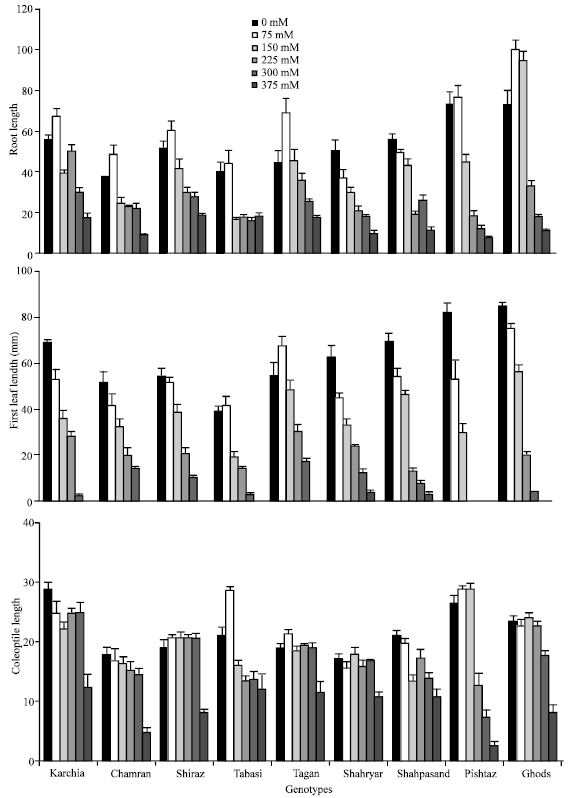 | |
| Fig. 4: | Effect of different salinity levels of the root length (A), first leaf length (B) and coleptiles length (C) for different wheat genotypes. Error bars represent standard deviations |
The classical explanation of water stress in plants growing in a saline environment is the reduced availability of soil water due to its osmotic potential. Osmotic adjustment and turgor maintenance in growing region was also important in sustaining root growth at low water potential (Morgan, 1995).
GENOTYPIC VARIATION IN PLANT HEIGHT
Plant height was measured in wheat cultivars and compared across salinity treatments. Height was found to decrease significantly (p<0.05) as the external salinity level increased. Root length of most genotypes at low salinity (75 mM NaCl) was increased, but at high salinity it was decreased by average 21% at 150 mM NaCl, 49% at 225 mM NaCl, 60% at 300 mM NaCl and 75% at 375 mM NaCl, as compared with the control (Table 1). The growth rate of root system in two sensitive genotypes (Ghods and Pishtaz) was approximately 1.46 fold of tolerant-line group under non-saline condition but it decreased remarkably at once plant pass through threshold concentration (150 mM NaCl). Therefore both genotypes located in the end of Duncan’s grouping at high concentration levels (Table 2). However as compared with sensitive salinity group, the tolerant-salinity group keep growth rate of root at higher level although they which had lower rate growth under non-saline condition. Root length of tolerant-group was 55 and 33% of the control at 300 and 375 mM NaCl, respectively, while it was 20.5 and 12% of the control at 300 and 375 mM NaCl within sensitive group genotypes.
Same to root length, elongation of the first leaf decreased significantly when salinity elevated (p<0.05). Development of leaf was strongly inhibited at 375 mM NaCl and leaves of most genotypes cannot grow. Compared with the control, first leaf length at 150, 225 and 300 mM NaCl was reduced by average 70, 45 and 17% in tolerant genotypes and by 66, 24 and 4% in sensitive genotypes, respectively. Statistically analysis showed that lowest leaf length was noted in genotype Pishtaz and the highest was in genotype Tajan at 300 mM NaCl, (Table 2). Salinity delays leaf emergence and reduced the leaf size of plants both longitudinally and laterally. Although, elongation of the first leaf decreased by enhanced salinity levels from 0 until 300 mM NaCl but the rate of reducing of total plant dry matter was slower (Fig. 3 and 4). This suggests that the relative effect of salinity stress on leaf elongation is significantly larger than that on photosynthesis. Leaf expansion is very sensitive to salt and water stress and it is completely inhibited under mild stress level that hardly affected photosynthetic rate (El-Hendawy et al., 2005; Taiz and Ziger, 2002). Mild water deficits also affect the development of the root system. Root-to-shoot biomass ratio appears to be governed by a functional balance between water uptake by the root and photosynthesis by the shoot. Simply state, a shoot will grow until it is so large that water uptake by the roots becomes limiting to further growth; conversely, root will grow until their demand for photosynthate from the shoot equals the supply. This functional balance is shifted if the water supply decreases (Taiz and Ziger, 2002).
Response of coleoptiles elongation to salinity treatment was not significantly different within varieties than control. There was non-considerable reduction in growth of coleoptiles among all genotypes at all salinity levels particularly at 0-300 mM NaCl. At the highest level of salt concentration, the reduction was lower in tolerant and moderate genotype groups but it being strongest in sensitive-genotype group compared to control (Fig. 4). Among these genotypes, the reduction of coleoptile’s length was more in Pishtaz. It decreased by about 47, 27 and 10% at 225, 300 and 375 mM NaCl, respectively, While it was 77, 86 and 42% in Karchia at 225, 300 and 375 mM NaCl, respectively, as compared with the control.
CONCLUSION
Little is know about seed germination responses of Iranian wheat cultivars to salinity stress. The studies were carried out to observe the influence of salinity on germination and seedling growth of germinating seeds of 9 wheat cultivars. The wheat genotypes in this study revealed significant difference for germinability and seedling survival under saline condition. Final germination of all genotypes decreased as the salinity level increased and salinity also delayed germination rate. It is assumed that germination rate and the final seed germination decrease with the decrease of the water movement into the seeds during imbibitions (Hadas, 1977). Increasing salinity concentrations in germination often cause osmotic and/or specific toxicity which may reduce or retard germination percentage (Waisel, 1972; Basalah, 1991). Similar declines in seed germination have been reported in the literature (Sharma et al., 2004; Gill et al., 2003; Garrciarrubio et al., 2003). Highly significant differences were observed among the sensitive and tolerant accessions for seed germination at high salinity treatments. These results are also similar to Jamil and Rha (2004). They reported that germination of sugar beet and cabbage decreased as the salinity concentration increased and salinity also delayed germination rate. Similar kind of results was reported by Jeannette et al. (2002). They found that the mean time to germination of almost all Phaseolus species increased with the addition of NaCl and this increase in median germination time was greater in higher concentration as compared to low concentration.
The result indicated that an increased salinity concentration caused delayed emergence of shoot compared to control. A continuous increase in length of shoot and root was detected in frequent days of germination in the control as well as salt treatments. The data on the average length of shoot and root shows that all wheat genotypes revealed a strong inhibition with the increasing level of salt solution. There was considerable reduction in the size of shoot and root at highest level of salinity (375 mM NaCl). These results showed sign of great inhibition of shoot and root growth with NaCl treatments. The decrease in length of shoot and first leaf length was more pronounced as compared to root in all NaCl salt treatments; however this decrease was more prominent in Pishtaz than others. Great inhibition in root length was also recorded in Pishtaz. The shoot and root length are the most important parameters for salt stress because roots are in direct contact with soil and absorb water from soil and shoot supply it to the rest of the plant. For this reason, root and shoot length provides an important clue to the response of plants to salt stress (Jamil and Rha, 2004). It was observed that the degree of the seedling growth reduction increased with the increasing concentration of salt. The reason for reduced shoot and root development may be due to toxic effects of the NaCl used as well as unbalanced nutrient uptake by the seedlings. High salinity may inhibit root and shoot elongation due to slowing down the water uptake by the plant (Werner and Finkelstein, 1995). Neumann (1995) indicated that salinity can rapidly inhibit root growth and hence capacity of water uptake and essential mineral nutrition from soil. Salt stress inhibited the growth of shoot more than root in Brassica species (Jamil et al., 2005). Similar observations have been reported in barley (Hordeum vulgare L.) (Huang and Redmann, 1995), pigeon pea (Cajanus cajan) (Subbarao et al., 1991), tepary bean (Phaseolus acutifolius) (Goertz and Coons, 1991) and tomato (Lycopersicon) (Foolad, 1996).
The results in this study indicate that the ranking among genotypes for salt tolerance based on the germination rate, dry weight of total plant, root/shoot dry weight ratio and first leaf length at the early growth stage show close correlated with their tolerance on salinity levels, whereas reduction of RGR, specially at low and moderate concentrations of NaCl, cannot revealed differences during early growth stage. This is probably due to short time period between two sampling times. Also, variation in storage content of the seed genotypes may be another reason for this. RGR can again be used as critical parameter by comparisons of their biomass production over a long growth period. Munns and James (2003) suggested that Salinity tolerance was defined as genotypic differences in biomass production in saline versus non-saline conditions over prolonged periods, of 3-4 weeks. In short-term experiments (1 week) measuring either biomass or leaf elongation rates revealed large decreases in growth rate due to the osmotic effect of the salt, but little genotypic differences, although there were genotypic differences in long-term experiments.
Among Iranian genotypes, Tajan indicated the highest salt tolerance while Pishtaz showed the lowest. Furthermore, Ghods and Shahpasand were more sensitive at moderate and high salinity levels and to become more tolerant at low salinity levels, it is suggested that maintaining the salinity at low levels is an important strategy for improving the growth of these two varieties. Also, among moderate genotypes Tabasi is more sensitive under saline condition at early growth stage.
Salt tolerance of plants varies during their successive growth stages (Bernstein and Hayward, 1958). The first stage, during which the crop is established, is regarded as particularly difficult, even for tolerant crops (Bernstein and Fireman, 1957; Maas and Hoffman, 1977). Although, studies of salt tolerance at early growth of seedling is important but it is not sufficient, the reproductive stage is the most important in terms of economic yield (Taiz and Zeiger, 2002; Wahid and Rasul, 2004). Therefore, assessment of the actual salt tolerance of the genotypes required to complete by studying of numerous physiological parameters at the reproductive stage for determining the salt tolerance of 9 genotypes that grown in soil under saline conditions within the same experiment.
ACKNOWLEDGMENTS
This study was supported in part by a grant-in-aid for research work from University of Azzahra, Tehran, Iran. We would also like to thank the management of the Seed and Plant Improvement Institute in Iran for providing seeds of wheat cultivars.
REFERENCES
- Baldwin, A.H., K.L. McKee and I.A. Mendelssohn, 1996. The influence of vegetation, salinity and inundation on seed banks of oligohaline coastal marshes. Am. J. Bot., 83: 470-479.
Direct Link - Murillo-Amador, B., E. Troyo-Dieguez, H.G. Jones, F. Ayala-Chairez, C.L. Tinoco-Ojanguren and A. Lopez-Cortes, 2000. Screening and classification of cowpea genotypes for salt tolerance during germination. Int. J. Exp. Bot., 67: 71-84.
Direct Link - Murillo-Amador, B., E. Troyo-Dieguez, A. Lopez-Cortes, C. Tinoco-Ojanguren, H.G. Jones and F. Ayala-Chairez, 2000. Path analysis of cowpea early seedling growth under saline conditions. Int. J. Exp. Bot., 67: 85-92.
Direct Link - Bernstein, N., W.K. Silk and A. Lauchli, 1993. Growth and development of sorghum leaves under condition of NaCl stress-spatial and temporal aspects of leaf growth-inhibition. Planta, 191: 433-439.
Direct Link - Cramer, G.R., G.J. Alberico and C. Schmidt, 1994. Leaf expansion limits dry matter accumulation of salt-stressed maize. Aust. J. Plant Physiol., 21: 663-674.
Direct Link - Delane, R., H. Greenway, R. Munnus and J. Gibbs, 1982. Ion concentration and carbohydrate status of the elongating leaf tissue of Hordeum vulgare growing at high external NaCl. 1. Relationship between solute concentration and growth J. Exp. Bot., 33: 557-573.
Direct Link - El-Hendawy, S., Y. Yakout, G.M. Hu, A.M. Awad, S.E. Hafiz and U. Schmidhalter, 2004. Evaluating salt tolerance of wheat genotypes using multiple parameters. Eur. J. Agron., 24: 445-452.
CrossRef - El-Hendawy, S.E., Y. Hu and U. Schmidhalter, 2005. Growth, ion content, gas exchange and water relations of wheat genotypes differing in salt tolerances. Aust. J. Agric. Res., 56: 123-134.
Direct Link - Flowers, T.J., A. Garcia, M. Koyama and A.R. Yeo, 1997. Breeding for salt tolerance in crop plants-the role of molecular biology. Acta Physiologiae Plantarum, 19: 427-433.
CrossRefDirect Link - Flowers, J. and M.A. Hajibagheri, 2001. Salinity tolerance in Hordeum vulgare: Ion concentrations in root cells of cultivars differing in salt tolerance. Plant Soil, 231: 1-9.
Direct Link - Gill, P.K., A.D. Sharma, P. Singh and S.S. Bhullar, 2003. Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regul., 40: 157-162.
CrossRefDirect Link - Goertz, S.H. and J.M. Coons, 1991. Tolerance of tepary and navy beans to NaCl during germination and emergence. HortScience, 26: 246-249.
Direct Link - Grieve, C.M. and D.L. Suarez, 1997. Purslane (Portulaca oleracea L.): A halophytic crop for drainage water reuse systems. Plant Soil, 192: 277-283.
CrossRefDirect Link - Hadas, A., 1977. Water uptake and germination of leguminous seeds in soils of changing matrix and osmotic water potential. J. Exp. Bot., 28: 977-985.
CrossRef - Hu, Y., W. Fricke and U. Schmidhalter, 2005. Salinity and the growth of non-halophytic grass leaves: The role of mineral nutrient distribution. Func. Plant Biol., 32: 973-985.
Direct Link - Ishikawa, S., T. Oikawa and A. Furukawa, 1991. Responses of photosynthesis, leaf conductance and growth to different salinities in 3 coastal dune plant. Ecol. Res., 6: 217-226.
Direct Link - Jamil, M. and E.S. Rha, 2004. The effect of salinity (NaCl) on the germination and seedling of suger beet (Beta vulgaris L.) and cabbage (Brassica oleracea capitata L.). Korean J. Plant Res., 7: 226-232.
Direct Link - Jamil, M., C.C. Lee, S.U. Rehman, D.B. Lee, M. Ashraf and E.S. Rha, 2005. Salinity (NaCl) tolerance of Brassica species at germination and early seedling growth. Electr. J. Environ. Agric. Food Chem., 4: 970-976.
Direct Link - Jaradat, A.A., M. Shahid and A. Al-Maskri, 2004. Genetic diversity in the Batini barley landrace from Oman: II. Response to salinity stress. Crop Sci., 44: 997-1007.
Direct Link - Kafi, M., W.S. Stewart and A.M. Borland, 2003. Carbohydrate and proline contents in leaves, roots and apices of salt-tolerant and salt-sensitive wheat cultivars. Russ. J. Plant Physiol., 50: 155-162.
CrossRefDirect Link - Katerji, N., J.W. Von Hoorn, A. Hamdy, F. Karam and M. Mastrorilli, 1994. Effect of salinity on emergence and on water stress and early seedling growth of sunflower and maize. Agric. Water Manage., 26: 81-91.
Direct Link - Malik, R.S., J.S. Dhankar and N.C. Turner, 1979. Influence of soil water irrigation deficits on root growth of cotton seedlings. Plant Soil, 53: 109-115.
Direct Link - Mansour, M.M.F., K.H.A. Salama, F.Z.M. Ali and A.F. Abou Hadid, 2005. Cell and plant responses to NaCl in Zea mays L. cultivars differing in salt tolerance. Gen. Applied Plant Physiol., 31: 29-41.
Direct Link - Mariko, S., N. Kachi, S. Ishikawa and A. Furukawa, 1992. Germination ecology of coastal plants in relation to salt environment. Ecol. Res., 7: 225-233.
CrossRefDirect Link - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Mass, E.V., S.M. Lesch, L.E. Francois and C.M. Grieve, 1994. Tiller development in salt-stressed wheat. Crop Sci., 34: 1594-1603.
Direct Link - Morgan, J.M., 1995. Growth and yield of wheat lines with differing osmoregulative capacity at high soil water deficit in seasons of varying evaporative demand. Field Crops Res., 40: 143-152.
Direct Link - Munns, R. and A. Termaat, 1986. Whole-plant responses to salinity. Aust. J. Plant Physiol., 13: 143-160.
Direct Link - Munns, R., D.P. Schachtman and A.G. Condon, 1995. The significance of a two-phase growth response to salinity in wheat and barley. Aust. J. Plant Physiol., 22: 561-569.
CrossRefDirect Link - Munns, R. and R.A. James, 2003. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil, 253: 201-218.
CrossRef - Munns, R., R.A. James and A. Lauchli, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot., 57: 1025-1043.
CrossRefPubMedDirect Link - Neves-Piestun, B.G. and N. Bernstein, 2005. Salinity-induced changes in the nutritional status of expanding cells may impact leaf growth inhibition in maize. Functional Plant Biol., 32: 141-152.
Direct Link - Pervaiz, Z., M. Afzal, Y. Xiaoe and L. Ancheng, 2002. Selection criteria for salt tolerance in wheat cultivars at seedling stage. Asian J. Plant Sci., 1: 85-87.
CrossRefDirect Link - Sharma, A.D., M. Thakur, M. Rana and K. Singh, 2004. Effect of plant growth hormones and abiotic stresses on germination, growth and phosphatase activities in Sorghum bicolor (L.) Moench seeds. Afr. J. Biotechnol., 3: 308-312.
Direct Link - Saqib, M., J. Akhtar and R.H. Qureshi, 2004. Pot study on wheat growth in saline and waterlogged compacted soil: II. Root growth and leaf ionic relations. Soil Tillage Res., 77: 179-187.
CrossRefDirect Link - Welbaum, G.E., T. Tissaoui and K.J. Bradford, 1990. Water relations of seed development and germination in muskmelon (Cucumis melo L.). 111. Sensitivity of germination to water potential and abscisic asid during development. Plant Physiol., 92: 1029-1037.
PubMedDirect Link - Werner, J.E. and R.R. Finkelstein, 1995. Arabidopsis mutants with reduced response to NaCl and osmotic stress. Physiol. Plant., 93: 659-666.
CrossRefDirect Link