
G. Rahimi
Department of Animal Science, Faculty of Agricultural Sciences, Mazandaran University, Sari, Iran,
A. Nejati-Javaremi
Department of Animal Science, Faculty of Agriculture, University of Tehran, Karaj, Iran,
K. Olek
Biopsytec GmbH, Berlin, Germany
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1389-1392







ABSTRACT
The objective of the present study was to estimate the allele and genotype frequencies of the BLAD, DUMPS and κ-CSN genes in Holstein young bulls of the national AI station of Iran. It is shown that the genetic mutation responsible for BLAD in Holstein cattle was absent in all young bulls entering to the national dairy cattle progeny test. Of the 37 young bulls genotyped for CD18/HaeIII gene mutation, 36 were normal homozygote (AA), 1 was heterozygote for the disease (AB) and no affected homozygote genotype (BB) were found. The frequency of the normal and BLAD mutant allele were 0.99 and 0.01, respectively. The analyzed results showed that the BLAD carrier prevalence in Iranian Holstein young bulls was approximately 2.8%. The genotype frequencies in κ-CSN/HinfI locus were 0.70 for homozygote (AA) and 0.30 for heterozygote (AB) animals while the frequency of the A allele was 0.85. In the present study it is shown that the genetic mutation responsible for DUMPS was absent in all of the young bulls analyzed from national AI station. It can be concluded that due to negative economical impact of the DUMPS and BLAD mutation on dairy cattle industry and their recessive mode of inheritance, attention has to be paid to any case of a young bulls entering to the testing program having in his origin any known carriers. Such a bull should be tested and if positive eliminated from the active population. Young bulls (under progeny testing program) should also be screened for both mutated alleles if in their progeny a high incidence of mortality is observed. This strategy would prevent the introduction of these mutated alleles in the Iranian endogenous cattle breeds.
PDF Abstract XML References Citation
How to cite this article
G. Rahimi, A. Nejati-Javaremi and K. Olek, 2006. Genotyping BLAD, DUMPS and κ-CSN Loci in Holstein Young Bulls of the National Animal Breeding Center of Iran. Pakistan Journal of Biological Sciences, 9: 1389-1392.
DOI: 10.3923/pjbs.2006.1389.1392
URL: https://scialert.net/abstract/?doi=pjbs.2006.1389.1392
DOI: 10.3923/pjbs.2006.1389.1392
URL: https://scialert.net/abstract/?doi=pjbs.2006.1389.1392
INTRODUCTION
The bovine leukocyte adhesion deficiency (BLAD) is an autosomal recessive hereditary disease characterized by greatly expression of the heterodimeric β-2 integrin adhesion molecules on leukocyte or white blood cells, resulting in multiple defects in leukocyte function. A defective protein on the surface of the white blood cells in affected calves prohibits the transport of these cells to the site of an infection (Nagahata et al., 1993). As a result, these calves have persistent infections and delayed wound healing. Most affected calves die within a few months after birth. While it is possible for some of the animals to live past two years of age, they are typically severely stunted in growth and suffer from recurrent various infectious conditions of the skin, gastrointestinal and respiratory tracts (Tajima et al., 1993). Two point mutations have been identified in the gene that encodes bovine CD18 in Holstein cattle affected with BLAD (Shuster et al., 1992). One mutation replaces adenine with guanine at nucleotide 383 and the other replaces cytosine with thymine at nucleotide 775. The latter is a silent mutation, causes no alteration in the amino acid sequence and has no phenotypic effect. The mutation at nucleotide 383 substitutes glycine for aspartic acid at amino acid 128. This mutation eliminates a TaqI restriction site and creates a HaeIII site, which allows the identification of normal, carrier and affected animals (Ribeiro et al., 2000). It is likely that the gene encoding defective CD18 on leukocyte is widespread in Holstein dairy cattle. The occurrence of BLAD in Holstein cattle has been reported in various countries, for example Denmark (Agerholm et al., 1993), England (Andrews et al., 1996), United States (Gilbert et al., 1993) and Japan (Nagahata et al., 1997). It is important to know the prevalence of the gene encoding impaired CD18 for BLAD for evaluation of a control program for the BLAD-associated gene in Holstein dairy herds. The prevalence of BLAD carriers and the gene frequency of BLAD in Holstein young bulls at national AI station of Iran are as yet unclarified.
The study of genetic polymorphisms of the caseins is of interest, since some variants could be more beneficial from the viewpoint of human nutrition (Boland et al., 2001) or be associated with milk quality, composition and technological characteristics (Yahyaoui et al., 2003). The kappa casein (κ-CSN) is the milk protein that determines the size and specific function of milk micelles and its cleavage by chymosin (rennin) is responsible for milk coagulation. It differs from other caseins in its solubility over a broad range of calcium ion concentrations. (Gutiérrez et al., 1996). Besides these characteristics, the mature κ-CSN protein has a labile peptide bound whose cleavage by chymosin produces a soluble hydrophilic glycopeptide (caseino-macropeptide) as well as an insoluble peptide or para-κ-CSN. The caseino-macropeptide is responsible for milk coagulation while the function of the para-κ-CSN is not well known (Yahyaoui et al., 2003). It has been reported that the bovine κ-CSN has six variants with A and B being the most common variants (Kaminski, 1996). The milk of cows carrying the B allele of κ-CSN contains a smaller and more homogeneous micelle size with an elevated concentration of κ-CSN in milk resulting in higher cheese yield (Ng-Kwai, 1998).
Deficiency of activity for uridine monophosphate synthase (DUMPS) of Holstein cattle is a component of the hypothesized multi-component complex. It is transmitted as an autosomal recessive trait. A carrier-normal mating results in one-half normal offspring and one-half carriers, regardless of sex. Embryos homozygous for DUMPS die early in gestation and do not survive to birth. The embryos often are resorbed during the first two-month of gestation, leading to more services per calving and longer than normal calving intervals (Shanks and Robinson, 1989). The only way to avoid economic losses is an early detection of DUMPS carriers. The purpose of the present study was to investigate the incidence of BLAD and DAMPS carriers and also κ-CSN genotyping in Holstein young bulls of Iran national AI station.
MATERIALS AND METHODS
DNA isolation: We genotyped 37 young bulls that have been born between 2003 and 2004 where registered for entering to the Iran national dairy progeny test programs. Ear tissue samples were collected from Holstein young bulls of National Animal Breeding Center of Iran using sample tagger developed by Biopsytec (Biopsytec GmbH, Kastanieallee, Berlin). Genomic DNA was extracted from ear tissue with the NucleoSpin Tissue Quik Pure Kit (Macherey and Nagel, Düren, Germany) according to the manufacture’s recommendations. All laboratory analyses have been done at Biopsytec’s laboratory in Germany.
| Table 1: | Primer sequences of the BLAD, DUMPS and κ-CSN loci in Holstein young bulls of Iran National AI station |
 | |
PCR conditions: The PCR reactions were carried out in a total volume of 20 μL, containing 10 mM dNTP-mix, 2.5 mM MgCl2, 2.5 U Thermo-start DNA polymerase and 100 ng genomic DNA. The PCR for BLAD was performed using the following conditions: an initial denaturation step of 5 min at 94°C, 35 cycles of 15 sec at 94°C, 20 sec at 69°C, 1.5 min at 72°C and a final extension of 10 min. at 72°C. The thermal cycling conditions for κ-CSN were: an initial denaturation step of 5 min at 94°C, 30 cycles of 15 sec at 94°C, 30 sec 60 sec at 69°C, 1.5 min at 72°C and a final extension of 5 min at 72°C. The PCR for DUMPS was performed using the thermal cycling conditions as: an initial denaturation step of 5 min at 94°C, 20 cycles of 1 min at 94°C, 1 min at 58°C, 1 min at 72°C and a final extension of 5 min at 72°C. The sequences of the primers for each locus are presented in Table 1.
PCR products and genotyping: The yield and specificity of PCR products were evaluated after electrophoresis in 1.5% agarose gel (Promega) with ethidium bromide staining. The PCR products were digested with specific restriction enzyme. For detection of CD18/HaeIII mutant allele a DNA fragment with 58 bp was amplified. The Aliquots of the amplified PCR products were digested with HaeIII restriction enzyme. The digested product was analyzed in a 3% agarose gel and stained with ethidium bromide. The allelic forms of the κ-CSN gene was identified based on the amplification of a 350 bp fragment of the resulted PCR products followed by digestion with the restriction enzyme HinfI. After digestion of the amplified products the DNA fragments were separated on 3% agarose gel. For DAMPS genotyping, the amplified of PCR products with 108 bp in length were digested by the AvaI enzyme to generate restriction fragments. The resulted fragments electrophoresed in 3.5% agarose gel. The gene frequencies in each locus were calculated by counting genotyping.
RESULTS AND DISCUSSION
We have genotyped 37 Holstein young bulls of Iran national AI station for occurrence of the CD18 and DUMPS mutations and also for detecting polymorphism in κ-CSN locus. The alleles and genotypes frequencies of the CD18/HaeIII locus are shown in Table 2. The primers were used to amplify CD18 locus produced a DNA fragment with the length of 58 bp.
| Table 2: | Distribution of genotypes (number of animals) and gene frequencies (%) of RFLP polymorphic at the BLAD, DUMPS and κ-CSN loci in Holstein young bulls of Iran national AI station |
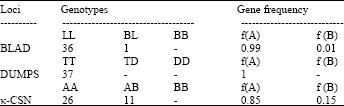 | |
In order to detect the occurrence of the CD18 mutation, the PCR products were digested with the HaeIII enzyme. This enzyme digests the PCR product into two fragments of 9 and 49 bp in normal homozygote individuals and three fragments of 9, 19 and 30 bp in affected animals. Heterozygote animals show four fragments of 9, 19, 30 and 49 bp. It is shown that the genetic mutation responsible for BLAD in Holstein cattle was absent in all the young bulls analyzed from the national dairy cattle progeny test. Analysis of the Holstein young bulls revealed that of the 37 bulls genotyped, 36 were normal homozygote (AA), 1 was heterozygote for the disease (AB) and no affected homozygote genotype (BB) were found. The frequency of the normal and BLAD mutant allele were 0.99 and 0.01, respectively. The analyzed results showed that the BLAD-carrier prevalence in Iranian Holstein young bulls was approximately 2.8%. Holstein herds have been genotyped for this mutation in several countries. The frequency of the BLAD mutant allele and the BLAD-carrier prevalence in Brazilian Holstein cows were 2.8 and 5.7%, respectively (Ribeiro et al., 2000). It has been shown that the BLAD frequencies are 3.5% in Argentinean Holstein cows (Poli et al., 1996), 13.4% in Danish Holstein-Friesian cattle (Jorgensen et al., 1993) and 8.1% in Japanese Holstein animals (Nagahata et al., 1997), 5.8 and 14.1% for American Holsteins bulls and cows (Shuster et al., 1992), respectively. The obtained results indicating that this mutation is prevalent among Holstein cattle throughout the world.
The different alleles of the κ-CSN gene were identified based on the amplification of 350 bp fragment located between nucleotide 201 of exon IV and nucleotide 149 of intron IV. The amplified fragment was digested in a reaction mixture containing restriction enzymes HinfI. The AA genotypes were characterized by the presence of three restriction fragments of 134, 132 and 84 bp and genotype BB with two fragments of 266 and 84 bp, respectively. The obtained allele and genotype frequencies of the CSN3/HinfI locus are shown in Table 2. The genotype frequencies were 0.70 for the AA genotype and 0.30 for the AB genotype while the frequency of the A allele was 0.85. No BB genotype for the CSN3/HinfI polymorphism was observed at the present study. The CSN3/HinfI polymorphism identified by Medrano and Cordova (1990) results in the substitutions of the amino acid alanine (allele A) with aspartic acid (allele B) at position 148 in the protein sequence. Thus, this polymorphism has the potential to directly or indirectly affect production traits. The present results showed high frequencies of allele A, similar to those reported in the literature. Kemenes et al. (1999) observed frequencies of 0.91 and 0.94 in Nelore and Gyr breeds, respectively. Regitano et al. (1999) found a frequency of 0.67 for the A allele in animals of the Canchim breed. In addition, other studies have shown higher frequencies of allele A compared to allele B in both taurine (Luciana et al., 2003) and Zebu breeds (Kemenes et al., 1999), regardless of whether dairy or beef cattle were studied. However, allele B has been found to predominate in the taurine dairy breeds Jersey and Brown Swiss (Van Eenennaam and Medrano, 1991). Literature data have demonstrated higher frequencies of the allele A In Zebu breeds compared to taurine breeds (Rogério et al., 2005).
DUMPS is a hereditary recessive disorder in Holstein cattle causing early embryo mortality during its implantation in the uterus. The only way to avoid the economic losses is early detection of DUMPS carriers. In order to detect the occurrence of the DUMPS mutation, the PCR products of the 108 bp were digested with the AvaI restriction enzyme. This enzyme digests the PCR product into three fragments of 53, 36 and 19 bp in normal homozygote individuals and two fragments of 89 and 19 bp in affected animals. Heterozygote animals show four fragments of 89, 53, 36 and 19 bp. At the present study we found that the genetic mutation responsible for DUMPS in Holstein cattle was absent in all the young bulls analyzed from the national dairy cattle progeny test. The frequency of affected allele in American Holstein dairy cattle has been estimated at 1-2% (Shanks and Robinson, 1990) but Kaminski et al. (2005) found no incidence of DUMPS carriers in Polish dairy cattle. Selection of bulls for artificial insemination is a very large contributor to decisions affecting genetic progress in current dairy cattle improvement. With increasing knowledge of the position and effects of major loci for quantitative variation, modifications of traditional selection procedures based only on phenotypes will be needed. Molecular data will help eliminate undesirable alleles and increase favorable alleles. Since BLAD and DUMPS is an economically important disease, Iranian dairy farmers should avoid the use of semen from carrier bulls. This strategy would prevent the death of Holstein calves from BLAD and DUMPS and also the introduction of these mutated alleles in the Iranian endogenous cattle breeds.
REFERENCES
- Andrews, A.H., J. Fishwick and R.J. Waters, 1996. Bovine leucocyte adhesion deficiency (blad) in a one year old Holstein Friesian bull-the first report in the United Kingdom Br. Vet. J., 152: 347-351.
PubMedDirect Link - Boland, M., A. MacGibbon and J. Hill, 2001. Designer milks for the new millennium Livest. Prod. Sci., 72: 99-109.
CrossRef - Gutierrez, A., E.A. Magen, H. Meade, C.F. Shoemaker, J.F. Medrano, G.B. Anderson and J.D. Murray, 1996. Alterations of the physical characteristics of milk from transgenic mice producing bovine κ-caseinAlterations of the physical characteristics of milk from transgenic mice producing bovine κ-casein J. Dairy Sci., 79: 791-799.
- Nagahata, H., T. Miura, K. Tagaki, M. Ohtake, H. Noda, T. Yasuda and K. Nioka, 1997. Prevalence and allele frequency estimation of Bovine Leukocyte Adhesion Deficiency (BLAD) in Holstein-Friesian cattle in Japan. J. Vet. Med. Sci., 59: 233-238.
PubMed - Kemenes, P.A., L.C.A. Regitano, A.J.M. Rosa, I.U. Paker and A.G. Razook et al., 1999. κ-casein, β-lactoglobulin and growth hormone allele frequencies and genetic distances in Nelore, Gyr, Guzera, Caracu, Charolais, Canchim and Santa Gertrudis cattle. Genet. Mol. Biol., 22: 539-541.
CrossRefDirect Link - Ribeiro, L.A., E.E. Baron, M.L. Martinez and L.L. Coutinho, 2000. PCR screening and allele frequency estimation of bovine leukocyte adhesion deficiency in Holstein and Gir cattle in Brazil. Genet. Mol. Biol., 23: 831-834.
CrossRefDirect Link - Medrano, J.F. and E. Aquilar-Cordova, 1990. Genotyping of bovine kappa-casein loci following DNA sequence amplification. Biotechnology, 8: 144-146.
PubMed - Nagahata, H., H. Nochi, K. Tamoto, H. Taniyama and H. Noda et al., 1993. Bovine leukocyte adhesion deficiency in Holtein cattle Can. J. Vet. Res., 57: 255-261.
Direct Link - Poli, M.A., R. Dewey, L. Semorile, M.E. Lozano, C.G. Albarino, V. Romanowski and O. Grau, 1996. PCR screening for carriers of Bovine Leukocyte Adhesion Deficiency (BLAD) and uridine monophosphate synthase (DUMPS) in Argentine Holstein cattle. Zentralbl Veterinaermed, 43: 163-168.
PubMed - Rogerio, A.C., N.O. Henrique, A.G. Marcos, C.S. Antonio and R.L. Catalina, 2005. Effects of CSN3 and LGB gene polymorphisms on production traits in beef cattle. Genet. Mol. Biol., 28: 262-266.
Direct Link - Shuster, D.E., M.E. Kehrli, M.R. Ackermann and R.O. Gilbert, 1992. Identification and prevalence of a genetic defect that causes leukocyte adhesion deficiency in Holstein cattle. Proc. Natl. Acad. Sci. USA., 89: 9225-9229.
PubMed - Kaminski, S., G. Grzybowski, B. Prusak and A. Rusc, 2005. No incidence of DUMPS carriers in polish dairy cattle. J. Applied Genet., 46: 395-397.
PubMedDirect Link - Van Eenennaam, A. and J.F. Medrano, 1991. Milk protein polymorphisms in California dairy cattle. J. Dairy Sci., 74: 1730-1742.
CrossRefDirect Link - Luciana, P.M.K.V., D.T. Talhari, A.P. Pereira, L.L. Coutinno and L.C.A. Regitano, 2003. Genetic characterization of Aberdeen Angus cattle using molecular markers. Genet. Mol. Biol., 26: 133-137.
Direct Link - Yahyaoui, M.H., A. Angiolillo, F. Pilla, A. Sanchez and J.M. Folch, 2003. Characterization and genotyping of the caprine kappa casein variants. J. Dairy Sci., 86: 2715-2720.
Direct Link