
Shahram Mohammady-D
Faculty of Agriculture, University of Shahrekord, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1354-1358







ABSTRACT
Twenty-one monosomic lines of variety Chinese Spring (apically fertile) were crossed with variety Y82187 (apically sterile) in order to identify chromosomes carrying genes for water-stress induced apical sterility. Segregation in a disomic F2 family obtained from a cross between CS and Y82187 was also evaluated under water-stress conditions. The results with the disomic F2 population suggested that 2 genes with complementary interaction control water-stress induced apical sterility. F2 monosomic analysis revealed that chromosome 3A of line Y82187 bred true for apical sterility. Monosomics for chromosomes 3D, 5D and 6D showed additional apically sterile plants in comparison with the disomic F2 population, whereas the data for other monosomic populations did not depart from the predicted 9:7 ratios. F2 progenies for some fertile and some sterile plants from F2 families monosomic for chromosomes 3D, 5D and 6D were evaluated under water-stress conditions in order to provide more information about these families. The results suggested that, in addition to chromosome 3A, chromosome 3D is the most likely chromosome to carry the second locus responsible for water-stress induced apical sterility in variety Y82187.
PDF Abstract XML References Citation
How to cite this article
Shahram Mohammady-D, 2006. The Chromosomal Locations of Genes Controlling Water-stress Induced Apical Sterility in Wheat (Triticum aestivum L.). Pakistan Journal of Biological Sciences, 9: 1354-1358.
DOI: 10.3923/pjbs.2006.1354.1358
URL: https://scialert.net/abstract/?doi=pjbs.2006.1354.1358
DOI: 10.3923/pjbs.2006.1354.1358
URL: https://scialert.net/abstract/?doi=pjbs.2006.1354.1358
INTRODUCTION
Reduced seed-set in wheat (Triticum aestivum) has become a common problem in many parts of the world particularly in regions with inadequate water reserves for irrigation (Joshi and Sthapti, 1995; Rawson, 1996). Environmental stresses, particularly drought and extreme high temperatures often result in reduced seed-set in some cultivars (Briggs et al., 1999; Rawson, 1996; Saini and Aspinal, 1981). Apical sterility or tip-sterility occurs in some wheat cultivars when the terminal spikelets fail to set seed following exposure to temperatures above 26°C or water-stress.
Dawson and Wardlaw (1989) reported that cultivar H757 from Afghanistan was susceptible to high temperatures (25-30°C) at the booting stage and set no seed in the apical half of the spike whereas the Australian cultivar Banks produced normal seed-set. Jamali (1991) reported the occurrence of sterile spikelets at the apex of spikes in the semi-dwarf varieties Faisalabad-85 and Y82187 from the Wheat Breeding Station at Faisalabad, Pakistan, when the temperature exceeded 26°C during the booting stages (Zadoks’ scale 40-49) (Zadoks et al., 1974) or when the plants encountered water-stress. However the seed-set was normal in the cultivar Kohinoor grown in the same environment.
Previous investigations confirmed the qualitative inheritance of apical sterility. Mohammady-D et al. (2003) reported that two recessive complementary genes control this character in Y82187 and that Faisalabad 85 possesses only one of these recessive genes. Shoran and Joshi (1987) in India reported a single recessive gene responsible for apical sterility in their varieties.
Additional information on the inheritance and chromosome location of the genes responsible for apical sterility will assist plant breeders to eliminate the recessive alleles for apical sterility whilst retaining beneficial genes on the same chromosomes.
Monosomic analysis is a suitable technique to determine chromosomal locations of major genes (McIntosh et al., 1996; Peng et al., 1998). F2 monosomic analysis is the most commonly used method to identify chromosomes carrying genes controlling qualitatively inherited characters in hexaploid wheat (Snape et al., 1983).
The chromosome carrying a dominant allele controlling a character in a wheat line can be identified by crossing that line with a series of 21 monosomic lines from another genotype which possesses the recessive allele. Monosomic plants (2n = 41) from each of the monosomic lines are emasculated and pollinated with the disomic variety (2n = 42). Monosomic F1 hybrids from each of the monosomic lines are self pollinated to produce the F2 generation in order to study the segregation of the contrasting phenotypes. As a result the F1 hybrid which contains the monosomic chromosome carrying the dominant allele will breed true in the F2 generation. The F1 hybrids from the other 20 monosomic lines will show normal Mendelian segregation in the F2 generation. A cross may be made between disomic plants (2n = 42) of the two cultivars and the F2 segregation is studied in the same environment as the F2 progenies from monosomic hybrids.
If two loci on different chromosomes determine a trait, 19 monosomic crosses should segregate in a similar ratio to the F2 disomic cross, whereas 2 monosomic crosses should generate segregation ratios that deviate from the F2 disomic ratio. The F2 disomic segregation ratio depends on the interaction between the two genes and ability to locate such genes using F2 monosomic analysis depends on the actual genotypes of the two parents.
The rationale for the current work was the fact that identifying the chromosomal location of the genes can pave the way for further studies of apical sterility using molecular markers specific to the candidate chromosomes. This would save times and money in comparison with a research across all chromosomes. In addition, when the chromosomal location of the genes is confirmed, the candidate chromosomes from apically fertile lines can be incorporated into the apically sterile ones using approaches such as inter-varietal substitution lines. The results of this study may lead to a better understanding of the inheritance of sterility in wheat and will help the researchers to include genes responsible for fertility so that the wheat varieties can remain fully fertile when subjected to water-stress. With these explanations, the objective of this study was to identify the possible chromosomal locations of the genes involved in water-stress induced apical sterility.
MATERIALS AND METHODS
Experiment 1: The current research was carried out at the Close House Field Station, University of Newcastle upon Tyne, UK during the period 1999-2003. The spikes of Chinese spring remain fully fertile under water-stress. The Pakistani line Y82187 shows apical sterility under water-stress conditions and set seeds only in the lower half of the spikes (Jamali, 1991; Kainth, 1994; Mohammady-D et al., 2003). Plants of 21 monosomic lines of Chinese Spring were crossed as female parents with the variety Y82187. Monosomic F1 plants were cytologically identified (except for chromosome 1B) and F2 seeds were produced for all populations. In addition, F2 seeds from a cross between euploid Chinese Spring and Y82187 was produced to provide a control F2 population. Monosomic plants were identified by examination of metaphase I in pollen mother cells prepared using the Feulgen staining method.
Germinated F2 seeds from 20 monosomic families and from the F2 disomic control were vernalised for 3 weeks in 7.5 cm pots in a cold cabinet at 2-7°C to promote early reproductive development. These seedlings were then transplanted into 10.5 cm pots in a green house where the minimum temperature was maintained 16°C and the maximum varied from 22 to 29°C. The pots were watered once daily to field capacity until the plants reached the booting stage (39-41 Zadok’s scale). Some nullisomic plants were observed in each F2 family. These plants were excluded from the experiment. The F2 families segregated for the time of booting stage development. Therefore the plants of F2 families were divided into 3 groups including early, intermediate and late. These groups were transferred successively to another bench in the green house for water-stress treatment, which involved withholding water for 6 days, followed by normal daily watering until the plants reached maturity.
The plants in each F2 family were categorised into apically fertile plants (with only one or no sterile spikelets at the apex of spikes) and apicall sterile plants with at least 2 sterile spikelets (terminal spikelets were not taken into account). The segregation ratio of apically sterile and fertile plants was calculated for each F2 monosomic family. The segregation ratio of the disomic F2 family was used as a control. The segregation ratios in monosomic F2 families were compared with the expected segregation ratio in the disomic F2 population using the Chi square (χ2) test.
Experiment 2: The results of experiment 1 revealed that, in addition to F2 monosomic family 3A, 3 other families also deviated from the expected ratio (Table 1). In experiment 2, the F2 fertile plants from each of these three lines were progeny tested and the F3 plants were subjected to a controlled water-stress treatment similar to Exp. 1. A few apical sterile F2 plants were also progeny tested to ensure that the water-stress imposed was adequate to induce apical sterility. In this experiment some extra F2 plants from the family monosomic for 3A were re-evaluated to provide more evidence for the true breeding observed in Exp. 1.
RESULTS
The number of F2 plants grown from each hybrid monosomic family was limited by the space available in the greenhouse. The aim was to identify the monosomic chromosomes which bred true for apical sterility and to grow additional F2 plants from segregating families to determine if the segregation ratio agreed with the F2 segregation observed in the disomic parental cross.
The monosomic population involving a chromosome carrying a factor involved in water-stress induced apical sterility was expected to have all plants in the sterile group. This was the case with chromosome 3A. segregation in monosomic populations, chromosomes 3D, 5D and 6D, did not fit to the proposed F2 ratio observed in the F2 disomic family. The monosomic F2 populations from the remaining chromosomes segregated according to the proposed F2 disomic ratio indicating that these chromosomes are not involved in tolerance to water-stress induced apical sterility. The numbers of fertile plants in the monosomic 3D, 5D and 6D populations were 6, 9 and 5 and the P values were 0.001, 0.041 and 0.002, respectively (Table 1).
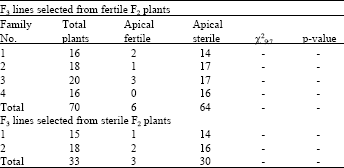
The results of F3 progeny tests for families 3D, 5D and 6D are shown in Table 2-4, respectively. Almost all F2 sterile plants selected from the above 3 families produced sterile F3 plants confirming the apical sterility in these F2 plants. Some F2 fertile plants selected from families 5D and 6D produced almost fertile plants when they were progeny tested at F3 generation. This indicates that these plants were homozygous for apical fertility. Some other F2 plants from the families 5D and 6D produced both fertile and sterile plants. The segregation ratios in the progeny of these plants fitted 9 fertile to 7 apically sterile using either individual or pooled data. Despite to families 5D and 6D, fertile F2 plants selected from family 3D produced almost sterile F3 plants (Table 2). This implies that the fertility observed in some plants of this family at the F2 generation is not conditioned by genes and they might remained fertile due to inadequate water-stress or due to water-stress escape.
DISCUSSION
The disomic F2 population obtained by crossing CS and Y82187 segregated in a ratio of 9:7 plants with fertile and apical sterile spikes. The 9:7 ratios indicate dihybrid segregation with complementary interaction between the two dominant genes. These results are in contrast with the results reported by Shoran and Joshi (1987) by which they emphasized that a single gene is responsible for the character. The literature is silent about this character and the author was not able to find any other studies regarding to apical sterility in wheat.
The results with F2 monosomic lines indicated that chromosome 3A of Y82187 carries a gene responsible for water-stress induced apical sterility and that one of the 3 chromosomes 6D, 5D and 3D is a possible carrier of the other gene. From these data it was impossible to identify which one of these 3 chromosomes is carrying a gene for apical sterility.
| Table 1: | Frequency of apical sterile and fertile plants in an F2 disomic and 20 F2 monosomic families and χ2 tests for the best goodness of fit |
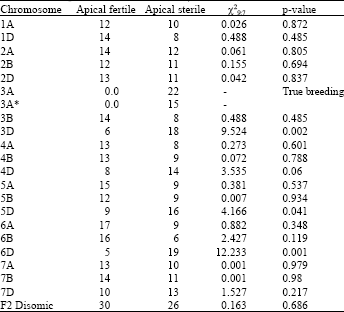 | |
| *: These plants were grown in Exp 2 to provide more evidence for true breeding | |
However, adjusting the χ2 for Yates’ correction revealed that deviation from 9:7 in F2 monosomic population for chromosome 5D is not actually significant (adjusted χ2 = 3.44 and p = 0.07). This F2 monosomic population was very late in reproductive development due to the absence of Vrn-D1 from CS, which resulted in the F2 plants being subjected to water stress at an earlier more susceptible stage of spike development. Thus this chromosome is not likely to be involved in controlling apical sterility.
If two genes are involved in the character, then it is theoretically expected that two F2 monosomic families breed true for apical sterility and the 19 remaining families indicate segregation in a 9:7 ratio. In the present study, more than two families indicated deviation from the expected ratio (Table 1). This deviation from the theory can be explained by two hypotheses. One is that the deviation observed for chromosomes 3D, 5D and 6D is due to the small number of plants and the two genes are located on chromosome 3A with a distance more than 50 cm or on the chromosome 1B which was not examined due to failure in seed production from F1 monosomic hybrids. The second is that the fertile plants from one of these three populations escaped the water-stress and are actually sterile. When plants are exposed to a biotic or an abiotic stress, it is likely that some plants escape from the stress. For instance, McIntosh et al. (1996) reported that in the stripe rust infection experiments there are often a proportion of plants that escape the infection. For this reason, they found many F2 monosomic populations that deviated significantly from a 3:1 expected ratio in addition to F2 monosomic 1B on which the gene for resistance was located.
| Table 2: | Number of apical sterile and apical fertile plants observed in F3 lines selected from either apical sterile or apical fertile plants of the F2 monosomic family CSM 3D x Y82187 |
 | |
| Table 3: | Number of apically sterile and apically fertile plants observed in F3 lines selected from either apical sterile or apical fertile plants of the F2 monosomic family CSM 5D x Y82187 |
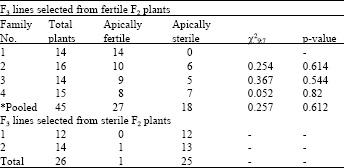 | |
| *: Segregating lines only | |
To test the latter hypothesis, the F2 fertile plants from lines 3D, 5D and 6D were progeny tested separately and the F3 plants were subjected to a controlled water-stress treatment. A few apical sterile F2 plants were progeny tested to ensure that the water-stress imposed was adequate to induce apical sterility. Both the fertile and apical sterile F2 plants from the cross with the line monosomic for 3D all bred true for apical sterility with a few plants possibly escaped the water-stress (Table 2). The fertile F2 plants from crosses with the lines monosomic for 5D and 6D either segregated in a ratio of 9 fertile: 7 apical sterile or bred true for fertile spikes. The F3 progeny from the F2 apical sterile plants tested from these 3 lines were all apical sterile. Segregation ratio in the F3 lines for chromosome 5D is presented in Table 3. As can be seen from the table, this segregation fitted the 9:7 ratio both for individual plants and for the pooled data. Similar results were observed for chromosome 6D (Table 4). These results provided evidence that chromosome 3A is the location of the first locus and 3D is possibly the location of the second locus. Crosses between ditelocentric lines of CS for chromosomes 3A and 3D and Y82187 could be used in the future to identify which chromosome arms carry the loci for apical sterility. This is the first time that the location of genes for apical sterility is reported and need to be studied further.
| Table 4: | Number of apically sterile and apically fertile plants observed in F3 families selected from either apical sterile or apical fertile plants of the F2 monosomic family CSM 6D x Y82187 |
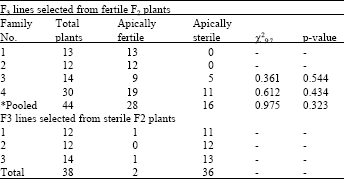 | |
| *: Segregating lines only | |
Since there are some limitations in using other cytogenetic stocks such as nullisomics for studying apical sterility, application of molecular markers specific to the candidate chromosomes (in particular 3A and 3D) is an alternative method to confirm the present results. The present results about location of genes for apical sterility is an important step towards methods to eradicate water-stress induced apical sterility from wheat varieties in dry regions such as Pakistan and Iran.
Similar to other genetic approaches, there are some advantages and limitations for the method used in the present study. F2 monosomic analysis is easily applied and has a desirable efficiency to establish the location of major genes (Peusha et al., 1996; Pirasteh and Welsh, 1975). Nonetheless, this method has not been satisfactory in locating minor genes and genes controlling quantitative characters where the effect of chromosome dosage can be as large as allelic differences under investigation (Law et al., 1987). To combat the above limitations, Giura and Saulescu (1996) used a modified method. They extracted 3 random F2 disomic plants to form F3 disomic families. They evaluated bulk seeds from the F3 progeny of each chromosome and compared the mean of each family with the overall mean of the experiment in order to identify chromosomal locations of the genes controlling grain size. This change in the method seems to produce more complications due to sampling errors arising from random selection of a small number of F2 plants from which trial materials were produced.
This study added to literatures that chromosomes 3A and 3D of variety CS may be promising for improving water-stress tolerance in wheat via conferring fertility to the apically sterile varieties and further studies including application of molecular markers specified for these chromosomes would provide more evidence about the role of these chromosomes in controlling water-stress induced apical sterility. Detecting genetic variations between varieties and landraces using cytogenetic analyses is still a proper way and the rapid increase in the application of molecular markers will not reduce the importance of cytogenetic approaches in plant breeding. Law et al. (1987) pointed out that “what is required is a fusion between the new marker techniques and both the established and developing methods of cytogenetics. With this regard, cytogeneticists have one prime asset in wheat which is not the case for other crops and that is the availability of a very large number of monosomic series and chromosome substitution lines”. With these explanations, using molecular methods in order to investigate various characters in cytogenetic stocks is a new area of research in order to determine the precise location of major genes previously located on the chromosomes using cytogenetic approaches and to find molecular markers associated with them.
Apical sterility induced by water-stress can be used as an indicator of the effect of water-stress on seed set.
REFERENCES
- McIntosh, R.A., J. Silk and T.T. The, 1996. Cytogenetic studies in wheat XVII. Monosomic analysis and linkage relationships of gene Yr15 for resistance to strip rust. Euphyt, 89: 395-399.
Direct Link - Mohammady-D, S., K. Moore and J. Ollerenshaw, 2003. Qualitative inheritance of water-stress induced apical sterility in wheat (Triticum aestivum). Hereditas, 138: 237-240.
CrossRefDirect Link - Peusha, H., S.L.K. Hsam and F.J. Zeller, 1996. Chromosomal location of powdery mildew resistance genes in common wheat (Triticum aestivum L. em. Thell.) 3. Gene Pm22 in cultivar Virest. Euphytica, 91: 149-152.
Direct Link - Saini, H.S. and D. Aspinall, 1981. Effect of water deficit on sporogenesis in wheat (Triticum aestivum). Ann. Bot., 48: 623-633.
Direct Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link