
Mahmoud Abu-Samak
Department of Medical Technology, Faculty of Allied Medical Sciences,
Applied Science University, Amman, Jordan
Fahmi Mahmoud
Faculty of Pharmacy, Al-Zaytoonah University of Jordan, Amman Jordan
Moayad Khataibeh
Department of Medical Technology, Faculty of Allied Medical Sciences,
Applied Science University, Amman, Jordan
Suhail Hamdan
Medical School, Al-Quds University, Al-Quds, Palestine
Aurelia Crevoi
Department of Human and Animal Physiology, Moldova State University, Moldova
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1350-1353







ABSTRACT
This study was designed to investigate the effects of Melanocyte stimulating hormone MSH on serum testosterone T and estradiol E2 hormones concentrations in alloxan-induced diabetic rats. Eighty male and female Sprague Dawley rats, weighing 180-200 g, were divided into four groups of normal rats and four groups of alloxan-induced diabetic rats were given intraperitonially (i.p.) a daily injection of 20 mg alloxan solution/100 g of body weight for 10 days. Two groups, male and female from the normal and 2 diabetic groups served as controls and did not inject with MSH 2 groups, male and female from the normal rats and 2 groups from the diabetic rats injected (i.p) daily with MSH at a dose of 2-microg/100 g of body weight, for 10 days. The control group was only injected with the same volume of normal saline. Serum glucose concentrations were higher and serum insulin, testosterone and estradiol concentrations were lower in diabetic rats than those in the control groups. MSH administration decreased the elevated blood glucose concentrations of the diabetic rats to the normal levels and decreased estradiol concentration in female normal rats while increased the testosterone concentration in male normal rats. Present findings indicate that MSH plays adaptive role during early stages of alloxan induced-diabetes mellitus. Further studies are needed to identify the mechanism.
PDF Abstract XML References Citation
How to cite this article
Mahmoud Abu-Samak, Fahmi Mahmoud, Moayad Khataibeh, Suhail Hamdan and Aurelia Crevoi, 2006. Effects of Melanocyte-stimulating Hormone on Plasma Levels of Testosterone and Estradiol Hormones in Alloxan-Induced Diabetic Rats. Pakistan Journal of Biological Sciences, 9: 1350-1353.
DOI: 10.3923/pjbs.2006.1350.1353
URL: https://scialert.net/abstract/?doi=pjbs.2006.1350.1353
DOI: 10.3923/pjbs.2006.1350.1353
URL: https://scialert.net/abstract/?doi=pjbs.2006.1350.1353
INTRODUCTION
Melanocyte Stimulating Hormone (MSH) has modulatory role in food intake, body fat and glucose metabolism (Biebermann et al., 2006; Fehm et al., 2001; Costa et al., 2006), but little is known about the effects of MSH on gonadal function during diabetes mellitus and practically there are no literature relating to the MSH influence on sex hormones levels during diabetes mellitus. A number of investigators reported on the relationship between gonadal dysfunction and Diabetes Mellitus (DM) in humans and animals (Matsushita et al., 2005, Komaki et al., 2005). Male and female reproductive alterations have been widely reported in individuals with diabetes including decreases in testicular testosterone production, reduced serum LH and male and female estradiol (Ballester et al., 2004; Steger and Rabb, 1997; Sanguinetti et al., 2004). Although several investigators have reported serum E2, free Testosterone and LH not affected in diabetic rats (Komaki et al., 2005) and diabetes-related effects on testicular function have been attributed to the lack of insulin. The regulatory action of this hormone is known and observations of a direct effect on both leydig cells (Khan et al., 1992; Hurtado de Catalfo et al., 1998) and Sertoli cells (Borland et al., 1984; Mita et al., 1985) have been reported. Nonetheless, the data are confusing and the exact role that insulin plays in the regulation of the male reproductive function is still unclear, even though the melanocortins are known to mediate some of the effects of both leptin and insulin (Benoit et al., 2004; O'Shaughnessy et al., 2003; Costa et al., 2006).
Therefore the aim of the present research is to investigate whether MSH can stimulate the gonadal axis by altering glucose homeostasis or sex hormones dynamic in diabetic rats.
MATERIALS AND METHODS
Animals: This research was carried out at the laboratories of Department of Medical Technology, Applied Science University from September 2005 to January 2006. Eighty male and female Sprague Dawley rats, were housed in our laboratory for at least one week before the experiments. The rats were maintained in a temperature-controlled room (22-25°C). Food and water were available for the animals all the time and without any restrictions. The weights of rats were taken on the day of the experiment and only those weighing 180-250 g were used in this study.
The animals were divided into two sections according to the sex of the animal and the animals of each section were divided into four treatment groups: control, MSH injected rats (MSH), Alloxan-induced diabetic rats (Allox) and Alloxan-diabetic rats injected with MSH (Allox+MSH). All the animals were given daily injections of following preparations, for 10 days: Control rats injected with 1 mL of normal saline, (MSH) and (Allox+MSH) Rats injected (i.p. daily with-alpha melanocyte stimulating hormone (Sigma Firm, USA) at a dose of 2 microgm (dissolved in 1 mL of normal saline)/100 g of body weight. (Allox) and (Allox+MSH) rats were given intraperitonially (i.p.) a daily injection of 20 mg alloxan (dissolved in 1mL of normal saline) solution/100 g of body weight (Sigma Firm, USA).
At the end of the of the experiment, all rats were fasted for 12 h before they were sacrificed and blood collected.
Serum analysis: The serum was isolated by centrifugation and analyzed by radioimmunoassay for serum insulin, estradiol and testosterone using a rat insulin, estradiol and testosterone radioimmunoassay kit (Cea-Jre-Sorin Firm, France). Serum glucose concentrations were measured by the glucose oxidase method using a spectrophotometer (Cecil ce 1010 England).
Statistical analysis: Data were expressed as means±SE and were analyzed with a two-way ANOVA followed by LSD multiple comparison test, using Statistica Software (OK, USA). Differences were considered significant at p<0.05.
RESULTS
AND DISCUSSION
Daily MSH administration for 10 days did not alter insulin levels but normalized blood glucose in the Allox rats, compared with control group (Table 1 ). Diabetic male rats showed a low weight gain and decreased levels of plasma insulin and testosterone when compared with healthy controls (Table 1). Diabetes also induced hyperglycemia (Table 1 and 2).
Allox-induced diabetes increased estradiol concentration, while decreased testosterone concentration compared with control groups (Fig. 1 and 2). MSH administration decreased estradiol concentration and increased testosterone concentration in diabetic groups (Fig. 1 and 2).
Present results indicate that MSH plays modulatory role on sex hormones levels in diabetic rats. MSH may exert this role through glucose homeostasis peripheral pathway because there was no change in insulin concentration while it normalized blood sugar in both male and female diabetic rats. Costa et al. (2006) reported that MSH has reciprocal effects in which MSH appears to increase sensitivity to insulin when present in the CNS, while MSH in the periphery is necessary for insulin resistance. Their results are in agreement with our results in that MSH plays modulatory role in diabetes mellitus type 2. Hochgeschwender et al. (2003) stated that the regulation of glucose homeostasis requires the integration of both central and peripheral melanocortin signalling systems. Furthermore, Abu-Samak (2000) and Savontaus et al. (2004) reported that MSH could serve as a potential strategy for anti-obesity and/or antidiabetic therapy.
Also the clear differences in sex hormones levels observed between male and female rats suggest that MSH has contrasting effects on male and females rats. This is supported by the reports of Kim et al. (1999), Robertson et al., 2003. There are researches indicating that estrogen normally acts in the brain to reduce food intake and body weight in female individuals (Clegg et al., 2003) and other reports show that MSH mimics these effects (Kim et al., 1999; Robertson et al., 2003 ; Fan et al., 1997). On the other hand, numerous experiments have reported that MSH has been shown to stimulate gonadotropin secretion in humans (Celis, 1985), in which an elevation of plasma testosterone hormone levels in male group injected with MSH have regulatory effects on the physiological function of rats. This is supported by previous lines of evidence that indicate that elevation of testosterone support the concept of a functional continuity between the central and peripheral actions of melanocortins in regulating certain physiological functions like sexual behavior (Van der Kraan et al., 1998). Further studies are needed to identify the mechanism underlying this enhanced sensitivity to peripherally administered MSH. There are diabetes.
| Table 1: | Effects of melanocyte stimulating hormone and diabetes on physical and seric parameters* |
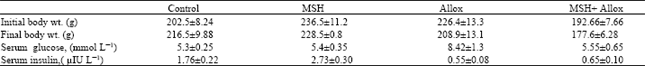 | |
| Table 2: | Significant differences (p-values) between female estradiol (a) and male testosterone (b) concentrations |
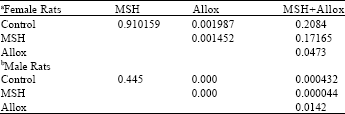 | |
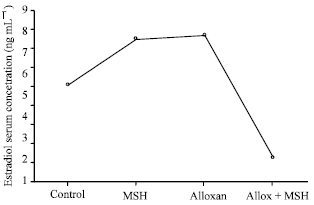 | |
| Fig. 1: | Effects of peripheral injection of MSH for 10 days on estradiol serum concentration. Values are means±SE |
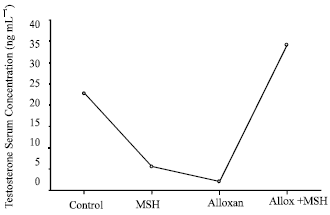 | |
| Fig. 2: | Effects of peripheral injection of a-MSH (2 microg/100 g bw) for 10 days on Testosterone serum concentration in the male rates. Values means±SE |
Another future research is in the area of anti-obesity and/or antidiabetic therapy in cases of several possible explanations that can be addressed in future work such as sex differences in the intracellular signalling cascades of MSH and insulin and if MSH-based therapeutics is specifically active in the CNS or peripheral circulation for the treatment of type 2 gonadal dysfunction.
REFERENCES
- Ballester, J., M.C. Munoz, J. Dominguez, T. Rigau, J.J. Guinovart and J.E.R. Gil, 2004. Insulin-dependent diabetes affects testicular function by FSH- and LH-linked mechanisms. J. Androl., 25: 706-719.
Direct Link - Benoit, S.C., D.J. Clegg, R.J. Seeley and S.C. Woods, 2004. Insulin and leptin as adiposity signals. Recent Prog. Horm. Res., 59: 267-285.
PubMed - Biebermann, H., T.R. Castaneda, F. van Landeghem, A. von. Deimling and F. Escher et al., 2006. A role for beta-melanocyte-stimulating hormone in human body-weight regulation. Cell Metab., 3: 141-146.
PubMedDirect Link - Costa, J.L., U. Hochgeschwender and M. Brennan, 2006. The role of melanocyte-stimulating hormone in insulin resistance and type 2 diabetes mellitus. Treat Endocrinol., 5: 7-13.
PubMed - Hochgeschwender, U., J.L. Costa, P. Reed, S. Bui and M.B. Brennan, 2003. Altered glucose homeostasis in proopiomelanocortin-null mouse mutants lacking central and peripheral melanocortin. Endocrinology, 144: 5194-5202.
PubMed - Fehm, H.L., R. Smolnik, W. Kern, G.P. McGregor, U. Bickel and J. Born, 2001. The melanocortin melanocyte-stimulating hormone/adrenocorticotropin 4-10 decreases body fat in humans. J. Clin. Endocrinol. Metab., 86: 1144-1148.
CrossRefPubMedDirect Link - Hurtado-de-Catalfo, G., I. Nelva and T. de Gomez-Dumm, 1998. Lipid dismetabolism in Leydig and Sertoli cells isolated from streptozotocin-diabetic rats. Int. J. Biochem. Cell Biol., 30: 1001-1010.
CrossRefDirect Link - Komaki, K., Y. Ohno and N. Aoki, 2005. Gonadal hormones and gonadal function in type 2 diabetes model OLETF (Otsuka Long Evans Tokushima Fatty) rats. Endocrine J., 52: 345-351.
Direct Link - Matsushita, M., K. Tamura, S. Osada and H. Kogo, 2005. Effect of troglitazone on the excess testosterone and LH secretion in thyroidectomized, insulin-resistant, type 2 diabetic Goto-Kakizaki rats. Endocrine, 27: 301-305.
CrossRef - O'Shaughnessy, P.J., L.M. Fleming, G. Jackson, U. Hochgeschwender, P. Reed and P.J. Baker, 2003. Adrenocorticotropic hormone directly stimulates testosterone production by the fetal and neonatal mouse testis. Endocrinology, 144: 3279-3284.
CrossRef - Robertson, R.P., J. Harmon, P.O. Tran, Y. Tanaka and H. Takahashi, 2003. Glucose toxicity in β-cells: Type 2 diabetes, good radicals gone bad and the glutathione connection. Diabetes, 52: 581-587.
CrossRef - Sanguinetti, R.E., K. Ogawa, M. Kurohmaru and I. Hayashi, 2004. Ultrastructural changes in mouse Leydig cells after streptozotocin administration. Exp. Anim., 44: 71-73.
Direct Link - Savontaus, E,. T.L. Breen, A. Kim, L.M. Yang, S.C. Jr. Chua and S.L. Wardlaw, 2004. Metabolic effects of transgenic melanocyte-stimulating hormone overexpression in lean and obese mice. Endocrinology, 145: 3881-3891.
CrossRef - Steger, R.W. and M.B. Rabe, 1997. The effect of diabetes mellitus on endocrine and reproductive function. Proc. Soc. Exp. Biol. Med., 214: 1-11.
PubMed