
Morteza Shahbazi-Moghadam
School of Physics, University College of Science, Tehran, Iran
Nazly Mahdavi
Institute of Biophysics and Biochemistry of University of Tehran, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1287-1292







ABSTRACT
Electric field (4400 V/m), magnetic field (B = 12 μT ), at zero frequency and electromagnetic magnetic field at 30 kHz and 92 μT, produced by video display terminals have been produced to observe how they affect ion transport processes and ATP (Adenosine Triphosphatase) splitting through changes in the activation of the membrane Na, K- ATPase in mousse brain. Both processes varied remarkably with magnetic and electromagnetic fields, while the variation with DC electric field was negligible. ATP splitting by Na, K-ATPase suspensions decreases for enzyme under normal conditions, with maximum effect at one hour of use of video display terminals. ATP splitting increases when the enzyme activity is lowered to less than half its optimal value by changes in temperature or ouabain concentration. These are because of the effect of the ionic currents on ion binding at the enzyme activation sites. Same mechanism could account for the effects of electromagnetic fields on cells, as the transmembrane enzyme can convey the effect of an extra cellular signal into the cell via ionic fluxes and the measured threshold field is within the range of reported biological effects. An oscillator similar to that of video display has been designed and constructed. The subjects were 60 mousses which were irradiated in a cage specially designed for our research purposes in different time of exposures. The results showed a good correlation between the time of exposure and the field intensities (R = 0.68, n = 60...., p<0.05). Present findings show that the specific activity decreases and is the minimum for 60 min of video display terminals usage and increases thereafter.
PDF Abstract XML References Citation
How to cite this article
Morteza Shahbazi-Moghadam and Nazly Mahdavi, 2006. Effect of Video Display Terminal Radiations on Ion-transporting Adenosine Triphosphatase in Brain. Pakistan Journal of Biological Sciences, 9: 1287-1292.
DOI: 10.3923/pjbs.2006.1287.1292
URL: https://scialert.net/abstract/?doi=pjbs.2006.1287.1292
DOI: 10.3923/pjbs.2006.1287.1292
URL: https://scialert.net/abstract/?doi=pjbs.2006.1287.1292
INTRODUCTION
A molecule which is immobilized oriented or tumbling more slowly than the frequency of a periodic field may interact with the field to produce chemical effects that are uncommon in a homogeneous solution. Among these effects are the alteration of the rate of a chemical reaction and the exchange of energy between the oscillating field and the conformation of the molecule (Tsong, 1992; Aizman and Aperia, 2003; Xie, 2003; Donnet et al., 2003). When certain conditions are satisfied, this exchange allows the molecule to absorb and couple the energy of the field to drive an undergone reaction (Bruce, 1999; Buyukusal and Mete, 1999). The efficiency of energy coupling depends on field strength and frequency and the ligand concentration (Dobretsou et al., 1999).
There are windows of these parameters to achieve efficient coupling. These windows can be expressed in terms of the rate constants and equilibrium constants of the catalytic reactions (Duran et al. 2003) and the amplitude and frequency of the periodic field (Blank and Soo, 1992; Fedosova et al., 2003). This mechanism allows cells to receive process and transmit energy of high and medium level periodic potentials by means of membrane enzymes or receptors (Tsong, 1992).
Many experiments on the effects of alternating currents on ATP-splitting by the membrane Na, K-ATPase suggest that the mechanism involves changes in the activation of the ion-binding sites by different fields (Sweadner and Feschenko, 2001; Faller et al., 2003). The model based on changes in ion activation differs from the electro-conformational coupling involving Na, K-ATPase (Blank, 1992). The electro-conformational coupling model suggests that there is a direct action of the electric field on the enzyme in the membrane (e.g., field-dipole interaction), leading to a change in its conformation, while the changes in ion activation model can be effective with interaction at the external binding site only (Sweadner and Feschenko, 2001; Fedosova et al., 2003) .
The similar effect of low-frequency electromagnetic fields 30-1000 Hz in human and animal was also studied and reported (Abdollahzadeh et al., 1996; Blank and Soo, 1992; Morgan and Nair, 1992). Because electromagnetic fields are also generated by video display terminals, it is plausible to investigate the role of electromagnetic field generated by these displays as a possible factor causing adverse health outcomes. Both the magnetic flux density at a fixed distance from video display and number of hours of video display use has been associated with adverse health outcomes (Paul, 1992). However, the relevance of such indices or surrogates to the actual average magnetic field exposure level, also known as time-weighted average magnetic field exposure, in the way the present work was focused, has not been demonstrated.
The aim of present research was to study the effect of different fields and especially the electromagnetic field on the brain through the in vitro study of the brain of mousses. So far few researches have been dedicated to the study of the effects of these fields on the brain. Recently Roots et al. (2003) and Bulygina (2003) have undertaken researches on the Na-pump kinetic properties of the brain and Glutamate receptors regulating Na, K-ATPase in cerebellum respectively. The main problem seemed to be the simulation of video display terminals out-put fields. This was feasible for us as we were able to design any kind of electronic and electric devices. To accomplish this aim, different field sources have been designed and constructed in order to simulate the fields in question. The results revealed that there was no remarkable effect of DC electric field on Na, K-ATPase splitting, while DC magnetic and electromagnetic fields similar to video display terminals had remarkable effects and somehow about the same effect.
MATERIALS AND METHODS
This study has been undertaken during 2004-2005 in the Department of Physics and the Department of Biophysics at Tehran University. The experiments were carried out on mousse brain. The mousses to be studied were placed into the plastic experimental chamber of about three liters in volume allowing the mousses be irradiated by different fields in different duration times; fifteen to ninety minutes (Fig. 1). The distance between the fields’ electrodes (parallel plates in electric and Helmholtz windings in magnetic and electromagnetic fields were 35 cm (Fig. 2).The electric field intensity varied by means of a DC supply varying from 200 to 880 volts. DC magnetic field was generated by DC current applied to Helmholtz windings and the field intensity was set to 3.125 mA. In order to produce the same electromagnetic field as in video display terminals, a saw-tooth oscillator with the same frequency as the horizontal frequency of video display terminals has been designed and constructed (Fig. 3).
 | |
| Fig. 1: | The plastic chamber of about three liters of volume allowing mousse irradiated. Holes are made for proper aeration |
 | |
| Fig. 2: | The aluminum circular plates for providing electric field and also Helholtz windings which are between circular aluminum plates with related suppliers |
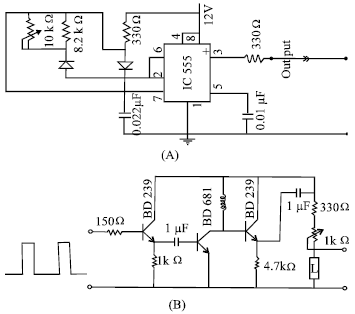 | |
| Fig. 3: | Circuit of Saw-tooth oscillator simulating the same wave and frequency as the horizontal frequency of video display terminals. (A) Square pulse generator and (B) square to saw-tooth converter |
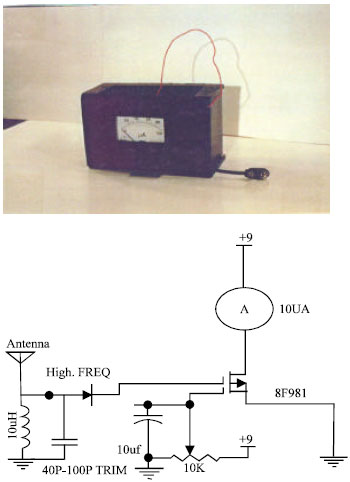 | |
| Fig. 4: | Circuit for flux-meter (photo) allowing the comparison of the both video display terminal and designed circuit intensities and necessary adjustment |
In order to compare the intensities, a special flux meter has also been designed and constructed (Fig. 4). The saw-tooth waves with appropriate current were then applied to Helmholtz windings to produce the same electromagnetic field. The estimated power density of the electromagnetic field in the vicinity of the subjects was about; 100 μ w cm-1.
The enzyme was prepared from ox-brain according to Skou, 1957, the specific (Na++K+) activity was 25-300 μmol Pi mg-1 protein-1 h-1 (4 mM Mg2+, 3 mM ATP, 130 mM Na+, 20 mM K+, pH 7.4, 37°C). The Mg2+ activity (equal to the ouabain-insensitivity) was less than 0.5% of the total activity. The activity was tested by measuring the amount of 32P released from ATP labeled with 32P in the γ position. The test volume was 1 mL, which besides 30 mM buffer, histidine-HCl or Tris-HCl adjusted to require pH at 37 or 0°C contained [32P] ATP, Na+, K+ and Mg2+ in the concentrations given on the figures. The reaction time was in all the experiments adjusted in such way that the maximum hydrolysis of the added ATP was less than 20%.
Under these conditions, the hydrolysis was a linear function of time. The reaction was stopped by the addition of 0.1 mL 50% trichloroacetic acid; blank was with 10-4 M ouabain in the test medium. The hydrolysis of [32P] ATP was determined according to the method of Lindberg and Ernester (Skou, 1957). The 32P was counted in a flow-type proportional counter (Shahbazi and Faghahati, 2006). [32P] ATP, Tris and ATP were from Sigma.
RESULTS AND DISCUSSION
This work revealed that the electric and magnetic fields at extremely low frequency on biological cells are confronted by two problems; a) low- frequency signals do not penetrate the cell membrane to any appreciable extent (induced currents pass around cells rather than through them) and b) the low intensities at which fields are unlikely to influence cellular reactions. In fact Na, K-ATPase, the membrane enzyme responsible for establishing ion gradients across cell membranes, catalyzes the coupled transport of three sodium ions from inside out and two potassium ions from outside in for each ATP split (Keep, 1999).
The enzyme is activated when potassium interacts with the outside surface and sodium with the inside, (the catalytic site is on the inside surface). As ionic currents can affect the ion binding and thus the activation, the enzyme can be used as a model to study the mechanism of interaction with the external fields. The Na, K-ATPase was used to find out how the fields could affect cells where the current flow around the cells rather than through them because of the high membrane resistance.
The ATPase splitting activity of the enzyme underwent a change when the fields applied to the samples, either increasing or decreasing, depending on the degree of activation by Na+, K+ ions. In either case, all the fields appeared to act by increasing the binding of activating ions; the effect on the enzyme depended on the increased binding decreased ATP-splitting activity, Fig. 5-9. The maximum ATP-splitting has occurred at 60 minutes of irradiation causing the decrease in specific activity and after this period the ATP-splitting decreases, causing the specific activity increasing. Figure 10 show that the effect of ATP-splitting is the maximum in all field. On the other hand, while the intrinsic activity increases until 30 min and remains approximately unchanged for magnetic and electromagnetic fields; there is no remarkable change for intrinsic activity in electric field (Fig. 11). The properties of Na, K-ATPase in magnetic and electromagnetic fields have led us to suggest a mechanism by which induced currents from electromagnetic fields can affect biochemical processes in cells.
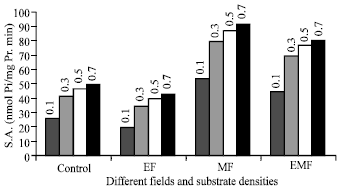 | |
| Fig. 5: | Specific activity as a function of substrate density in different fields for 15 min |
 | |
| Fig. 6: | The inhibition starts after 30 min and decrease in S.A. begins |
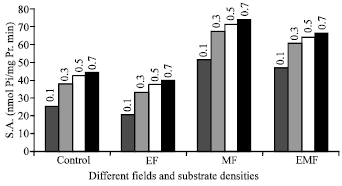 | |
| Fig. 7: | The S.A. is remarkably decreasing but still not at its minimum |
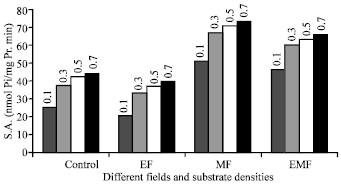 | |
| Fig. 8: | The S.A. is at its minimum after 60 min of irradiation |
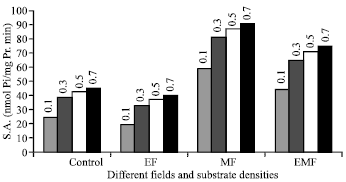 | |
| Fig. 9: | The S.A. after 90 min increased showing that the inhibition is no more effective |
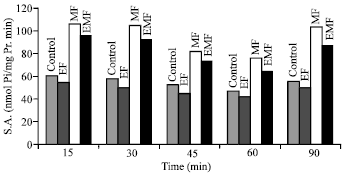 | |
| Fig. 10: | The variation of S.A. with different fields and with different substrate densities support well the maximum inhibition at 60 min of VDT usage |
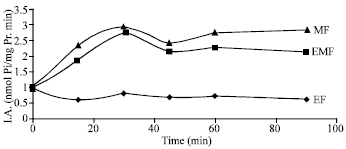 | |
| Fig. 11: | Intrinsic activity increases remarkably in 30 min and remains approximately unchanged for MF and EMF, but there is no remarkable change for EF |
Because ion-activation sites are on the outer as well as inner surfaces of the cell, signals do not have to penetrate the cell membrane to have an effect on the enzyme. Once the enzyme’s activity is affected, changes in ion pumping can alter the concentration of ions within the cell. Such a transition could explain how even weak electromagnetic signals that do not normally penetrate cell membranes are nevertheless transduced by cells, therefore that is why the effect for magnetic field is higher than the electromagnetic effect. The affect of the electromagnetic radiation caused severe irritated ears and eyes for the mousses (Fig. 12). Present study also may indicate why electromagnetic fields affect damaged brain tissue but do not affect normal tissue.
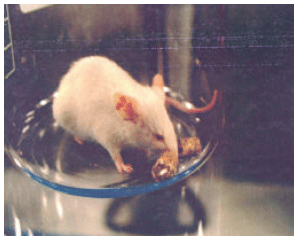 | |
| Fig. 12: | The fields’ affection demonstrated in the ears and eyes (ocular asthenopia) of the mousse |
This is because under normal conditions electromagnetic fields interferes only moderately with enzyme activity, but it greatly enhances activity when the enzyme functions under less than optimal conditions. In fact when tissue is damaged and leakage of ions results in altered ionic concentrations at the binding sites. This means, when conditions are no longer optimal the enzyme activity is reduced. Our study showed that the electromagnetic fields should remarkably increase the enzyme’s activity.
CONCLUSIONS
We have been able to obtain insight into the mechanism of action of electric, magnetic, and electromagnetic fields of the video display terminals on the Na, K-ATPase. The electric field may act directly on the enzyme, but the observed variation of enzyme activity under constant electric field suggests that the mechanism involves a change in ion binding to the activation sites. The result of changes in binding can be either an enhancement or decrease in enzyme activity. The greatest effects appear to be in the period of one hour use of the video display terminals. After this period, the effects decrease for all the fields. In fact, under optimal conditions all fields inhibit and when the enzyme activity is lowered to less than half its optimal level, there is an increase in activity. On the other hand, the applied fields show the effects that occur on the outer surface of the cell can transmit the information into the cell. Present study suggests that it can occur through the functioning of an enzyme that normally affects the ion gradients across the cell membrane.
Changes in the activation of the ion sites on the outside surface of the Na, K-ATPase change the ability of the enzyme to pump ions across the membrane. The enzyme can be activated or inhibited by passing currents along the outside surface, and the trans-membrane signaling occur via the effect on the ion fluxes and changes in the inner composition. The theoretical threshold for these effects is still being debated although is in accordance with experimental results and indicating that the magnitudes are quite low.
REFERENCES
- Abdollahzadeh, S., S.K. Hammond and M.B. Schenker, 1996. Validity of surrogates for determination of 30-1000 Hz magnetic field exposure for video display terminal users in office settings. Bioelectromagnetics, 17: 406-410.
Direct Link - Shahbazi-Moghadam, M. and E. Faghahati, 2006. Tritium concentration study of surface and ground water with a specially designed proportional-counting technique. Pak. J. Biol. Sci., 9: 681-685.
CrossRefDirect Link - Skou, J.C., 1957. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochem. Biophys. Acta, 23: 394-401.
CrossRefPubMedDirect Link