
Ahmad Ali Enayati
School of Public Health and Environmental Health Research Centre,
Mazandaran University of Medical Sciences, Sari, Iran
Hossein Ladonni
School of Public Health, Tehran University of Medical Sciences, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1265-1270







ABSTRACT
Bioassays were performed on 1-2 days old sugar fed adult mosquitoes of field and susceptible Beech strains of An. stephensi using WHO standard kit with some modifications. The KT50s of exposure to 0.08% permethrin impregnated papers were 40±1, 43±1 and 44±2 for susceptible, Kazeroun and Bandar Abbas strains, respectively. Biochemical assays of esterases, monooxigenases and glutathione S-transferases were undertaken on An.stephensi from Kazeroun and Bandar Abbas and the results were compared to those of the susceptible Beech strain. The mean alfa-esterase activity of Kazeroun, Bandar Abbas and Beech strains were 6.5×10-2, 7.1×10-2 and 6.1×10-2; the mean beta-esterase activity in those strains were 7.2×10-3, 3.2×10-3 and 2.4×10-3; the mean content of p450 in the above-mentioned strains were 8×10-3, 4.6×10-3 and 3.4×10-3; the mean GST activity were 8.7, 10.5 and 3.6, respectively. Although according to the results of this study, An. stephensi from Iran is not considered to be resistant to pyrethroids, the results of bioassays and the mean activity of all enzyme groups in field strains were significantly different from those of the susceptible strain. The results of bioassays were significantly different from those performed some 15 years ago on the same strains. Hence, insecticide resistance monitoring techniques should be put in place and also resistance management strategies and measures should be considered implementing in the area to prevent or postpone insecticide resistance development.
PDF Abstract XML References Citation
How to cite this article
Ahmad Ali Enayati and Hossein Ladonni, 2006. Biochemical Assay Baseline Data of Permetrhrin Resistance in Anopheles stephensi (Diptera, Culicidae) from Iran. Pakistan Journal of Biological Sciences, 9: 1265-1270.
DOI: 10.3923/pjbs.2006.1265.1270
URL: https://scialert.net/abstract/?doi=pjbs.2006.1265.1270
DOI: 10.3923/pjbs.2006.1265.1270
URL: https://scialert.net/abstract/?doi=pjbs.2006.1265.1270
INTRODUCTION
Anopheles stephensi is a serious urban vector of malaria in many countries in the Middle East and the Indian subcontinent. There are reports of pyrethroid resistance or tolerance in An. stephensi in the region (Ladonni, 1988; Vatandoost, 1996; WHO, 2000; Ganesh et al., 2003). Laboratory selection of An. stephensi with pyrethroids lead to resistance in as few as 5 generations (Kumar et al., 1991; Chakravorthy and Kalyanasundaram, 1992). In some strains of An. stephensi DDT-resistance produced cross-resistance to pyrethroids and selection of DDT-resistant An. stephensi by pyrethroids produced higher levels of resistance to both insecticides (Omer et al., 1980; Ladonni, 1988; Vatandoost et al., 1996). In strains from Iran and Pakistan, however, kdr-type cross-resistance was not observed (Malcolm, 1990; Enayati, 1992). When the KDR site-insensitivity resistance mechanism is present, cross-resistance between DDT and pyrethroids occurs. In the DUB-S strain of An. stephensi and a strain from Pakistan, DDT and pyrethroid susceptibility was not fully restored by synergists like dichlorophenyl ethanol (DMC), piperonyl butoxide (PBO) and S,S,S-tributylphosphorothiolate (DEF), suggesting that a site insensitivity mechanism was present (Omer et al., 1980; Ladonni, 1988; Vatandoost, 1996; Enayati et al., 2003).
Monooxygenases, esterases and glutathione S-transferases are the three major enzyme groups involved in metabolic insecticide resistance (Oppenoorth et al., 1977; 1979; Hemingway and Karunaratne, 1998; Scott, 1999). Monooxygenase involvement in pyrethroid resistance has been well studied in different vectors (Scott and Georghiou, 1985, 1986; Ladonni, 1988; Kumar et al., 1991; Anspaugh et al., 1994; Vatandoost, 1996; Liu and Scott, 1998; Kasai and Scott, 2000; Valles et al., 2000). Highly elevated esterases, due to gene amplification in Culex, are only important in OP and carbamats resistance (Karunaratne et al., 1993; Vaughan et al., 1995; De Silva et al., 1997; Small et al., 1998; Hemingway and Ranson, 2000; Paton et al., 2000). However, the elevated E4 and FE4 esterases in aphid M. persicae contribute at least marginally to permethrin resistance (Devonshire and Moores, 1982; Devonshire et al., 1998). In some other agricultural insect pests e.g. Helicoverpa armigera, H. punctigera and Bemisia tabaci some permethrin hydrolysing esterases have been characterised (Gunning et al., 1996; 1997; 1999). Although for the new generations of α-cyano-pyrethroids, esterases are more important resistance route (Casida et al., 1997), they have not been as well studied as monooxygenase-based pyrethroid resistance. The route of glutathione S-transferase-based pyrethroid resistance in insects has only recently been determined (Vontas et al., 2001; 2002). In this study the susceptibility status of the field collected strains of An. stsphensi from Iran and their potential biochemical resistance mechanisms are studied.
MATERIALS AND METHODS
Mosquito strains: Field strains of An. stephensi were collected from Bandar Abbas and Kazeroun, southern Iran and reared to the first generation in insectary under standard condition and used for bioassays and biochemical assays. Beech, a pyrethroid susceptible strain originated from India in 1940 and kept in the insectary without being exposed to insecticides.
Bioassays: World Health Organisation (WHO) standard kits for adult mosquito susceptibility tests with some modifications were used for bioassay experiments. Permethrin impregnated papers (0.08%) were prepared locally using pure olive oil, acetone and analytical grade permethrin. Groups of 25, 1-2 days old adults were placed into holding tubes and held for 30 min to control possible mortality due to handling. During this holding period, mosquitoes were provided with cotton pads moisturized with 10% sucrose solution. Exposure tubes containing insecticide impregnated papers were attached to the holding tubes and mosquitoes were gently transferred to the exposure tubes by blowing through the holding tubes. After a fixed exposure time, mosquitoes were transferred back to the holding tubes, provided with 10% sucrose and kept at 25°C and 70-80% relative humidity. Thirty min after exposure, the knockdown rates were scored and the data were subjected to probit analysis.
Biochemical assays: Biochemical assays for general esterases, glutathione S-tranferases and monooxyganases were performed on adult mosquito according to Hemingway (1998) with some modifications. Adult mosquitoes were individually homogenised in 200 μL of distilled water in a flat-bottomed microtitre plate on ice. The plate was spun at 3000 rpm for 15 min in a Beckman Culter centrifuge (Beckman) at 4°C and the supernatant was used as the source of enzymes in reaction mixtures. In each biochemical assay, blank replications (all components of the reaction mixture except for the enzyme source) were provided.
Monooxigenases assay: This assay measures the total amount of haem containing protein using a heam-peroxidase assay. As cytochrome P450s make up the bulk of the proteins in non-blood-fed insects, results can crudely be expressed as equivalent units of cytochrome P450 within the insect (Brogdon et al., 1999b). The reaction mixture in each well of the microtitre plate contained 20 μL of insect homogenate, 80 μL of 0.0625 M potassium phosphate buffer pH 7.2, 200 μL of 3, 3`, 5, 5` tetramethyl benzodine (TMBZ) solution (0.01 g TMBZ dissolved in 5 mL methanol plus 15 mL of 0.25 M sodium acetate buffer pH 5.0) and 25 μL of 3% hydrogen peroxide. The plates were incubated at room temperature for 2 h and absorbance was read at 450 nm as an endpoint in a Biotek ELX 808U plate reader. The values were compared with a standard curve of purified cytochrome C and were reported as equivalent units of cytochrome P450/mg protein corrected for the known content of cytochrome C and P450.
General esterases: General esterases activities with the substrates α- and β-naphthyl acetate were determined. Reaction mixtures contained 20 μL of insect homogenate in duplicate in adjacent microtitre plate wells (assigned α and β) and 200 μL of α- or β-naphthyl acetate solution (120 μL of 3 0 mM α or β-naphthyl acetate dissolved in 12 mL 0.02 M phosphate buffer pH 7.2), respectively. The reaction mixtures were incubated at room temperature for 30 min before the addition of 50 μL of fast blue solution (0.023 g fast blue dissolved in 2.25 mL distilled water and 5.25 mL of 5% SDS 0.1 M sodium phosphate buffer pH 7) to each well. Plates were incubated at room temperature for 5 min and then absorbance was read at 570 nm as an endpoint value. The resulting optical densities (OD) were compared with standard curves of ODs for known concentrations of the products α- and β-naphthol, respectively to convert the absorbances to product concentrations. The enzyme activities were reported as nmol of product formed/min/mg protein.
Glutathione S-transferases: Glutathione S-tansferase activity was measured using a reaction mixture of 10 μL of the homogenate plus 200 μL of reduced glutathione (GSH) and 1-chloro-2, 4-dinitrobenzene (CDNB) solution (10 mM reduced glutathione dissolved in 0.1 M phosphate buffer pH 6.5 and 3 mM CDNB originally dissolved in 1 mL methanol). The increase in absorbance was measured at 340 nm for 5 min. The enzyme activity was reported as μΜ/min/mg protein using the extinction co-efficient corrected for the path length of the solution in the microtitre plate well.
RESULTS
Bioassays: The results of bioassays performed on adult An. stephensi using 0.08% permethrin impregnated papers were subjected to probit analysis and summarised in Table 1. The bioassays shows that there are statistically significant differences between the KT50 of Bandar Abbas and Kazeroun field strains with that of Beech (p<0.05). Also the results show that the order of KT50 of the strains is Bandar abbas>Kazeroun>Beech.
Biochemical Assays: Monooxigenase, general esterase and glutathione S-transferase assays were carried out on adults of Bandar Abbas, Kazeroun and Beech strains of An. stephensi.
| Table 1: | The results of probit analysis on the bioassay results performed on adults of different strains of An. stephensi from Iran compared with a susceptible strain Beech |
 | |
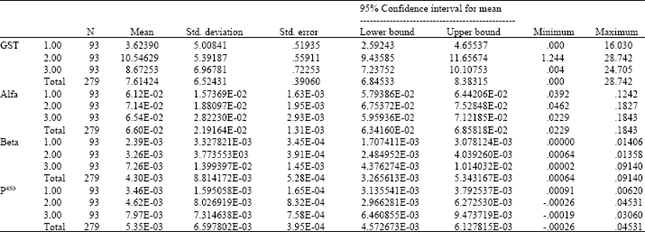
| Table 2: | Descriptive analysis of the results of biochemical assays performed on field and susceptible strains of An. stephensi from Iran. GST: Glutathione transferases, Alfa: Alfa esterases, Beta: Beta esterases, P450: Cytochrome P450. 1: Beech susceptible strain, 2: Kazeroun strain, 3: Bandar Abbas strain |
 | |
| Table 3: | One way ANOVA analysis of the results of biochemical assays performed on field and susceptible strains of An. stephensi from Iran. GST: Glutathione transferases, Alfa: Alfa esterases, Beta: Beta esterases, P450: Cytochrome P450. 1: Beech susceptible strain, 2: Kazeroun strain, 3: Bandar Abbas strain |
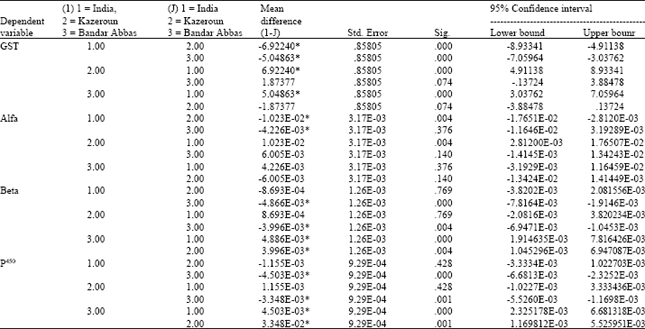 | |
| * The mean difference is significant at 0.05 level | |
All raw data were analysed using formulas and standard curves in Microsoft Excel software and reported as the equivalent units of cytochrome P450 for monooxigenase and activity rates for the latter two enzymes. The mean alfa-esterase activity of Bandar Abbas, Kazeroun and Beech strains were 6.5 x 10-2, 7.1 x 10-2 and 6.1 x 10-2; the mean beta-esterase activity in those strains were 7.2 x 10-3, 3.2 x 10-3 and 2.4 x 10-3; the mean content of p450 in the above-mentioned strains were 8 x 10-3, 4.6 x 10-3 and 3.4 x 10-3; the mean GST activity were 8.7, 10.5 and 3.6, respectively. Analysis of variance of the data showed that the differences between the enzymes activities of the field and Beech strains were statistically significant (α<0.001) (Table 2 and 3).
DISCUSSION
Monitoring insecticide resistance and its underlying mechanisms are crucially important in vector control as well as insecticide resistance management. Classic bioassay is a powerful, cheap and relatively easy method of monitoring and measuring insecticide resistance, however, it lacks adequate sensitivity and does not give a clear picture of mechanisms involved in insecticide resistance. When bioassay is undertaken along with more sensitive biochemical assays, more information and solid evidence come out about the insecticide resistance status and its possible mechanisms. In this study, possible mechanisms of insecticide resistance were investigated using bioassays based on WHO standards and biochemical assays according to Hemingway (1998).
Although the results of bioassays performed on the field strains and the susceptible strain showed statistically significant differences, based on WHO criteria, the results revealed that there is neither a resistance nor even a tolerance to permethrin in the field strains of An. stephensi collected from southern Iran. However, it is obvious that long term use of pyrethroid insecticides will eventually lead to resistance and in fact pyrethroid resistance in An. stephensi has already been documented from many countries in the Middle East and Indian subcontinent (Ladonni, 1988; Kumar et al., 1991; Chakravorthy and Kalyanasundaram, 1992; Guillet et al., 1996; Vatandoost, 1996; Enayati et al., 2003; Ganesh et al., 2003). The results of the bioassays showed a clear and statistically significant increase compared to the results of a study on the same strains performed some 15 years ago (Enayati, 1992). This implies that a trend of building up pyrethroid resistance has been started and developed over the years and sooner or latter may lead to insecticide resistance.
Comparative analysis of the results of biochemical assays performed on the field strains and the susceptible strain Beech revealed that esterases, monooxygenases and glutathione S-transferases were at higher levels in the field strains. The increase in P450 quantity, beta esterases and glutathione S-transferases activities in the field strains were about three times as much as those in Beech susceptible strain. Therefore, biochemical assays confirm that esterases, monooxygenases and possibly glutathione S-transferases are involved in pyrethroid resistance in the field strains of An. stephensi from Iran. Involvement of these enzymes in pyrethroid resistance in mosquitoes is well documented in previous studies on the same species as well as other mosquitoes (Omer et al., 1980; Ladonni, 1988; Kumar et al., 1991; Chakravorthy and Kalyanasundaram, 1992; Enayati, 1992; Vatandoost, 1996; Vulule et al., 1996, 1999; Enayati and Ladonni, 1997; Enayati et al., 2003; Ganesh et al., 2003).
The activity of beta esterase and the quantity of P450 in Bandar Abbas strain is much more than that of the susceptible strain as well as Kazeroun strain. However, the activities of alfa esterase and glutathione transferase in Kazeroun strain are only slightly more than those of the Bandar Abbas strain. This might be because the population of An. stephensi from Bandar Abbas has been under more insecticide selection pressure than the Kazeroun population. These findings are in accord with the results of other studies that esterases and oxidases are mainly involved in pyrethroid resistance (Oppenoorth et al., 1977; 1979; Scott and Georghiou, 1985; 1986; Ladonni, 1988; Kumar et al., 1991; Anspaugh et al., 1994; Vatandoost, 1996; Hemingway and Karunaratne, 1998; Liu and Scott, 1998; Scott, 1999; Kasai and Scott, 2000; Valles et al., 2000).
A quick look at the levels of the enzymes in the field and susceptible strains shows that the order of importance of the enzymes in conferring insecticide resistance is beta esterases, glutathione transferases and P450. Involvement of esterases, glutathione transferases and cytochrome oxidases in pyrethroid redidtance have been identified in various mosquito species and strains (Jianhua and Jialiang, 1987a, b; Ladonni, 1988; Kumar et al., 1991; Enayati, 1992; Sahgal et al., 1994; Vatandoost, 1996; Vulule et al., 1996, 1999; Enayati and Ladonni, 1997; Kasai et al., 1998; Brogdon et al., 1999a; Kasai and Scott, 2000; Enayati et al., 2003; Enayati and Hemingway, 2006).
There is a great deal of coordination between the results of bioassays and biochemical assays, each confirming the results of the other. The resistance ratios obtained by bioassays shows slower increase compared with that of biochemical assays. This is at least partly because biochemical assays are rather more sensitive and give a more clear picture of resistance before it is too late. In other words, when resistance is confirmed by bioassays, the gene frequency is at such a level that no resistance management strategies can easily be put in place with success. The indications that biochemical assays give at much higher levels, helps the authorities and experts in implementing vector control and resistance management strategies in time and with higher chance of success.
Although no resistance to insecticide has been reported for An. stephensi from Iran, but the trend of increase in the level of enzymes responsible for insecticide resistance should be a cause for concern as this means that the process of building up tolerance and finally resistance to insecticides has already been started. In fact detection of a 3-fold increase in the activity or amount of the esterases, glutathione transferases and P450 in the field strains compared to the susceptible strain should encourage the authorities and the experts to think of implementing resistance management strategies such as rotation, mosaics, mixture, change or any other methods in order to prevent or postpone the process of insecticide resistance development.
ACKNOWLEDGMENT
This study was funded by a grant from Mazandaran Management and Planning Organization, grant No. 404/26606.
REFERENCES
- Anspaugh, D.D., R.L. Rose, P.G. Koehler, E. Hodgson and R.M. Roe, 1994. Multiple Mechanisms of pyrethroid resistance in the German Cockroach Blattella germanica (L.). Pesticide Biochem. Physiol., 50: 138-140.
Direct Link - Brogdon, W.G., J.C. McAllister, A.M. Corwin and C. Cordon-Rosales, 1999. Independent selection of multiple mechanisms for pyrethroid resistance in Guatemalan Anopheles albimanus (Diptera: Culicidae). J. Econ. Entomol., 92: 298-302.
Direct Link - Brogdon, W.G., J.C. McAllister, A.M. Corwin and C. Cordon-Rosales, 1999. Oxidase-based DDT-pyrethroids cross-resistance in Guatemalan Anopheles albimanus. Pesticide Biochem. Physiol., 64: 101-111.
Direct Link - Chakravorthy, B.C. and M. Kalyanasundaram, 1992. Selection of permethrin resistance in the malaria vector Anopheles stephensi. Indian J. Malariol., 29: 161-165.
PubMed - De Silva, D., J. Hemingway, H. Ranson and A. Vaughan, 1997. Resistance to insecticides in insect vectors of Disease: Estα3, a novel amplified esterase associated with Amplifiedest β1 from insecticide resistant strains of the mosquito Culex quinquesfasciatus. Exp. Parasitol., 87: 253-259.
CrossRefDirect Link - Devonshire, A.L., L.M. Field, S.P. Foster, G.D. Moores, M.S. Williamson and R.L. Blackman, 1998. The evolution of insecticide resistance in the peach-potato aphid, Myzus persicae. Phil. Trans. R. Soc. Lond. B, 353: 1677-1684.
Direct Link - Enayati, A.A., H. Vatandoost, H. Ladonni, H. Townson and J. Hemingway, 2003. Molecular evidence for a kdr-like pyrethroid resistance mechanism in the malaria vector mosquito Anopheles stephensi. Med. Vet. Entomol., 17: 138-144.
PubMedDirect Link - Ganesh, K.N., J. Urmila, P. Guillet and L. Manga, 2003. Pyrethroid susceptibility and enzyme activity in two malaria vectors, Anopheles stephensi (Liston) and A. culicifacies (Giles) from Mysore, India. Ind. J. Med. Res., 117: 30-38.
Direct Link - Gunning, R.V., G.D. Moores and A.L. Devonshire, 1996. Esterases and esfenvalerate resistance in Australian helicoverpa armigera (Hubner) lepidoptera noctuidae. Pesticide Biochemis. Physiol., 54: 12-23.
CrossRef - Gunning, R.V., G.D. Moores and A.L. Devonshire, 1997. Esterases and fenvalerate resistance in a field population of Helicoverpa punctigera (Lepidoptera: Noctuidae) in Australia. Pesticide Biochem. Physiol., 58: 155-162.
Direct Link - Gunning, R.V., G.D. Moores and A.L. Devonshire, 1999. Esterase inhibitors synergise the toxicity of pyrethroids in Australian Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae). Pesticide Biochem. Physiol., 63: 50-62.
Direct Link - Hemingway, J. and S.H. Karunaratne, 1998. Mosquito carboxylesterases: A review of the molecular biology and biochemistry of a major insecticide resistance mechanism. Med. Vet. Entomol., 12: 1-12.
PubMed - Hemingway, J. and H. Ranson, 2000. Insecticide resistance in insect vectors of human disease. Ann. Rev. Entomol., 45: 371-391.
CrossRefPubMedDirect Link - Karunaratne, S.H.P.P., K.G.I. Jayawardena, J. Hemingway and A.J. Ketterman, 1993. Characterization of a B-type esterase involved in insecticide resistance from the mosquito Culex quinquefasciatus. Biochem. J., 294: 575-579.
PubMedDirect Link - Kasai, S., I.S. Weerasinghe and T. Shono, 1998. P450 Monooxygenases are an important mechanism of permethrin resistance in Culex quinquefasciatus say larvae. Arch. Insect Biochem. Physiol., 37: 47-56.
CrossRef - Liu, N. and J.G. Scott, 1998. Increased transcription of CYP6D1 causes cytochrome P450 mediated insecticide resistance in house fly. Insect Biochem. Mol. Biol., 28: 571-580.
Direct Link - Malcolm, C.A., 1990. Location of a gene conferring DDT resistance but no pyrethroid cross resistance in larvae of Anopheles stephensi. Genetica, 82: 51-55.
CrossRef - Paton, M.G., S.H. Karunaratne, E. Giakoumaki, N. Roberts and J. Hemingway, 2000. Quantitative analysis of gene amplification in insecticide-resistant Culex mosquitoes. Biochem. J., 346: 17-24.
PubMed - Sahgal, A., S. Kumar and M.K.K. Pillai, 1994. Microplate Assay of elevated esterase-activity in individual pyrethroid-resistant mosquitos. J. Biosci., 19: 193-199.
CrossRef - Scott, J.G., 1999. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol., 29: 757-777.
CrossRefDirect Link - Valles, S.M., K. Dong and R.J. Brenner, 2000. Mechanisms responsible for cypermethrin resistance in a strain of German cockroach, Blattella germanica. Pestic. Biochem. Physiol., 66: 195-205.
CrossRefDirect Link - Vaughan, A., M. Rodriguez and J. Hemingway, 1995. The independent gene amplification of indistinguishable esterase B electromorphs from the insecticide resistant mosquito Culex quinquefasciatus. Biochem. J., 305: 651-658.
Direct Link - Vontas, J.G., G.J. Small and J. Hemingway, 2001. Glutathione S-transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem. J., 357: 65-72.
Direct Link - Vontas, J.G., G.J. Small, D.C. Nikou, H. Ranson and J. Hemingway, 2002. Purification, molecular cloning and heterologous expression of a glutathione S-transferase involved in insecticide resistance from the brown plant hopper Nilaparvata lugens. Biochem. J., 362: 329-337.
PubMedDirect Link - Vulule, J.M., R.F. Beach, F.K. Atieli, D.L. Mount, J.M. Roberts and R.W. Mwangi, 1996. Long-term use of permethrin-impregnated nets does not increase Anopheles gambiae permethrin tolerance. Med. Vet. Entomol., 10: 71-79.
Direct Link - Vulule, J.M., R.F. Beach, F.K. Atieli, J.C. McAllister and W.G. Brogdon et al., 1999. Elevated oxidase and esterase levels associated with permethrin Tolerance in Anopheles gambiae from Kenyan villages using permethrin-impregnated nets. Med. Vet. Entomol., 13: 239-244.
Direct Link - Omer, S.M., G.P. Georghiou and S.N. Irving, 1980. DDT/pyrethroid resistance inter-relationships in Anopheles stephensi. Mosquito News, 40: 200-209.
Direct Link