
A. Cheriguene
Departement de Biologie, Facult6 des Sciences, Laboratoire de Microbiologie,
Universite de Mostaganem, BP 227 Mostaganem 27000 Algeria
F. Chougrani
Departement de Biologie, Facult6 des Sciences, Laboratoire de Microbiologie,
Universite de Mostaganem, BP 227 Mostaganem 27000 Algeria
A. Bensoltane
D6partement de Biologie, Facult6 des Sciences, Laboratoire de J.1icrobiologie Alimentaire,
Universit6 d Oran Es Senia. Algeria
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1242-1249







ABSTRACT
One hundred twenty strains of lactic acid bacteria were isolated from raw goat`s milk in Western Algeria. They were identified and technologically characterized. Of the isolates, 25.33% were classified as Lactococcus, 34.66% as Enterococcus, 37.33% as lactobacilli and 2.66% as Streptococcus thermophilus. The dominant lactococci species was Lactococcus lactis subsp. lactis (18.66%). Twenty five of the Enterococci isolates were identified as Enterococcus faecium, while the remaining 07.33% were identified as Enterococcus durans. Other species identified included Lactobacillus rhamnosus (10.66%), Lb. pentosus (09.33%), Lb. paracasei subsp. paracasei (08%), Lb. plantarum (05.33%), Lb. delbrueckii subsp. bulgaricus (04%). Strains were screened for their acidifying, proteolytic and antibacterial activities. The strains of L. lactis subsp. lactis exhibited the highest acidifying and proteolytic activity. The highest acidifying activity was found in L. lactis subsp. lactis LC2, while E. faecium EC3 exhibited the highest proteolytic activity. Antimicrobial activity was detected in eight strains of Enterococcus, seven Lactococcus and six Lactobacillus. Some strains were able to maintain a high activity of two or three technological characteristics together.
PDF Abstract XML References Citation
How to cite this article
A. Cheriguene, F. Chougrani and A. Bensoltane, 2006. Identification and Characterization of Lactic Acid Bacteria Isolated from Algerian Goat’s Milk. Pakistan Journal of Biological Sciences, 9: 1242-1249.
DOI: 10.3923/pjbs.2006.1242.1249
URL: https://scialert.net/abstract/?doi=pjbs.2006.1242.1249
DOI: 10.3923/pjbs.2006.1242.1249
URL: https://scialert.net/abstract/?doi=pjbs.2006.1242.1249
INTRODUCTION
Goats are mainly found in semi-mountainous, mountainous and arid areas in Algeria. Their population consists of local breeds that have not been genetically improved and as a result, their annual milk yield per animal is low. However, today almost 14% of milk produced annually derives from goat in Algeria (Anonymous, 2000).
These animals provide meat, milk and (or) fiber to many people of the world, playing a special role in achieving food and economic securities in developing regions. In recent years demand for goat products has increased in developed countries as well, which has been accompanied by a rising number of farmers raising goats (Sahlu et al., 2004).
Transformation of goat’s milk into traditional Algerian dairy products such as Raib Lben, Jben and many others is achieved through spontaneous fermentation without the addition of any selected starter. Such products generally present irregular sensorial qualities.
The lactic acid bacteria used in the production of different cheeses in Algeria come in the form of commercial starters composed of mesophilic bacteria. In most cases, these starters are not adapted to the manufacture conditions of the local products.
Lactic Acid Bacteria (LAB) are generally considered as safe in food products and are of major interest in the food industry especially in dairy products.
The LAB used in commercial starter cultures possesses numerous metabolic characteristics such as acidification activity, proteolytic activity, synthesis of bacteriocin and resistant to bacteriophage are strain dependant.
All of these important activities contribute to the flavour, texture and frequently the nutritional attributes of the products. Research has been focused on the role of starter and its required properties of the dairy industries (Crow et al., 1993; De Vuyst and Degeest, 1999).
The purpose of this study was to examine the lactic microflora which predominates throughout Algerian goat’s milk and to determine the technological properties of selected species. Such work was developed hoping that a starter consisting of` wild lactic acid bacteria isolates suitable for cheese manufacture will eventually be possible.
MATERIALS AND METHODS
Samples: Samples were collected from raw goat’s milk collected from different farms in West Algeria. The samples were immediately cooled and brought to the laboratory in an isotherm container, being analysed in arrival.
Isolation and identification of strains: The milk samples were incubated at 30, 37 and 42°C respectively. Coagulated samples were then streaked on MRS agar (De Man et al., 1960) used for isolation of LAB such as Lactobacillus, Leuconostoc and Pediococcus and M17 agar (Terzaghi and Sandine, 1975) used for isolation of Lactococcus and Enterococcus. Streaked samples on MRS were incubated on anaerobic conditions using the Gas Pak Plus Anaerobic System (Beckon Dickinson Microbiology System, Cockeysville, MD, USA) at 30, 37 and 42°C, respectively for 48 h. The isolates were purified by successive streaking on the appropriate agar media before being subjected to characterization.
The purified isolates were stored at -20°C in sterile reconstituted skim milk 12.5% (w/v) containing 15% glycerol for further analysis. Working cultures were also kept on MRS or M17 agar slant at 4°C and restreaked every 4 weeks (Samelis et al., 1994; Herrero et al., 1996).
Preliminary identification: Isolates were selected on the basis of Gram reaction, morphology, catalase activity and spore formation (Harrigan and McCance, 1976). Catalase-negative and Gram-positive rods and cocci were selected and screened for the production of CO2 from glucose determined in MRS or M17 broth containing inverted Durham tubes. The isolates were incubated for 2 to 5 days at 30°C. Arginine hydrolysis was performed on selective strains using the method of Schillinger and Lücke (1987).
Growth at different temperatures 15, 37 and 45°C was observed in MRS and M17 broth, respectively after 5 days of incubation and 12 days at 4 and 10°C (Schillinger and Lücke, 1987; Samelis et al., 1994). For cocci strains, growth on SF broth medium and in the presence of 6.5% NaCl was also considered.
The ability to grow at pH 3.9 and 9 was tested on MRS and M17 broth. Citrate utilization in the presence of carbohydrates was performed on the media of Kempler and Mc Kay (1980). Acetoïne production from glucose was determined by Voges-Proskauer test (Harrigan and McCance, 1976).
Phenotypic characterization: The carbohydrate fermentation was analysed in a MRS broth with bromocresol purple mixed at 0.02% as a pH indicator. Sugars were sterilized by filtration and added to make a final concentration of 1% (w/v). Samples were incubated at 37°C for 48 h on MRS broth without glucose but supplemented at a final concentration of 1% (w/v) with fructose, galactose, glucose, mannose, lactose, trehalose, cellobiose, xylose, raffinose, sorbitol, arabinose, melibiose and sucrose. To ensure anaerobic conditions, each tube was supplemented with two drops of sterile liquid paraffin after inoculation.
Hydrolysis of aesculin was also tested using the MRS broth supplemented with 2% aesculin (Milliere et al., 1989).
Further characterization of 25% of the isolates was carried out using the API 50 CH system (API System, bioMerieux, Marcy l’Etoile, France). Cells used as inoculum were prepared as recommended by the manufacturer. The results were recorded after 24 and 48 h and interpreted using the APILAB Plus computer-aided identification program (bioMerieux). A percentage correct identification (%Id) value was obtained for each strain and standard taxonomic descriptions from Wood and Holzapfel (1995).
Performances tests
Acidifying activity: Acidifying activity of the strains was measured according to the International Dairy Federation (IDF) standard 306 (IDF, 1995) and Allonso-Calleja et al. (2002). The strains were subcultured in MRS broth at 30°C for 24 h. The microbial culture was inoculated at a level of 1 mL 100 mL-1 in sterile skim powder milk 10% (w/v). Titratable acidity and pH were determined after 6, 12 and 24 h of incubation at 30°C.
Proteolytic activity: The proteolytic activity of the strains grown in milk was measured by the tyrosine method (International Dairy Federation Standard 149A, 1997; Brussels, Belgium). The proteolytic activity of the cultures results in liberation of the amino acids tyrosine and tryptophan from the milk substrate, which then react with the phenol reagent, yielding a blue colour which is measured at 650 nm. The results were calculated from a calibration curve obtained from dilutions of tyrosine in distilled water and expressed as μg tyrosine mL-1.
Antagonistic activity: Antagonistic activity was evaluated according to the protocol described by Geis et al. (1983). Cultures were grown overnight (16 h) in the suitable media at the optimum growth temperature. The overnight cultures were spotted (5 μL) onto appropriate agar medium. The plates were incubated for 48 h at the optimum temperature to allow producer colonies to develop, then overlaid with 3.5 mL of soft agar (0.75%) inoculated with the indicator organism at a concentration of 107 cfu mL-1 cells from a turbid overnight culture of the indicated target organisms. The plates were incubated under anaerobic conditions (GasPak, BBL, Cockeysville, MD, USA) at the optimum growth temperature for 24 h. Anaerobic incubation was used to rule out any inhibition due to hydrogen peroxide production (Lewus et al., 1991).
The plates were incubated 24 h at 30 or 37°C and checked for inhibition zones around the spot area after 24 h. Cultures belong to the same genus were interacted together. Each tested strain was applied as inhibitor organism while another one was taken as indicator organisms.
RESULTS AND DISCUSSION
Identification of isolates: Seventy five of lactic acid bacteria were isolated from goat’s milk. All isolates were Gram-positive, catalase-negative, oxydase-negative and non-spore-forming.
The morphological, physiological and biochemical tests revealed a diversity of lactic acid bacteria which were classified into the genera Lactococcus (19 strains, 25.33%), Enterococcus (26 strains, 34.66%), homofermentative lactobacilli (03 strains, 04%), facultatively heterofermentative lactobacilli (25 strains, 33.33%) and Streptococcus thermophilus (02 strains, 02.66%). Figure 1 shows the percentage distribution of the 75 bacteria isolated from goat’s milk in Western Algeria.
Among the cocci, 19 isolates were able to grow at 10 and 40°C, but not at 45°C and in pH 9.6 broth, except for two (LC4 and LC7). In addition, these strains did not survive at 60°C for 30 min. Fourteen strains could grow in 3 and 4%, but not in 6.5%, NaCl broth, produced NH3 from arginine and hydrolyzed esculin. The strains formed acid from glucose, galactose, fructose, lactose, trehalose and cellobiose but acid production from xylose, raffinose, sorbitol, arabinose and melibiose was negative. The phenotypic characteristics of the strains (Table 1) suggest their close resemblance to L. lactis subsp. lactis (Sharpe, 1979; Schleifer et al., 1985; Balows et al., 1991). The five remaining strains were classified as L. lactis subsp. cremoris.
In present study, most of Lactococcus isolated belonged to Lactococcus lactis subsp. lactis. According to other reports, Lactococcus lactis subsp. lactis was more frequently isolated than Lactococcus lactis subsp. cremoris from raw milk samples (Moreno and Busani, 1990) and Dahi and buttermilk samples from India (Padmanabha-Reddy et al., 1994). According to Holler and Steele (1995), Lactococcus lactis subsp cremoris was isolated only rarely from natural sources.
 | |
| Fig. 1: | Distribution of lactic acid bacteria isolated from Algerian goat’s milk |
According to Crow et al. (1993) and Weerkamp et al. (1996) lactococci isolated from natural sources were usually identified as Lactococcus lactis subsp. lactis, whereas the phenotype Lactococcus lactis subsp. cremoris, which is common in industrial mixed strain starter cultures, was isolated rarely (Beukes et al., 2001).
Twenty six isolates of cocci were able to grow at 10 and 45°C, in 6.5% NaCl and pH 9.6 broth; they also formed NH3 from arginine but not CO2 from glucose and were characterized as enterococci. Nineteen of them seemed to be E. faecium, as suggested by their ability to form acid from sugars (Sharpe, 1979; Schleifer and Kilpper-Balz, 1984; Devriese et al., 1991). Otherwise, six strains seemed to be E. durans by their inability to ferment melibiose and sucrose and unable, in general, to ferment sugars (Schleifer and Kilpper-Balz, 1984; Devriese et al., 1991).
The 28 isolates of Gram-positive rods grew at 15°C and did not form either CO2 from glucose or NH3 from arginine. These characteristics suggest the classification as facultatively heterofermentative lactobacilli (25 strains) and facultatively heterofermentative lactobacilli (03 strains) (Sharpe, 1979; Balows et al., 1991. Eight of 28 isolates did not form acid from xylose, raffinose and sucrose and were characterized as Lb. rhamnosus (Collins et al., 1991; Balows et al., 1991). Six isolates were able to form acid from nearly all sugars; they were characterized as Lb. paracasei subsp. paracasei.
Four isolates of rods were classified as Lb. plantarum, as suggested by their sugar fermentation patterns. All strains fermented cellobiose, lactose, maltose, melibiose, raffinose, lactose, maltose, sucrose and trehalose (Sharpe, 1979; Balows et al., 1991). The strains did not form acid from xylose and acid production from arabinose was variable and strain dependent.
| Table 1: | Phenotypic characteristics of the isolated strains* |
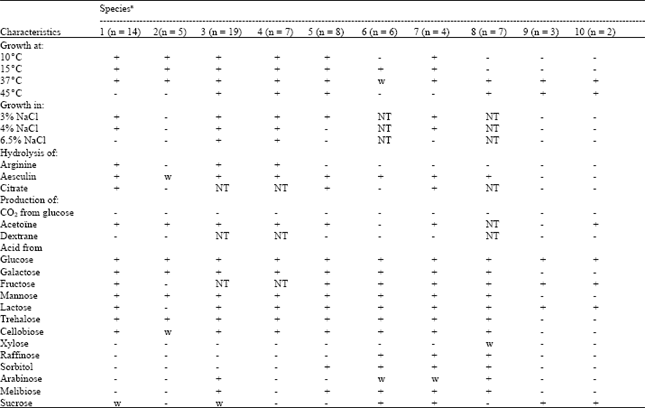 | |
*: All strains were Gram-positive, catalase-negative and non-spore-forming, w, weak reaction. NT: Not Tested, aSpecies: 1. Lc. lactis subsp. lactis; 2. Lc. lactis subsp. cremoris; 3. En. faecium; 4. En. durans; 5. Lb. rhamnosus; 6. Lb. paracasei subsp. paracasei; 7. Lb. plantarum; 8. Lb. delbrueckii bulgaricus; 9. Lb. pentosus; 10. S. thermophilus | |
The seven isolates classified as Lb. pentosus were characterized by their incapacity to grow in 10 and 15°C and by their ability to form acid from all sugars.
The two strains identified as Lb. delbrueckii subsp. bulgaricus were characterized by their sensibility to NaCl and their inability to ferment trehalose, cellobiose, xylose, raffinose, sorbitol, arabinose, melibiose, galactose and mannose.
Two strains were considered as Streptococcus thermophilus to their inability to grow at 15 and 30°C. They are heat-resistant, do not produce acetoïn and do not hydrolyse aesculin. This species is included in starter cultures during the production of yoghurt and cheese manufacture.
The diversity of lactic acid bacteria isolated from raw milk could be explained by the fact that samples were collected from different areas and different varieties of goats. Thus, this diversity is relative and dependent primarily on the nature of the material isolated and the different criteria used from each study, as reported by Fitzsimons et al. (1999) and Bissonnette et al. (2000).
Technological characteristics of strains
Acidifying activity: The strains of L. lactis subsp. lactis were those which showed the highest acidifying capacity, developing an acidity of 0.39 g 100 mL-1 lactic acid after 6 h and 0.50-0.70 g 100 mL-1 after 12 h (Table 2). After 24 h, the pH of the culture medium decreased to values lower than 4.1 and the acidity reached values around 0.7-0.8 g 100 mL-1. Basically, most the differences with respect to acidifying capacity were observed after 6 h.
All the strains had a similar behavior after 24 h, although the LC 2, LC 5 and LC 6 strains showed the numerically highest acidifying capacity. Lb. rhamnosus showed an acidifying capacity similar to that of L. lactis subsp. lactis strains throughout the incubation time. L. lactis subsp. cremoris showed a relatively low acidifying capacity, with average values of acidity of 0.22-0.32 g 100 mL-1 after 6 h. The strains of this species showed differences after 12 and 24 h and LC 12 was the strain with the highest acidifying capacity after 24 h (0.69 g 100 mL-1) (Table 2 and Fig. 2).
The acidifying ability of enterococci were in general low, showing a lower acidifying capacity than L. lactis subsp. lactis and developed an acidity of 0.58-0.61 g 100 mL-1 lactic acid after 24 h incubation. A significant difference was observed only among the strains of E. faecium EC 3 after 12 h.
| Table 2: | Acidifying activity of some strains of lactic acid bacteria isolated from Algerian goat’s milk |
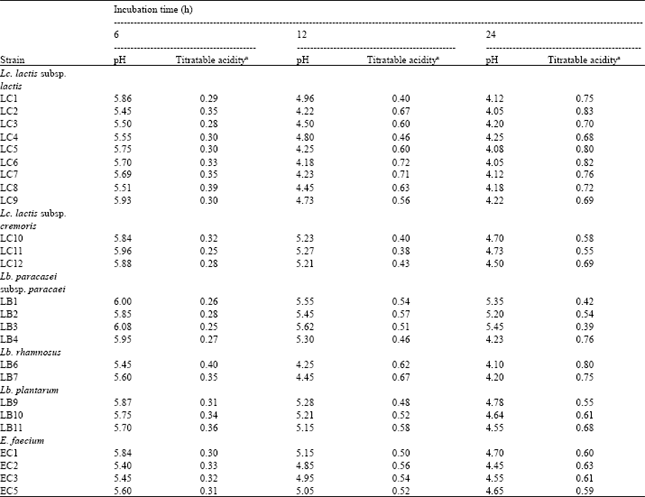 | |
| aTitratable acidity expressed as g 100 mL-1 lactic acid | |
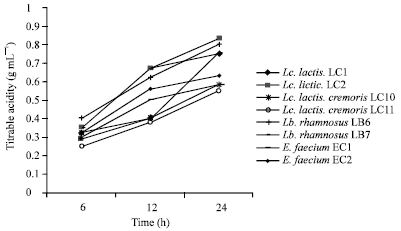 | |
| Fig. 2: | Acid production of some LAB strains tested in this study |
The acidification developed by strains of L. paracasei subsp. paracasei was lower than that produced by most of the strains of L. plantarum after 24 h. The strain L. paracasei subsp. paracasei LB4, after 24 h of incubation, was developing acidity as rapidly as some lactococci (Table 2). Lactobacilli metabolize lactose more slowly than Lactococci but the final acid production can be similar to, or even higher than, that of the Lactococci.
The strains of L. lactis subsp. lactis isolated from goat’s milk showed an acidifying activity after 6 h of incubation similar to that detected for lactococci isolated from goats’ milk cheeses by Requena et al. (1991) which suggested that strains of L. lactis subsp. lactis whose acidifying capacity are higher than 0.25 g 100 mL-1 of lactic acid after 6 h of incubation could be used as starter culture in cheese manufacture.
The differences observed from one lactic acid bacteria species to another were explained by De Roissart (1986). In fact, the acidifying acidity if each strain is related to its specific capacity to break down the substances in the medium and render them capable of assimilation. On occasion, differences are also due to the presence or absence of nutrient transport systems (Albenzino et al., 2001).
Proteolytic activity: Our test strains were characterized by different caseinolytic breakdown ability (Fig. 3). The range of proteolytic activity was 18-65.3 μg tyrosine mL-1 for L. lactis subsp. lactis strains. The proteolytic activity of the enterococci strains was measured at levels between 29.3 (E. faecium strain EC2) and 81.6 μg mL-1 (E. faecium strain EC3) and ~36% of the strains had activity > 55 μg tyrosine mL-1. The proteolytic activity of lactobacilli ranged between 23.4 (strains LB3 and LB4) and 51.3 μg tyrosine mL-1 (three strains). The data reported here on proteolytic activity suggest that there was no relationship between the proteolytic and acidifying activities of the strains, as also suggested by Fortina et al. (1998) for strains of lactobacilli and Durlo-Ozkaya et al. (2001). Thus, Lactobacilli strains LB and LB4 and enterococci strains EC2 and EC3 revealing good acidifying abilities did not exhibit the highest proteolytic activities and there were strains with very low acidifying but high proteolytic activity (e.g. E. faecium strain EC3) and strains with high acidifying and proteolytic activity (for example, L. lactis subsp. lactis strain LC6). The proteinase system of Lactococcus has been studied for several years and consists of cell wall-bound proteinases and several peptidases (Bockelmann, 1995). Enzymes formed by Lactobacillus strains were studied in detail and many authors have described enzymes that were biochemically similar to those of Lactococcus and their importance for cheese ripening is obvious (Bockelmann, 1995). The proteolytic activity and acid production of Fig. 3. Proteolytic activities of lactic acid bacteria isolated from raw goat's milk. According to phenotypic characterization: LC, strains of Lactococcus; LB, Lactobacillus, EC, Enterococcus.enterococci during growth in milk are sometimes comparable to those of S. thermophilus (Gatti et al., 1994).
Antagonistic activity: Antimicrobial activity was determined in direct antagonistic test; the strains belonging to the same species were tested against each other in agar plates. All of the strains were used as potential producers and as indicators. The results showed that seven strains of Lactococcus were inhibitory against more than 50% of the indicator strains. Six strains of Lactobacillus and eight of Enterococcus appeared to have antimicrobial activity. Since various LAB can produce bacteriocins active against LAB from the same species or closely related species (Jack et al., 1995; Piard and Desmazeaud, 1992), these antimicrobial activity are likely to be a consequence of bacteriocin production. Twenty five percent of tested Lactococcus strains exhibited antagonistic activity against each other; most of these strains were L. lactis subsp. lactis and 10% L. lactis subsp. cremoris. Among the Lactobacillus, bacteriocins-producing strains were Lb. rhamnosus (2 strains), Lb. delb. subsp. bulgaricus (one strain), Lb. plantarum (2 strains), Lb. paracasei subsp. paracasei (one strain) and one strain of Lb. pentosus. Eight enterococci were bacteriocins producing strains; E. faecium (6 strains) and E. durans (2 strains).
These results indicated that the percentage of bacteriocins-producing strains in this study is higher than those previously reported by Cogan et al. (1997) and Estepar et al. (1999).
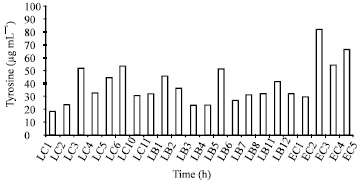 | |
| Fig. 3: | Proteolytic activities of lactic acid bacteria isolated from raw goat’s milk. According to phenotypic characterization: LC, strains of Lactococcus; LB, Lactobacillus, EC, Enterococcus |
Lactic acid bacteria known to produce antimicrobial substances such as organic acids, hydrogen peroxide, diacetyl and bacteriocins which are antagonistic towards spoilage and pathogenic organisms (Daeschel, 1989; Piard and Desmazeaud, 1992). Bacteriocin-producing strains have been used in starter cultures for cheese making to improve safety and quality of the cheese (Delves-Broughton et al., 1996) and recently, these strains are added with sensitive adjunct cultures to increase their autolysis in order to accelerate cheese ripening (Ross et al., 1999). Further research needs to focus on characterization and identification of the antimicrobial compounds produced by selected strains before use as starter cultures.
REFERENCES
- Albenzino, M., M.R. Corbo, S.U. Rehman, P.F. Fox and M. de Angelis et al., 2001. Microbiological and biochemical characteristics of Cane Strato Pugliese cheese made from raw milk, pasteurized milk or by heating the curd in hot whey. Int. J. Food. Microbiol., 67: 35-48.
CrossRef - Allonso-Calleja, C., J. Carballo, R. Capita, A. Bernardo and M.L. Garcia-Lopez, 2002. Comparison of the acidifying activity of Lactococcus lactis subsp. lactis strains isolated from goat's milk and Valdeteja cheese. Lett. Applied Microbiol., 34: 134-138.
PubMed - Beukes, E.M., B.H. Bester and J.F. Mostert, 2001. The microbiology of South African traditional fermented milks. Int. J. Food Microbiol., 63: 189-197.
CrossRefPubMedDirect Link - Bissonnette, F., S. Labrie, H. Deveau, M. Lamoureux and S. Moineau, 2000. Characterization of mesophilic mixed starter cultures used for the manufacture of aged cheddar cheese. J. Dairy Sci., 83: 620-627.
Direct Link - Bockelmann, W., 1995. The proteolytic system of starter and non-starter bacteria: Components and their importance for cheese ripening. Int. Dairy J., 5: 977-994.
CrossRef - Cogan, T.M., M. Barbosa, E. Beuvier, B. Bianchi-Salvadori and P.S. Cocconcelli et al., 1997. Characterization of lactic acid bacteria in artisanal dairy products. J. Dairy Res., 64: 409-421.
CrossRefDirect Link - Crow, V.C., T. Coolbear, R. Holland, G.G. Pritchard and F.G. Martley, 1993. Starters as finishers: Starter properties relevant to cheese ripening. Int. Dairy J., 3: 423-460.
Direct Link - Daeschel, M.A., 1989. Antimicrobial substances from lactic acid bacteria for use as food preservatives. Food Technol., 43: 164-167.
Direct Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Delves-Broughton, J., P. Blackburn, R.J. Evans and J. Hugenholtz, 1996. Applications of the bacteriocin, nisin. Antonie van Leeuwenhock, 69: 193-202.
CrossRefDirect Link - Devriese, L.A., B. Pot and M.D. Collins, 1993. Phenotypic identification of the genus Enterococcus and differentiation of phylogenetically dinstinct enterococcal species and species groups. J. Applied Bacteriol., 75: 399-408.
PubMed - De Vuyst, L. and B. Degeest, 1999. Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev., 23: 153-177.
CrossRefPubMedDirect Link - Durlu-Ozkaya, F., V. Xanthopoulos, N. Tunail and E. Litopoulou-Tzanetaki, 2001. Technologically important properties of lactic acid bacteria isolates from Beyaz cheese made from raw ewes' milk. J. Applied Microbiol., 91: 861-870.
PubMed - Estepar, J., M.M. del Sanchez, L. Alonso and B. Mayo, 1999. Biochemical and microbiological characterization of artisan Penamellera cheese: Analysis of its indigenous lactic acid bacteria. Int. Dairy J., 9: 737-746.
CrossRef - Fitzsimons, N.A., T.M. Cogan, S. Condon and T. Beresford, 1999. Phenotypic and genotypic characterization of non-starter lactic acid bacteria in mature cheddar cheese. Applied Environ. Microbiol., 65: 3418-3426.
Direct Link - Fortina, M.G., G. Nicastro, D. Garminati, E. Neviani and P.L. Manachini, 1998. Lactobacillus helveticus heterogeneity in natural cheese starters: The diversity in phenotypic characteristics. J. Applied Microbiol., 84: 72-80.
Direct Link - Geis, A., J. Singh and M. Teuber, 1983. Potential of lactic Streptococci to produce bacteriocin. Applied Environ. Microbiol., 45: 205-211.
Direct Link - Herrero, M., B. Mayo, B. Gonzalez and J.E. Suarez, 1996. Evaluation of technologically important traits in lactic acid bacteria isolated from spontaneous fermentation. J. Applied Bacteriol., 82: 565-570.
Direct Link - Holler, B.J and J.L. Steele, 1995. Characterization of lactococci other than Lactococcus lactis for possible use as starter cultures. Int. Dairy J., 5: 275-289.
CrossRef - Jack, R.W., J.R. Tagg and B. Ray, 1995. Bacteriocins of gram-positive bacteria. Microbiol. Rev., 59: 171-200.
Direct Link - Kempler, G.M. and L.L. McKay, 1980. Improved medium for detection of citrate-fermenting Streptococcus lactis subsp. diacetylactis. J. Applied Environ. Microbiol., 39: 926-927.
PubMedDirect Link - Lewus, C.B., A. Kaiser and T.J. Montville, 1991. Inhibition of food-borne bacterial pathogens by bacteriocins from lactic acid bacteria isolated from meat. Applied Environ. Microbiol., 57: 1683-1688.
PubMedDirect Link - Padmanabha-Reddy, V., M.M. Habibulla-Khad and V. Purushothaman, 1994. Plasmid linked starter characteristics in lactococci isolated from dahi and buttermilk. Cult. Dairy Prod. J., 29: 25-30.
Direct Link - Piard, J.C. and M. Desmazeaud, 1992. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances. Lait, 72: 113-142.
CrossRefDirect Link - Requena, T., C. Pel!aez and M.J. Desmazeaud, 1991. Characterization of lactococci and lactobacilli isolated from semi-hard goats cheese. J. Dairy Res., 58: 137-145.
Direct Link - Ross, P., M. Galvin, O. McAuliffe, S. Morgan and M. Ryan et al., 1999. Developing applications for lactococcal bacteriocins. Antonie van Leeuwenhoek, 76: 337-346.
PubMed - Samelis, J., F. Maurogenakis and J. Metaxopoulos, 1994. Characterization of lactic acid bacteria isolated from naturally fermented Greeck dry salami. Int. J. Food. Microbiol., 23: 179-196.
PubMedDirect Link - Schillinger, U. and F.K. Lucke, 1987. Identification of lactobacilli from meat and meat product. Food Microbiol., 4: 199-208.
CrossRef - Schleifer, K.H. and R. Kilpper-Balz, 1984. Transfer of Streptococcus faecalis and Streptococcus faecium to the genus Enterococcus nom. rev. as Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. Int. J. Syst. Bacteriol., 34: 31-34.
CrossRef - Terzaghi, B.E. and W.E. Sandine, 1975. Imoroved medium for lactic Streptococci and their bacteriophages. Applied Environ. Microbiol., 29: 807-813.
PubMedDirect Link - Weerkamp, A.H., N. Klijn, R. Neeter and G. Smit, 1996. Properties of mesophilic lactic acid bacteria from raw milk and naturally fermented raw products. Neth. Milk Dairy J., 50: 319-322.
Direct Link