
F. Mahmoodi Kordi
A. Majd
M. Valizadeh
M. Sheidai
H. Sabaghpour
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1225-1230







ABSTRACT
Karyotypic analysis was performed for 11 genotypes of chickpea (Cicer arietinum L.) in order to determine their potential genetic relationships. Pretreatment of root tips in 8-hydroxyquinoline, followed by fixation in Lewitsky solution and hydrolysis in 1N NaOH at 60°C, then staining in aceto-iron-hematoxylin for 15-20 h, confirmed a somatic chromosome number of 2n = 16 in all genotypes. This karyotype consisted of: a pair with the longest satellited submetacentric chromosome, a pair with the shortest metacentric chromosome and six pairs of metacentric to submetacentric chromosomes. Intraspecific variabilities were observed for haploid chromosome length, primary constriction position and karyotypic formula. But all of the studied genotypes showed the same karyotype asymmetry, 1B. Haploid genome length ranged from 24.88 μm±0.50 in G6 (Bieanij cv) to 18.48±0.60 μm in G10 (Flip93-93c cv). Cluster analysis classified the 11 genotypes in three groups, having different sizes of chromosomes and arms.
PDF Abstract XML References Citation
How to cite this article
F. Mahmoodi Kordi, A. Majd, M. Valizadeh, M. Sheidai and H. Sabaghpour, 2006. A Comparative Study of Chromosome Morphology among Some Genotypes of Cicer arietinum L. Pakistan Journal of Biological Sciences, 9: 1225-1230.
DOI: 10.3923/pjbs.2006.1225.1230
URL: https://scialert.net/abstract/?doi=pjbs.2006.1225.1230
DOI: 10.3923/pjbs.2006.1225.1230
URL: https://scialert.net/abstract/?doi=pjbs.2006.1225.1230
INTRODUCTION
The genus Cicer (family Leguminosae, subfamily Papilionaceae) which was traditionally placed in the tribe Vicieae, has now been removed and placed in its own monogeneric tribe Cicereae. The genus consists of 43 species, of which nine are annual, 33 perennial and one with as yet an unspecified life cycle (Van der Maesen, 1987).
The only commonly cultivated species, Cicer arietinum L. (chickpea) is an important grain legume in several countries including: Algeria, Ethiopia, India, Iran, Mexico, Morocco, Myanmar, Pakistan, Spain, Syria, Tanzania, Tunisia and Turkey (Upadhyaya et al., 2002). A perennial species, Cicer songoricum steph ex DC is also cultivated to a limited extent as a food plant in Afghanistan, western Himalayas and Tibet (Ohri, 1999). All annual species of Cicer are self-pollinating diploids with 2n = 2x = 16 chromosomes (Van der Maesen, 1987). Many cytological studies have been carried out on Cicer (Mercy et al., 1974; Lavania and Lavania, 1982; Sharma and Gupta, 1986; Venora et al., 1991, 1995a, b; Galasso and Pignone, 1992; Ocampo et al., 1992; Ahmad and Hymowitz, 1993; Tayyar et al., 1994) but a comparative study of chromosome morphology among genotypes in Cicer arietinum (kabuli and Desi types) is not well documented. Information about the relationships among cultivars or genotypes and the genetic diversity in available germplasm is important for the optimal design of chickpea breeding programs.
The main objective of this research was to compare chromosome polymorphism at intraspecific level of Cicer arietinum.
MATERIALS AND METHODS
Eleven chickpea genotypes including nine Kabuli type (Arman; Flip93-93c; Grist; Bieanij; Hashem; Shiraz; Jam; ILC-482) and two Desi type (Kaka; Pirouz) were used in this study (Table 1). The Kabuli types have owl-shaped, large cream coloured seeds, whereas the Desi types have angular shaped small and dark colored seeds (IBPGR, ICRISAT and ICARDA, 1993). The seeds of the genotypes were obtained from Dryland Agricultural Research Institute of Kermanshah-Iran. These genotypes are commercially grown in Iran, where about 650,000 ha are under cultivation of chickpea, as the most important pulse crop (Ministry of Agriculture, 2003).
| Table 1: | List and other features studied genotypes of Cicer arietinum L. for karyotype analysis |
 | |
The study was conducted at laboratory of cytogenetic of Agricultural of Faculty of Tabriz University, Iran, 2004. Seeds were germinated on moist filter paper in petri dishes, kept at room temperature (20-25°C) in the dark. Actively growing roots of about 1-1.5 cm in length were excised, pretreated with a 0.002 M aqueous solution of 8-hydroxyquinoline for 4 h at 4°C, washed in distilled water for 10-15 min and fixed in Lewitsky fixative at 4°C for 30-36 h. The fixative was prepared by mixing equal parts (in volume) of 1% chromic acid and 4% formaldehyde (10% formalin) just before using (Agayev, 2002). The root tips were washed and hydrolized in NaOH 1N for 10 min at 60°C and stained with aceto-iron-hematoxylin 4% for 18 h at 30°C. It was important to drain excess water before immersing roots into the stain (5-6 roots/2.5 mL stain). Little porcelain crucibles (2.5-3 mL) were used for staining. Stained roots were washed in distilled water for at least 30 min, then 1-1.2 mm the root of tips were cut and macerated in cytase for at least 0.5 h. Each of the root tips was carefully transferred on a drop of 45% acetic acid on a slide for 3-5 min, covered by cover slip and gently squashed. A number of 7-10 cells with well spread chromosomes were studied and photographed. All measurements i.e., the haploid genome length (equal to the half of the total length of the chromosome complement) and the proportion of the total chromosome length occupied by satellite were taken from 7-10 cell with similar levels of chromosome condensation. Measurements of the total length of each chromosome and the length of the long and short arms as well as satellites were recorded and measured with Micro Measure 3.3 software (Reeves, 2001).
The chromosome were designated according to Kobayashi and Iwatsubo (1989), reported for Seasanum species and Ahmad (2000), reported for Cicer species. Karyological data were obtained over the eight most definitive cells in each genotype. Statistical analyses, including standard errors for means were performed at 5% probability level by using SPSS software. Karyotype asymmetry was calculated according to Stebbins (1971).
RESULTS
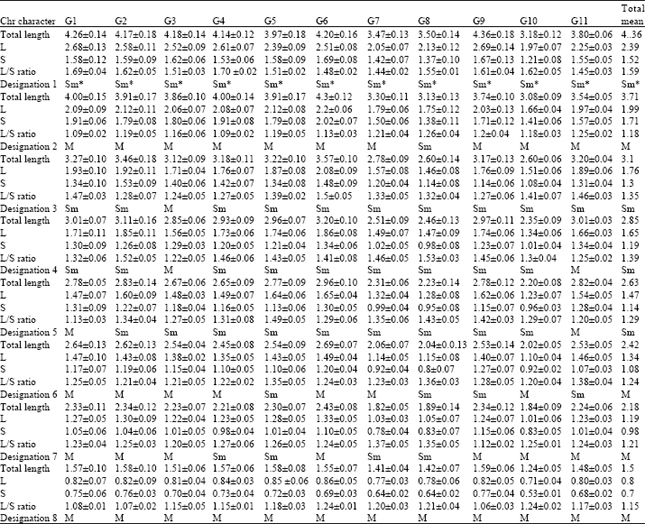
All of the 11 Cicer arietinum genotypes showed a diploid somatic chromosome number of 2n = 16. The designation of some chromosomes in these genotypes presented some difficulties, due to their small size, which affected both the clear discrimination of the arm ratios and the total chromosome length. Average chromosomes lengths, size of long and short arms, L/S ratios are presented in Table 2. Detailed karyotypic complement formula and karyotypic measurement are available in Table 3. A typic karyogram for 11 genotypes of chickpea in Fig. 1 shows that there are a pair of the largest and satellited chromosomes (number 1) submetacentric, a pair of the shortest metacentric chromosomes (number 8) and six pairs of metacentric to submetacentric chromosomes. The distinct characteristics of each chromosome are described as follows:
Chromosome 1: The largest chromosome and its total length ranged from 4.36±0.18 μm in G9 to 3.18±0.12 μm in G10, indicating significant differences among genotypes. All of the genotypes clearly depicted a submetacentric primary constriction. This chromosome also carried a satellite on secondary constriction in the long arm in all genotypes, with apparently another satellite on that in some genotypes.
Chromosome 2: The exception of being submetacentric in G8, this chromosome was metacentric in other genotypes. It’s size is ranged from 4.3±0.12 μm in G6 to 3.08±0.09 μm in G10, with a total mean of 3.71 μm and total arm ratio of 1.18 μm.
Chromosome 3: The longest in G2 and the shortest in G8, being 3.46±0.18 μm and 2.60±0.14 μm, respectively. This chromosome which was submetacentric in all of the genotypes, with the exception of G3 which showed a median centromere.
Chromosome 4: A submetacentric chromosome, except in G3 and G11 with total length of 2.85 μm and arm ratio of 1.39 μm. As in chromosome 2, G6 continued to reveal the largest chr. 4 (3.2±0.10 μm), while G10 had the shortest chr. 4 (2.35±0.09 μm) among the 11 genotypes. Metacentric designation in G3 and G11 can be attributed to the elongation of their short arms.
Chromosome 5: The size of this chromosome was not significantly different from chromosome 4 in any genotype.
| Table 2: | Somatic chromosome (chr) characteristics (size, ratio and designation) of the eleven genotypes of Cicer arietinum. Standard Errors (SE) are given for means. M and Sm refer to metacentric, submetacentric chromosomes. G1-G11 are chickpea genotypes (Table 1) |
 | |
| L = Long arm, S = Short arm | |
| Table 3: | Karyotypic complement measurements and features of Cicer arietinum L. genotypes |
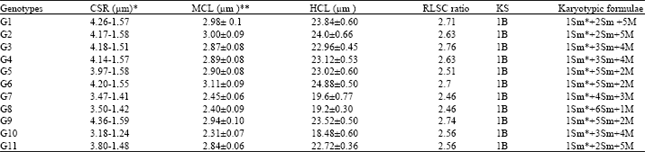 | |
*Chromosome size range in each genotype, **Mean chromosome length±standard error of the genotype, HCL, mean haploid chromosome complement length±standard deviation, RLSC, ratio of the longest to shortest chromosome in the genotype, KS, Karyotype asymmetry according to Stebbins (1971). Materials and Methods for genotypes number | |
The size of this chromosome is ranged from 2.96±0.10 μm in G6 to 2.2±0.08 μm in G10. Chromosome 5 was submetacentric in all of genotypes, except in G1 and G11, in which, similar to chromosome 4,elongation of short arms was observed.
Chromosome 6: A rather small chromosome which size is ranged from 2.69±0.07 μm in G6 to 2.02±0.05 μm in G10. This chromosome was submetacentric in G5, G8, G9 and G11 and metacentric in other genotypes.
 | |
| Fig. 1: | Karyotype of the eleven Genotypes of Cicer arietinum L. (A) Kaka, (B) ILC-482, (C) Pirouz, (D) Bieanij, (E) Ardebil, (F) Hashem, (G) Flip 93-93c, (H) Grist, (I) Jam, (J) Shiraz, (K) Arman. Scale: 3 μm |
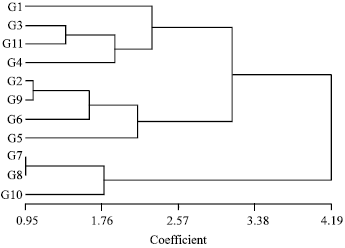 | |
| Fig. 2: | Cluster analysis of 11 genotypes, based on total, long arm, short arm lengths and arm ratio by using UPGMA and Euclidean distances |
It seems that in genotypes having metacentric chromosome, the length of short arm was augmented in relation to total mean, but in those having submetacentric designation, the long arm was increased.
Chromosome 7: This chromosome is not significantly different in size from chromosome 6 in any of the eleven genotypes, with total length of 2.18 μm and total arm ratio of 1.21. Seven genotypes showed metacentric and 4 submetacentric designation.
Chromosome 8: This chromosome is the smallest chromosome in all of the genotypes, which size is ranged from 1.59±0.06 μm in G9 to 1.24±0.05 μm in G10. Chromosome 8 was metacentric in all of the genotypes.
All of the genotypes showed the same karyotype asymmetry of 1B (Table 3), indicating that the Cicer arietinum L. can be considered as a relatively primitive species. However, some polymorphisms was observed among 11 genotypes for karyotype formula.
The genotypes could be generally classified into three karyotypic groups based on individual chromosomes lengths and arm ratios (Fig. 2). It is clear from the present study that two Desi types G7, G8 and G10 formed the first group, all having the smallest haploid chromosome complement (Table 3). While four kabuli types G2, G9, G6 and G5 formed a second cluster, having the largest short arms. Finally; G1, G3, G11 and G4 formed a third cluster, having the largest long arms.
DISCUSSION
The karyotype of Cicer arietinum, derived from 11 genotypes in the present investigation, generally agrees with the previously reported karyotypes (Venora, 1997; Ohri and Pal, 1991; Galasso et al., 1996; Ahmad, 2000). Total length of the chromosomes significantly differed among chickpea genotypes, as observed in some previous studies (Gupta and Sharma, 1991). However, haploid genome lengths observed in this study were generally longer than (about 22.3 μm) those reported earlier (Tayyar et al., 1994; Ahmad, 2000). The discrepancies could be due to a variety of reasons, including variation in the degree of chromosome contraction, methods of measurement or use of different genotypes. Karyotype formulae obtained in present study (1Sm*+3Sm+4M) was also different compared with Ahmad (2000) report, who found (1Sm*+2 Sm+5M). This can be attributed to different genotypes used and different chromosome designations (Levan et al., 1964; Kobayashi and Iwatsubo, 1989). Differences also were observed among genotypes, in the position of primary constrictions and in lengths of the individual chromosomes. Similar results were found by Phadnis (1970), Kutarekar and Wanjari (1983) and Sharma (1983). The submetacentric chromosome 1 was the only pair that was consistently satellited. This is in agreement with the previous mitotic studies (Mercy et al., 1974; Lavania and Lavania, 1982; Ocampo et al., 1992; Tayyar et al., 1994). However, the results were different from the more precise pachytene analysis described by Ahmad and Hymowitz (1993) who found chromosomes but not chromosome 1, to be satellited and associated with the nucleolus.
Additionally, some Cicer arietinum genotypes showed tandem satellited on chromosome 1 during late prophase/early metaphase, which was no longer evident as mitotic cells progressed into metaphase proper, apparently due to chromatin condensation. Such tandem satellites in Cicer arietinum have been previously recorded (Meenakshi and Subramaniam, 1960; Ahmad, 1989), although its significance remains as yet unclear (Ahmad, 2000).
Some studies have even recorded certain Cicer species karyotypes with no satellites (Sharma and Gupta, 1986; Ocampo et al., 1992). Stebbines (1971) classified karyotype asymmetry based on four levels of asymmetry in the centromer position and three levels of asymmetry in chromosome length. The same classification system, when applied to the 11 genotypes of Cicer arietinum, indicated that all of them have relatively asymmetric 1B (Table 3). This is consistent by finding of Ahmad (2000). The 1B type of asymmetry is the characteristic of Cicer arietinum genotypes. While, the A type of asymmetry is the characteristic of perennial Cicer species (Sharma and Gupta, 1986; Ohri and Pal, 1991; Ocampo et al., 1992). Symmetrical karyotypes are primitive features in the genus Cicer, since the 1B type include the cultivated species.
Tayyar et al., (1994) and Galasso et al. (1996) reported that there are rearrangements both in heterochromatin and euchromatin during evolution in this genus, possibly in the same manner as occurred in another leguminous genus Lathyrus (Narayan, 1982). Additionally, it is evident that the size variation among different chromosomes in Cicer arietinum is much greater than that recorded in other annual Cicer species, indicating that ample rearrangement of chromatin has occurred among these chromosomes (Ahmad, 2000). This is true about genotypes studied in this research. Variation among HCL (Haploid Chromosome Length) values in 11 genotypes of Cicer and different karyotypic formulae could be attributed to these rearrangements. But further investigations are needed by banding methods to confirm these rearrangements.
Although, with different methods of clustering, different results could be obtained. Converting Euclidean distances to dendrogram by using UPGMA in this study showed reasonably good results. There for, it can be suggested that for possible use of heterosis, it is suitable to organize breeding programs between the genotypes belonging to different groups.
REFERENCES
- Mercy, T., S.K. Kakar and J.B. Chowdhry, 1974. Cytological studies in three species of the genus Cicer. Cytologia, 39: 383-390.
Direct Link - Reeves, A., 2001. MicroMeasure: A new computer program for the collection and analysis of cytogenetic data. Genome, 4: 439-443.
PubMedDirect Link