
Surajit Sinha
Department of Physics, Jadavpur University, Kolkata-700032, India
Moumita Maiti
Department of Physics, Jadavpur University, Kolkata-700032, India
Krishna Chattopadhyay
Department of Chemical Technology, Calcutta University, Kolkata-700009, India
Brajadulal Chattopadhyay
Department of Physics, Jadavpur University, Kolkata-700032, India
Journal of Pharmacology and Toxicology
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 166-180







ABSTRACT
Nicotine plays an important role on reproductive disorders, cardio-vascular diseases, different cancers and destruction of defence system in human. Its effect in protein malnourished condition is very little understood. This investigation was designed to determine the nicotine-induced toxicity (subcutaneously injected at a dose of 2.5 mg kg-1 b. wt. day-1 for 21 days) on different tissues of female albino rats and corresponding ameliorative observations of curcumin (orally supplemented at a dose of 80 mg kg-1 b. wt. day-1 during the treatment) under protein malnourished condition. Nicotine causes more significant (p<0.001) increase of glucose level, depletion protein and alteration of triglycerides and cholesterol levels in serum. Disruption of liver function and alteration of phosphatase enzymes are more significantly (p<0.001) occurred in protein restricted condition due to nicotine treatment. Nicotine also decreases the activity of antioxidant enzymes, increases lipid peroxidation and affects the production of female hormones more significantly (p<0.001) in such condition. Nicotine disrupts the structural integrity of ovary and uterus that may hamper normal pregnancy. Curcumin effectively ameliorates the deleterious effects of nicotine in protein malnourished condition and may be used in the diet as potential blocker of nicotine to protect the health of malnourished female population.
PDF Abstract XML References Citation
Received: January 10, 2012;
Accepted: March 26, 2012;
Published: June 11, 2012
How to cite this article
Surajit Sinha, Moumita Maiti, Krishna Chattopadhyay and Brajadulal Chattopadhyay, 2012. Potential Amelioration of Curcumin Against Nicotine-induced Toxicity of Protein Malnourished Female Rats. Journal of Pharmacology and Toxicology, 7: 166-180.
DOI: 10.3923/jpt.2012.166.180
URL: https://scialert.net/abstract/?doi=jpt.2012.166.180
DOI: 10.3923/jpt.2012.166.180
URL: https://scialert.net/abstract/?doi=jpt.2012.166.180
INTRODUCTION
India ranks third among the most important tobacco producing countries next only to China and U.S.A. and India (NIOH, 2004). Along with the use of common tobacco forms consumed worldwide, such as cigarettes and pipes, the Indian population are also habituated with Chutta Beedi, (locally use hand made smoking elements) and chewing tobacco those are very common amongst them (Rahman et al., 2003). Nicotine is the major constituent of tobacco (Wu et al., 2002). It has also been implicated in pathologies such as delayed wound healing (Sorensen et al., 2010), reproductive disorders (Windham et al., 2005), cardiovascular disease (Adedeji and Etukudo, 2006; Frost-Pineda et al., 2011) lung and oral cancer (Jung et al., 2001) etc. Nicotine disrupts anti oxidant mechanism (Mary et al., 2002) by enhancing Reactive Oxygen Species (ROS) production and thereby decreases antioxidant level causing peroxidative tissue damages (Ramesh et al., 2007; Chattopadhyay and Chattopadhyay, 2008). The chemicals in tobacco smoke alter endocrine function, perhaps at the level of the ovary which in turn effects release of female hormones. This endocrine disruption likely contributes to the reported associations of smoking with adverse reproductive outcomes, including menstrual dysfunction, infertility and earlier menopause (Windham et al., 2005).
Nutritional factors play an important role in the metabolism and pharmacotoxicological activities of various drugs and xenobiotics (Campbell and Hayes, 1976). A dietary reduction in either quality or quantity of protein causes a depression of hepatic microsomal mixed function oxidase activities. According to Boyd (1969) these results have a direct application to the toxicity of drugs and chemical agents in persons whose dietary protein is low. The major nutritional disorders which are widely prevalent in all developing countries, are due to Protein Energy Malnutrition (PEM). This type of malnutrition affects all age groups and more so of poorer segments of the population. However, the repercussions are the strongest in children and women (Krishnaswami, 1987). Low dietary protein possesses a constraint on the biosynthetic activity, disposition and toxicity (Das et al., 1996). Though it is already established that nicotine causes various damage in our body in normal dietary condition but the toxic effects of nicotine particularly in protein restricted dietary situation are still cause of concern.
Turmeric, cultivated in India, is not only used in Indian delicacies but also used extensively in Ayurveda, an ancient Indian medical system of preventive health care (Aggarwal et al., 2007). Curcumin the main colouring component of turmeric possesses antitumeric (Abdel-Latif and Sadek, 1999), antihepatotoxic, antioxidant (Park et al., 2000) and anti-inflammatory effect (Shehzad et al., 2011). Curcumin also appears to be beneficial in preventing diabetes-induced oxidative stress in rats (Hussein and Abu-Zinadah, 2010). Reports say that administration of curcumin significantly lowers the biochemical marker enzymes, inhibits lipid peroxidation (Kalpana et al., 2007) and enhances the antioxidant status (Kalpana and Menon, 2004). Recently, Tarasub et al. (2011) have shown that curcumin partially protects Cd-induced nephrotoxicity. Our study demonstrates that toxicity of nicotine on different tissues is aggravated under protein restriction condition and curcumin shows its potential to nullify those effects significantly under such condition.
MATERIALS AND METHODS
Chemicals: Nicotine hydrogen tartrate and curcumin were purchased from Sigma Chemicals, St. Louis, USA. All other required chemicals were purchased from Genei. Bangalore, India and Spectrochem Pvt. Ltd. India. All the chemicals and reagents used were analytical grade.
Animals and diet: Thirty adult female albino rats of Wistar strain having body weights 120-130 g were procured at a time from the animal housing facility. The animals were acclimatized under standard conditions of temperature and humidity with 12 h light/dark cycles. They were maintained in accordance with the guidelines of the rule of Instructional Animal Ethics Committee of Jadavpur University, Kolkata, India (constituted as per the “Gazette of India “notification part II Sec. 3 (ii) 17 of the Ministry of Environment and Forestry, Government of India, dated 8th September 1998 for the “Prevention to cruelty to animal 1968”). They were housed to polypropylene cages and were fed standard pellet diet (Hindustan Lever Ltd., India) for 1 week and water ad libitum. Animals were divided equally into two groups (15 animals in each). One group was provided with normal protein diet (18% casein, 70% carbohydrate, 7% fat, 4% salt mixture and 1% vitamin mixture) and the other group with protein-restricted diet (5% casein, 83% carbohydrate, 7% fat, 4 % salt mixture and 1% vitamin mixture) according to Hawk et al. (1954). Each group was further divided into three subgroups (Control, Nicotine treated and Nicotine+Curcumin treated) having 5 animals in each. The animals in each group were maintained on their respective dietary regimen from 7 days before the treatment start till the completion of nicotine treatment.
Mode of treatment: After maintaining the animals in their respective dietary regimen for 7 days, nicotine tartrate (Sigma Chemical Company, St. Louis, MO, USA) dissolved in 0.9% saline (w/v) was injected subcutaneously to the rats of experimental (nicotine treated and nicotine+curcumin treated) subgroups at a dose of 2.5 mg kg-1 b. wt. daily-1 for 21 days at 4 p.m. to avoid diurnal variation. The basis of dose selection was according to the previous study (Mandal et al., 2004). The dilution of the drug was prepared in such a way that 0.5 mL of physiological saline contained the required dose of nicotine. Animals of nicotine+curcumin treated subgroups only received curcumin (80 mg kg-1 b.wt. day-1) orally at the same time (Bandyopadhyaya et al., 2008). The animals in control subgroups of each group received subcutaneous injection of 0.5 mL physiological saline only at the same time.
Sample collection: After the last dose of injection received, animals were kept fasting overnight and sacrificed on the following morning by decapitation (Venkataraman et al., 1984). Anesthetics have been reported to alter the hypothalamo-pituitary axis (Brown and Hegde, 1974). Blood was collected in sterilized tubes and serum was separated out after centrifugation and stored at -20°C prior to further analysis. Liver, ovary and uterus were dissected, cleaned properly and stored for further investigations.
Biochemical analysis: Blood glucose was estimated through Contour TS (Bayer Health Care, Bayer Polychem, India Ltd.). Serum protein concentration was determined by Lowry et al. (1951). The lipid components such as Total Cholesterol (TC) (Allain et al., 1974) and triglyceride (Werner et al., 1981) were estimated in serum by using standard kits supplied by Ranbaxy Diagnostic Ltd., Mumbai, India. By the assay of thiobarbituric acid reactive substances (TBARS), serum and liver lipid peroxidation was measured (Chatterjee and Agarwal, 1998). The amount of MDA was calculated by taking the extinction coefficient of MDA to be 1.56x105 M-1 cm-1. Superoxide Dismutase (SOD) (Beauchamp and Fridovich, 1971), catalase (CAT) (Cohen et al., 1970), Glutathione Peroxidase (GPx) (Levander et al., 1983) and reduced Glutathione (GSH) (Davila et al., 1991) content of liver and serum were determined in rats of all the four groups. Aspartate aminotransferase (AST or SGOT) and alanine aminotransferase (ALT or SGPT) were measured by the method used by Reitman and Frankel (1957). The phosphatase activity in plasma was assayed according to the method of Bassey et al. (1946).
Estimation of steroidogonic hormones: Estradiol is biologically the most active form of the naturally produced estrogen. The VIDAS Estradiol II and VIDAS Progesterone are two automated quantitative tests which were used for detection and measurement of the total 17β-estradiol and progesterone respectively on the VIDAS instruments by using ELFA technique (Enzyme Linked Fluorescent Assay) (New and Used Lab Equipment-GMI Inc.).
Histological study: Cleaned tissues of ovary and uterus were fixed by using Bouin’s fluid. After fixation, the tissues were washed several times by different graded alcohol to remove excess fluid and then embedded in paraffin. Using of rotary microtome sliced the embedded tissues. The paraffin sections were then attached on the slide and washed by xylol before staining. The tissue section was then stained by using haematoxylin and eosin staining.
Statistical analysis: Whole experimental set up was repeated twice and data (n = 10) were averaged and given mean±SD. The statistical analysis of the data obtained from control, nicotine and nicotine+cur cumin supplemented groups from both normal diet and protein restricted diet was performed by one way Analysis of Variance (ANOVA) along with all pair wise multiple comparison procedures (Holm-Sidak method) by using Sigma Stat (version 3.2). The significant levels of the observed data were determined at p<0.01 (significant) and p<0.001 (more or highly significant).
RESULTS
Nicotine significantly (p<0.001) increased the blood glucose levels and decreased the serum protein at both dietary conditions (Table 1). Curcumin significantly (p<0.001) ameliorated the observed nicotine induced effects as shown in Table 1.
Both cholesterol and triglyceride of serum were significantly (p<0.001) increased due to nicotine treatment at both dietary conditions (Table 2). It was seen that the effect of nicotine was aggravated in protein-restricted condition. The amelioration of curcumin was more or less similar in both dietary conditions. Among the most sensitive and widely used liver enzymes are AST or SGOT and ALT or SGPT.
| Table 1: | Effect of nicotine and curcumin on blood glucose and serum protein |
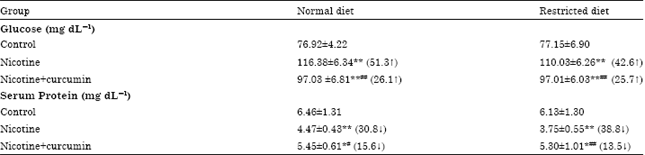 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ##More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 2: | Effect of nicotine and curcumin on serum lipid component |
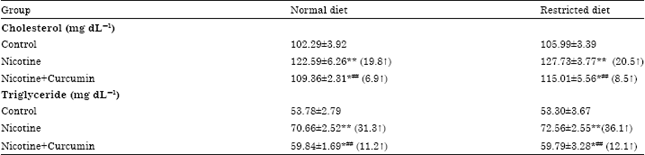 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ##More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 3: | Effect of nicotine and curcumin on liver function enzymes |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 4: | Effect of nicotine and curcumin on superoxide dismutase |
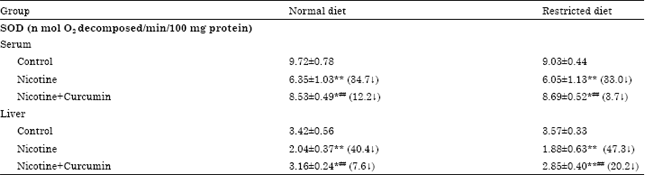 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
These enzymes were increased significantly (p<0.001) in serum of nicotine treated rats at both dietary conditions (Table 3). The increments of ACP were more significant (p<0.001) at protein-restricted condition. Where as higher concentrations of ALP was observed in normal diet fed rat (Table 3). Curcumin supplementation showed better attenuation against nicotine-induced effect on liver enzymes and ACP in protein restriction as reflected by Table 3.
Considerable (p<0.001) decreased SOD activity of serum and liver tissues both in both dietary conditions was seen in nicotine treatment (Table 4). Similar effect of nicotine was also observed in case of other antioxidant enzyme i.e., CAT (Table 5). Curcumin was found to be more active against nicotine-induced effect on antioxidant enzymes particularly in protein restriction condition (Table 4, 5).
| Table 5: | Effect of nicotine and curcumin on catalase |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 6: | Effect of nicotine and curcumin on lipid peroxidation |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 7: | Effect of nicotine and curcumin on reduced glutathione of serum |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
Nicotine more significantly (p<0.001) increased lipid peroxidation in restricted diet fed rats for which higher MDA levels were observed in serum and liver tissues of the rats (Table 6). Again amelioration of curcumin was more significantly (p<0.001) noted in both dietary conditions.
More significant (p<0.001) decreased of glutathione (Table 7) and significant (p<0.01) decreased of GPx (Table 8) in serum were seen due to nicotine treatment on rats of both dietary groups. Curcumin increased the glutathione and GPx levels as seen from Table 7 and 8.
Nicotine caused more significant (p<0.001) reduction of female sex hormones (17β-estradiol and progesterone) in protein-restricted condition as seen in Table 9. Curcumin was found to be more protective to 17β-estradiol than progesterone against nicotine-induced effect in both dietary conditions.
| Table 8: | Effect of nicotine and curcumin on glutathione peroxidase of serum |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
| Table 9: | Effect of nicotine and curcumin on steroidogonic hormones |
 | |
| Values are given mean±SD (where n =10), *Significant at p<0.01, **More significant at p<0.001 with respect to control, #Significant at p<0.01, ## More significant at p<0.001 with respect to nicotine treated rats, the value with in the parenthesis are the percentage of increase (↑) or decrease (↓) with respect to the control value | |
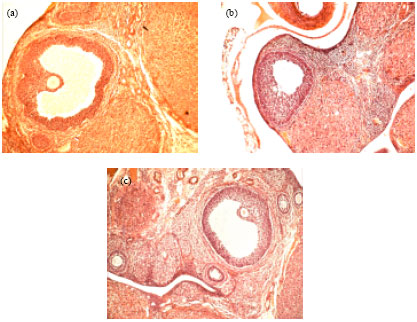 | |
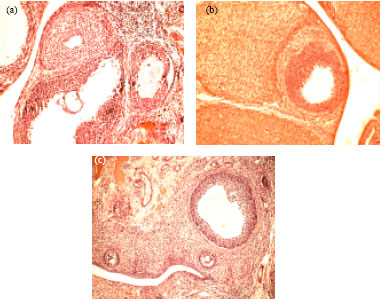
| Fig. 1(a-c): | Histological section of ovary (100x) of rat maintained with normal diet, (a) Control group, (b) Nicotine treated group, (c) Nicotine treated and curcumin supplemented group |
Graafian follicles with an oocyte were observed in the ovarian cross section of both control groups of rats (Fig. 1a and 2a). There was no Graafian follicles were seen in nicotine treated ovarian sections of both groups (Fig. 1b, 2b).
 | |
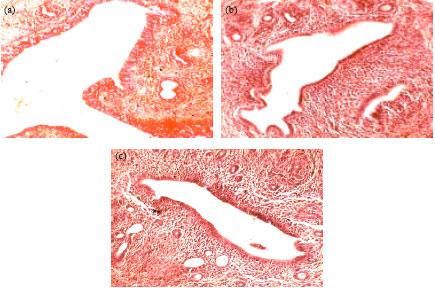
| Fig. 2(a-c): | Histological section of ovary (100x) of rat maintained with protein-restricted diet, (a) Control group, (b) Nicotine treated group, (c) Nicotine treated and curcumin supplemented group |
 | |
| Fig. 3(a-c): | Histological section of uterus (100 x) of rat maintained with normal diet, (a) Control group, (b) Nicotine treated group, (c) Nicotine treated and curcumin supplemented group |
Nicotine inhibits the normal growth of endometrium wall in the uterus as a result of which thinner endometrium wall is observed in the nicotine treated uterus section (Fig. 3b, 4b) than control groups (Fig. 3a, 4a). Curcumin treatment restores the normal structure of ovary (Fig. 1c, 2c) and uterus (Fig. 3c, 4c) to some extent in both dietary conditions.
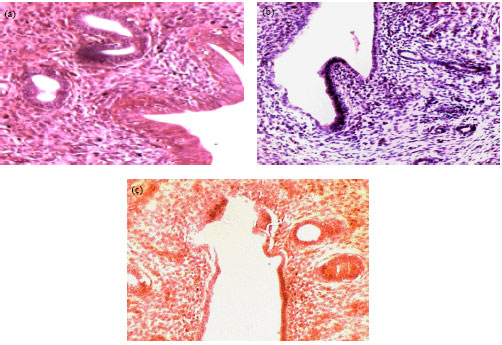 | |
| Fig. 4(a-c): | Histological section of uterus (100 x) of rat maintained with normal diet, (a) Control group, (b) Nicotine treated group, (c) Nicotine treated and curcumin supplemented group |
DISCUSSION
In the present study a significant (p<0.001) elevation of blood glucose level is observed in both groups of animals exposed to nicotine insult (Table 1). Probably this elevation is due to the immediate response to tremendous physiological stress caused by nicotine or by impaired action of insulin. Nicotine induces the increased production of cAMP in blood (Al-Wadei et al., 2009). Increased cAMP in blood stimulates gluconeogenesis and glycogenolysis (Xie et al., 2000) as a result of which the blood glucose level is increased. Fujiwara et al. (2008) have shown that the curcumin possesses anti-diabetic property which is partly due to a reduction in hepatic glucose production caused by activation of AMP kinase and inhibition of G6Pase activity and PEPCK activity. The reduction of blood glucose level by curcumin may be due to its anti-diabetic effect which shows an additive inhibitory effect with insulin on both hepatic gluconeogenesis and glycogenolysis. This result corroborates with the finding of Sivabalan and Anuradha (2010). Nicotine decreases total protein concentration in serum but the decrement is higher in protein-restricted diet (38.8%) than normal protein diet (30.8%) fed rats as seen from Table 1. We have already reported that nicotine aggravates DNA damage under protein-restricted condition (Bandyopadhyaya et al., 2008; Banerjee et al., 2010). The reduced protein concentration in serum may be due to inhibition of protein synthesis caused by DNA damage as well as to low protein content supplied in the diet. The antagonizing effect of curcumin is due to the interaction of curcumin with nicotine as explained earlier (Banerjee et al., 2010).
Cholesterol is an extremely important biological molecule that has roles in membrane structure as well as being a precursor for the synthesis of the steroid hormones and bile acids. Nicotine stimulates catecholamine synthesis which lipolysis adipose tissue resulting an increment in the level of serum cholesterol as well as of triglyceride (Balakrishnan and Menon, 2007). Our findings of significantly (p<0.001) increased levels of serum cholesterol and triglyceride of the nicotine treated rats under both dietary conditions are in agreement with earlier studies (Balakrishnan and Menon, 2007; Chattopadhyay and Chattopadhyay, 2008). In curcumin treated group the level of cholesterol is decreased probably due to the increase in the CYP7A1 gene expression which is a rate limiting enzyme in the biosynthesis of bile acid from cholesterol as suggested by Kim and Kim (2010). Curcumin showed an obvious hypocholesterolemic effect that could be due to an effect on total cholesterol absorption (Hasimun et al., 2011), degradation or elimination (Arafa, 2005). Triglyceride lowering effect is probably due to multiple inductions of fatty acid catabolism (Asai and Miyazawa, 2001).
Increased activities of AST and ALT by nicotine administration (Table 2) indicate liver tissue damage that is occurred due to loss of functional integrity of cells membranes (Balakrishnan and Menon, 2007; Wetscher et al., 1995). These enzymes are normally contained within liver cells. If the liver is injured or damaged, the liver cells spill these enzymes into the blood, raising the enzyme levels in the blood and signaling the liver disease. Curcumin decreases the levels of the liver enzymes by preserving the structural integrity of the cell membrane (Al-Jassabi and Azirun, 2010). Acid phosphatase can free attached phosphate groups from other molecule during digestion. Basically it is a phosphomonoesterase. Nicotine increases the ACP and ALP by oxidative stress providing tissue injury and directly involved in the generation of reactive oxygen species (ROS) as explained by Mohammad and Ahmad (2010). In Curcumin treated group, both ACP and ALP levels are decreased probably due to improve in antioxidant status (Table 3).
Nicotine is a highly addictive alkaloid induced oxidative stress both in vivo and in vitro (Mahapatra et al., 2009). It has been reported that the nicotine disrupts the mitochondrial respiratory chain leading to an increase generations of super oxide ions and hydrogen peroxide. (Yildiz et al., 1998). Superoxide anion and hydrogen peroxide are the main sources of the nicotine induced free radical generation and depletion of the cellular antioxidant (Chakraborty et al., 2010). Glutathione being an important cellular reductant involves in protection against free radicals, peroxides and toxic compounds (Grester, 1995). Therefore depletion of GSH not only impairs cell defense against toxic compounds but also results in enhanced oxidative stress and tissue damage (Sener et al., 2007; Sreekala and Indira, 2009). Our observation shows that nicotine treatment more significantly (p<0.001) depletes GSH level of serum of both dietary groups indicating higher level of tissue damage (Table 7). GPx decomposes excess hydro peroxides including hydrogen peroxide (H2O2). Decreased level of GPx also confirms that nicotine challenges the cell defense mechanism and disrupts antoxidative activities (Table 8). The ameliorative effect of curcumin is due to its scavenging or neutralizing free radicals activities that inhibits peroxidation of membrane lipids and maintains cell membrane interiority and functions (Ritukumari et al., 2002).
Smoking has multiple effects on hormone secretion which is mediated by nicotine Kapoor and Jones (2005). Nicotine induces significant (p<0.001) reduction of 17β-estradiol and progesterone levels in both experimental groups. It is observed that percentage decrease of plasma 17 β-estradiol and progesterone levels during protein restriction are greater than the normal diet fed groups (Table 9). Proteins are the building blocks for enzyme biosynthesis. It may be conjectured that low dietary level of protein causes a decrease in the synthesis of steroidogenic enzyme, resulting in lowering of steroid hormone levels at restricted diet control groups. This is further aggravated by exposure to nicotine. 17β-estradiol level was decreased in nicotine treated group due to an anti estrogenic effect of nicotine which affected in hepatic estrogen metabolism (Michnovicz et al., 1986; Kapoor and Jones, 2005). Smoking has a powerful effect on the 2-hydroxylation pathway of estradiol metabolism leading to increased production of 2-hydroxyestrogens. These compounds have minimal estrogenic activity and are rapidly cleared from the circulation. Nicotine also decreases progesterone level by increasing PGF2α, an important factor for luteolysis and also by increasing VEGF mRNA expression (Miceli et al., 2005). Curcumin reduces the effect in nicotine treated group due to its high estradiol binding capacity (Zava et al., 1998). In Curcumin treated group progesterone level is increased because curcumin inhibits microvascular endothelial cell angiogenesis through inhibition of COX-2 expression (Binion et al., 2008).
By the histological studies, we have seen that there is marked decrease in growth of follicles in ovary under both normal and restricted dietary conditions due nicotine challenge. Graafian follicles with an oocyte are observed in the ovarian cross section in both normal groups of rats. But no graafian follicles are observed in the ovarian cross section when the animals are exposed in nicotine. This is probably due to the decrement of estrogen level in nicotine treatment. Growth of follicles are increased as well as oocyte appeared with normal form in the ovary after curcumin supplementation (Fig. 1) which indicates that curcumin restores the normal ovarian function by minimizing the toxic effect of nicotine which in turns increases estrogen levels (Table 9). According to Soares et al. (2007), heavy smoking disrupts the stability of cells in the lining of uterus differently which overall reduces general pregnancy rate. He also reports that there is a change of endometrium in heavy smokers. In our finding it is noted that endometrium of the uterus of nicotine treated rats becomes thinner than that of control groups of both dietary conditions (Fig. 2). Curcumin regains the normal structure and architecture of endometrium as a result of which the endometrium becomes thick. Many scientists have already shown that curcumin has a broad range of biological activities which are beneficial for our health (Foda et al., 2007; Srivastava et al., 2011). Our results are in line with the observations reported earlier.
CONCLUSION
Low dietary protein induces a stress in the body. Stress and nicotine therefore synergistically heightens the immunological disorder in protein-restricted condition. Curcumin effectively antagonizes the deleterious effects of nicotine and be used in the diet as potential blocker of nicotine to protect the female population at malnourished condition.
ACKNOWLEDGMENTS
Authors are gratefully acknowledged the technical support of Biophysics Laboratory under Physics Department of Jadavpur University. The technical help from Mr. Tapan Kumar Mukherjee, Director, Immuno Diagnostics Centre, Netaji Nagar, Kolkata, India is gratefully acknowledged.
REFERENCES
- Abdel-Latif, H. and I.A. Sadek, 1999. Curcumin and its effect on cytochrome P450 and GST in toad liver tumor induced by DMBA. Pak. J. Biol. Sci., 2: 1193-1196.
CrossRefDirect Link - Adedeji, O.A. and M.H. Etukudo, 2006. Lipid profile of cigarette smokers in calabar municipality. Pak. J. Nutr., 5: 237-238.
CrossRefDirect Link - Aggarwal, B.B., C. Sundaram, N. Malani and H. Ichikawa, 2007. Curcumin: The Indian solid gold. Adv. Exp. Med. Biol., 595: 1-75.
CrossRefDirect Link - Allain, C.C., L.S. Poon, C.S.G. Chan, W. Richmond and P.C. Fu, 1974. Enzymatic determination of total serum cholesterol. Clin. Chem., 20: 470-475.
CrossRefPubMedDirect Link - Al-Wadei, H.A., H.K. 3rd Plummer and H.M. Schuller, 2009. Nicotine stimulates pancreatic cancer xenografts by systemic increase in stress neurotransmitters and suppression of the inhibitory neurotransmitter γ-aminobutyric acid. Carcinogenesis, 30: 506-511.
PubMedDirect Link - Arafa, H.M., 2005. Curcumin attenuates diet induced hypercholesterolemia in rats. Med. Sci. Monit., 11: 228-234.
Direct Link - Balakrishnan, A. and V.P. Menon, 2007. Protective effect of hesperidin on nicotine induced toxicity in rats. Indian J. Exp. Biol., 45: 194-202.
PubMedDirect Link - Bandyopadhyaya, G., S. Sinha, B.D. Chattopadhyay and A. Chakraborty, 2008. Protective role of curcumin against nicotine induced genotoxicity on rat liver under restricted dietary protein. Eur. J. Pharmacol., 588: 151-157.
CrossRef - Banerjee, S., G. Bandyopadhyaya, K. Chattopadhyay and B.D. Chattopadhyay, 2010. Amelioration of nicotine-induced damage of blood cells in protein malnourished female rats by curcumin. Int. J. Pharmacol., 6: 444-455.
CrossRefDirect Link - Binion, D.G., M.F. Otterson and P. Rafiee, 2008. Curcumin inhibits VEGF-mediated angiogenesis in human intestinal microvascular endothelial cells through COX-2 and MAPK inhibition. Gut, 57: 1509-1517.
CrossRefDirect Link - Brown, M.R. and G.A. Hegde, 1974. In vivo effects of postaglandins on TRH-induced TSH secretion. Endocrinology, 95: 1392-1397.
PubMedDirect Link - Campbell, T.C. and J.R. Hayes, 1976. The effect of quantity and quality of dietary protein in drug metabolism. Fed. Proc., 35: 2470-2474.
PubMedDirect Link - Chakraborty, S.P., S.K. Mahapatra, S.K. Sahu, P. Pramanik and S. Roy, 2010. Antioxidative effect of folate modified chitosan nanaparticals. Asian Pac. J. Trop. Biomed., 1: 29-38.
Direct Link - Chatterjee, S.N. and S. Agarwal, 1998. Liposomes as membrane model for study of lipid peroxidation. Free Radic. Biol. Med., 4: 51-72.
PubMedDirect Link - Chattopadhyay, K. and B.D. Chattopadhyay, 2008. Effect of nicotine on lipid profile, peroxidation and antioxidant enzymes in female rats with restricted dietary protein. Indian J. Med. Res., 127: 571-576.
PubMedDirect Link - Cohen, G., D. Dembiec and J. Marcus, 1970. Measurement of catalase activity in tissue extracts. Ann. Biochem., 34: 30-38.
CrossRefPubMedDirect Link - Davila, J.C., P.L. Davis and D. Acosta, 1991. Changes in glutathione and cellular energy as potential mechanisms of papaverine induced hepatotoxicity in vitro. Toxicol. Applied Pharmacol., 108: 28-36.
Direct Link - Foda, M.I., M. Abd El-Aziz and A.A. Awad, 2007. Chemical, rheological and sensory evaluation of yoghurt supplemented with turmeric. Int. J. Dairy Sci., 2: 252-259.
CrossRefDirect Link - Frost-Pineda, K., Q. Liang, J. Liu, L. Rimmer and Y. Jin et al., 2011. Biomarkers of potential harm among adult smokers and nonsmokers in the total exposure study. Nicotine Tob. Res., 10: 182-193.
PubMedDirect Link - Fujiwara, H., M. Hosokawa, X. Zhou, S. Fujimoto and K. Fukuda et al., 2008. Curcumin inhibits glucose production in isolated mice hepatocytes. Diabetes Res. Clin. Pract., 80: 185-191.
PubMed - Windham, G.C., P. Mitchell, M. Anderson and B.L. Lasley, 2005. Cigarette smoking and effects on hormone function in premenopausal women. Environ. Health Perspec., 113: 1285-1290.
CrossRefPubMedDirect Link - Grester, H., 1995. β-Carotine, vitamin E and vitamin C in different stages of carcinogenesis. Eur. J. Clin. Nutr., 149: 155-168.
Direct Link - Hasimun, P., E.Y. Sukandar, I.K. Adnyana and D.H. Tjahjono, 2011. Synergistic effect of curcuminoid and s-methyl cysteine in regulation of cholesterol homeostasis. Int. J. Pharmacol., 7: 268-272.
Direct Link - Hussein, H.K. and O.A. Abu-Zinadah, 2010. Antioxidant effect of curcumin extracts in induced diabetic Wister rats. Int. J. Zool. Res., 6: 266-276.
CrossRefDirect Link - Jung, C., Y. Rong, S. Doctrow, M. Baudry, B. Malfroy and Z. Xu, 2001. Synthetic superoxide dismutase/catalase mimetics reduce oxidative stress and prolong survival in a mouse amyotrophic lateral sclerosis model. Neurosci. Lett., 304: 157-160.
PubMedDirect Link - Kalpana, C., A.R. Sudheer, K.N. Rajasekharan and V.P. Menon, 2007. Comparative effects of curcumin and its synthetic analogue on tissue lipid peroxidation and antioxidant status during nicotine-induced toxicity. Singapore Med. J., 48: 124-130.
PubMedDirect Link - Kapoor, D. and T.H. Jones, 2005. Smoking and hormones in health and endocrine disorders. Eur. J. Endocrinol., 152: 491-499.
PubMedDirect Link - Mahapatra, S.K., S. Das, S. Bhattacharjee, N. Gautam, S. Majumdar and S. Roy, 2009. In vitro nicotine-induced oxidative stress in mice peritonial macrophages: A dose-dependant approach. Toxicol. Mech. Methods, 19: 100-108.
PubMedDirect Link - Kim, M. and Y. Kim, 2010. Hypocholesterolemic effects of curcumin via up-regulation of cholesterol 7a-hydroxylase in rats fed a high fat diet. Nutr. Res. Pract., 4: 191-195.
CrossRefDirect Link - Krishnaswami, K., 1987. Drug/Xenobiotic-metabolism, disposition and toxicity in malnutrition. Def. Sci. J., 37: 133-142.
Direct Link - Levander, O.A., D.P. Deloach, V.C. Morris and P.B. Moser, 1983. Platelet glutathione peroxidase activity as an index of selenium status in rats. J. Nutr., 113: 55-63.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mandal, K., S. Dasgupta and B.D. Chattopadhyay, 2004. Nicotine induced alterations on reproductive organs of female rats under protein deficient condition. Indian J. Exp. Biol., 42: 330-332.
PubMedDirect Link - Mary, B.N., W.A. Gary, S.R. Douglas, C.B. Paula, T. Thomas and R.S. Paul, 2002. Nicotine oxidative and antioxidant properties in CNS. Life Sci., 71: 2807-2820.
Direct Link - Miceli, F., F. Minici, A. Tropea, S. Catino and M. Orlando et al., 2005. Effects of nicotine on human lutea cells in vitro: A possible role on reproductive outcome for smoking women. Biol. Reprod., 72: 628-632.
PubMedDirect Link - Michnovicz, J.J., R.J. Hershcopf, H. Naganuma, H.L. Bradlow and J. Fishman, 1986. Increased 2-hydroxylation of estradiol as a possible mechanism for the anti-estrogenic effect of cigarette smoking. N. Eng. J. Med., 315: 1305-1309.
PubMedDirect Link - Park, E.J., C.H. Jeon, G. Ko, J. Kim and D.H. Sohn, 2000. Protective effect of curcumin in rat liver injury induced by carbon tetrachloride. J. Pharm. Pharmacol., 52: 437-440.
CrossRefDirect Link - Ramesh, T., R. Mahesh and V. Hazeena Begum, 2007. Effect of Sesbania grandiflora on lung antioxidant defense system in cigarette smoke exposed rats. Int. J. Biol. Chem., 1: 141-148.
CrossRef - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Sener, G., H.Z. Toklu and S. Cetinel, 2007. β-Glucan protects against chronic nicotine-induced oxidative damage in rat kidney and bladder. Environ. Toxicol. Pharmacol., 23: 25-32.
Direct Link - Shehzad, A., T. Ha, F. Subhan and Y.S. Lee, 2011. New mechanisms and the anti-inflammatory role of curcumin in obesity and obesity-related metabolic diseases. Eur. J. Nutr., 50: 151-161.
CrossRefPubMedDirect Link - Sivabalan, S. and C.V. Anuradha, 2010. A comparative study on the antioxidant and glucose-lowering effects of curcumin and bisdemethoxycurcumin analog through in vitro assays. Int. J. Pharmacol., 6: 664-669.
CrossRefDirect Link - Sorensen, L.T., B.G. Toft, J. Rygaard, S. Ladelund and M. Paddon et al., 2010. Effect of smoking, smoking cessation and nicotine patch on wound dimension, vitamin C and systemic markers of collagen metabolism. Surgery, 148: 982-990.
CrossRef - Sreekala, S. and M. Indira, 2009. Effects of exogenous selenium on nicotine-induced oxidative stress in rats. Biol. Trace Elem. Res., 130: 62-71.
CrossRefDirect Link - Pandey, A., R.K. Gupta and R. Srivastava, 2011. Curcumin-the yellow magic. Asian J. Applied Sci., 4: 343-354.
CrossRefDirect Link - Venkataraman, B.V., T. Joseph, P.S. Shetty and P.M. Stephen, 1984. Acetylcholine levels of rat brain and heart in starvation and protein restriction. Ind. J. Physiol. Pharmacol., 28: 223-226.
PubMedDirect Link - Werner, M., D.G. Gabrielson and J. Estman, 1981. Ultramicro determination of serum triglycerides by bioluminescent assay. Clin. Chem., 27: 268-271.
PubMed - Wetscher, G.J., D. Bagchi, G. Perdikis, M. Bagchi and E.J. Redmond et al., 1995. In vitro free radical production in rat esophageal mucosa induced by nicotine. Digest. Dis. Sci., 40: 853-858.
CrossRefDirect Link - Wu, Y.P., K. Kita and N. Suzuki, 2002. Involvement of human heat shock protein 90α in nicotine-induced apoptosis. Int. J. Cancer, 100: 37-42.
CrossRefDirect Link - Yildiz, D., N. Ercal and D.W. Armstrong, 1998. Nicotine enantiomers and oxidative stress. Toxicology, 130: 155-165.
CrossRef - Beauchamp, C. and I. Fridovich, 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem., 44: 276-287.
CrossRefPubMedDirect Link