
N.A. Al-Saady
Department of Crop Sciences, College of Agricultural and Marine Sciences, Sultan Qaboos University, P.O. Box-34, Al-Khod 123, Sultanate of Oman
A.J. Khan
Department of Crop Sciences, College of Agricultural and Marine Sciences, Sultan Qaboos University, P.O. Box-34, Al-Khod 123, Sultanate of Oman
L. Rajesh
Department of Crop Sciences, College of Agricultural and Marine Sciences, Sultan Qaboos University, P.O. Box-34, Al-Khod 123, Sultanate of Oman
H.A. Esechie
Department of Crop Sciences, College of Agricultural and Marine Sciences, Sultan Qaboos University, P.O. Box-34, Al-Khod 123, Sultanate of Oman
Journal of Plant Sciences
Year: 2012 | Volume: 7 | Issue: 5 | Page No.: 176-185







ABSTRACT
Fenugreek (Trigonella foenum gracium L.) is a widely known medicinal semi-arid crop, cultivated worldwide. But this herb is susceptible to salt stress especially during the vegetative growth phases. In order to investigate selected physiological characteristics like germination stress, proline and chlorophyll content under salinity stress, 8 Omani fenugreek accessions were exposed to 0, 4, 6, 8, 10 dS m-1 NaCl for 8 days. All accessions showed salt tolerance at germination and seedling growth at low level of salinity. But germination was delayed as NaCl concentration increased and 10 dS m-1 NaCl showed very poor germination rate. The accessions were then transferred to pots under same salt concentration to analyze the proline and chlorophyll content. The presence of NaCl in soil significantly influenced both proline and chlorophyll content and they both correlated positively.
PDF Abstract XML References Citation
Received: October 19, 2012;
Accepted: December 19, 2012;
Published: February 19, 2013
How to cite this article
N.A. Al-Saady, A.J. Khan, L. Rajesh and H.A. Esechie, 2012. Effect of Salt Stress on Germination, Proline Metabolism and Chlorophyll Content of Fenugreek (Trigonella foenum gracium L.). Journal of Plant Sciences, 7: 176-185.
DOI: 10.3923/jps.2012.176.185
URL: https://scialert.net/abstract/?doi=jps.2012.176.185
DOI: 10.3923/jps.2012.176.185
URL: https://scialert.net/abstract/?doi=jps.2012.176.185
INTRODUCTION
Plant growth is affected by a combination of environmental stresses including drought, salinity and temperature causing substantial and unpredictable losses in productivity of valuable crops (Khanna-Chopra and Selote, 2007). Soil salinity causes stress to crop plants at various magnitudes that are predominantly sensitive to the presence of high salt concentration (Dodd and Donovan, 1999). Seed germination is the most critical phase of plant life which can greatly be influenced by salinity. Plant growth and metabolism can be altered by saline stress (Misra and Dwivedi, 1990, 1995, 2004; Misra and Gupta, 2005). Cellular events such as salt compartmentation and osmotic adjustment in salt-tolerant plants allow continued growth in presence of NaCl (Volkmar et al., 1998). Salt ions play an important role in osmotic adjustment, in stressed sugar beet plants (Ghoulam et al., 2002) as they were efficiently compartmentalized at the cell level. Osmotic adjustment being an adaptive mechanism involved in drought or salinity tolerance (Misra and Gupta, 2005) which allows the balance of turgor under conditions of water deficit (Morgan, 1984) commonly proposed mechanism include compartmentation of ions in vacuoles (Yeo, 1998), accumulation of compatible solutes in the cytoplasm (Hare et al., 1998) and also genetic salt resistance. Mineral toxicity and also genetic salt resistance. Mineral toxicity and nutritional imbalance often accompany the accumulation of ions. Production and accumulation of compatible organic solutes in the cytoplasm obtain osmotic adjustment, in addition to several other cellular and molecular mechanisms. The major compatible solutes include proline and glycinebetaine (Misra and Gupta, 2005, Girija et al., 2002) are thought to function as osmoprotectants for protein (Bohnert and Jenson, 1996). These solutes also provide a protective environment for enzymes and macromolecular structure and function. To counteract these stresses, plants accumulate some compatible osmolytes among which proline plays a very important role (Boyer, 1982; Skriver and Mundy, 1990). However plants species differ in their sensitivity and tolerance to salts (Troech and Thompson, 1993).
Germination is one of the most critical periods for a crop subjected to salinity (Fowler, 1991). Several researchers have observed a decrease in germination rate as salinity increase and the osmotic potential of the germination medium decreases (Greenway, 1973; Redmann, 1974; Sharma, 1976).
During growing phases, plant cells accumulate proline as an osmoprotectant to conserve osmotic stability and to prevent damage (Delauney and Verma, 1993; Roosens et al., 1999). The quantity of proline may vary depending on the amount of NaCl in the soil or water which may indeed differ from plant to plant. Plants also show high chlorophyll degradation symptoms like chlorosis, a common morphological and physiological response to salt stress. The elevated proline contents coupled with reduced chlorophyll levels under salt stress condition were reported by Harinasut et al. (2000).
Fenugreek (Trigonella foenum gracium L.) is an annual legume which is extensively cultivated in most regions of the world for its medicinal value (Petropoulos, 2002). In many countries this species is grown in arid and semi-arid regions such as Oman where high salt concentration is an important characteristic of the soil. Local cultivars and landraces have persisted in situ because of the unfavorable climatic factors like sandy soil and saline water in Oman. Reduction in chlorophyll pigments and photosynthesis activity on fenugreek growth under seawater stress was recorded by Haroun (2002). But studies showing whether proline and chlorophyll content correlate during salt stress in fenugreek are least investigated. The objective of the present research was to study the effect of salinity on germination in fenugreek seeds and to find out the correlation between proline and chlorophyll content during salt stress.
MATERIALS AND METHODS
Plant material and salt treatments: The seeds of 8 fenugreek accessions were obtained from the Ministry of Agriculture (MOA) Al-Rumais, Sultanate of Oman. Seeds were washed thoroughly in tap water followed by distilled water as a part of sterilization. Selected 50 seeds of each accession were grown in sterile Petri dishes containing Whatman # 2 filter paper of 9 cm (Whatman, England) as a supporting medium. Four different dilution levels were used which were 4, 6, 8 and 10 dS m-1 NaCl and distilled water as the control, made up a total of 5 treatments. Water and fresh salt solutions were added periodically added to maintaining the filter paper’s moistness. The Petri dishes were kept in growth chamber for 8 days at day/night temperature of 27°/23°C. The seeds were considered germinated when the plumule and radicle protruded to a length of 3 mm. The number of germinated seeds was counted every other day starting two days from the beginning of the test.
The Germination Stress Index (GSI) was calculated using the equation;
Promotion index (PI) = nd2 (1.00)+nd4 (0.75)+nd6 (0.50)+nd8 (0.25) |
where, nd2, nd4, nd6, nd8 are percentages of seeds germinated on the 2nd, 4th, 6th and 8th day, respectively.
Germinated seeds were further subjected to the same salinity level treatments to evaluate the effect of salinity in growing stages of seedlings. Seedlings of 8 days old were selected from all accessions and were transplanted in sterile plastic pots (16 cm diameter) contains sterile sand. Before applying salinity treatment a period of one week was given to the seedling as a period of pre-conditioning. During this period the seedlings were supplies with Hoagland’s solution. Then the salinity treatments were imposed on alternative weeks for a period of 6 weeks, during which the seedlings also received Hoagland’s solution. All morphological changes were observed during plant growth.
Determination of leaf proline content: The free proline content of both control and treated plants were determined by the method of Bates et al. (1973). Youngest fully expanded leaves of 0.5 g fresh weight from all treatments were harvested and homogenized in 10 mL 3% aqueous sulfosalicylic acid and the homogenate was filtered through Whatman # 1 filter paper of 110 mm diameter. Two milliliter of the filtrate was then reacted with equal volume of freshly prepared ninhydrin reagent and glacial acetic acid and incubated for 1 h at 100°C in boiling water bath. The reaction was then terminated immediately by placing the test tubes on ice bath. Four ml toluene was added into each test tube and shaken vigorously for 10 sec. The pink coloured chromophore was then carefully separated and measured for proline quantification at 520 nm with a Lambda EZ 150 UV/Vis spectrophotometer (Perkin Elmer, USA). A standard curve made from L-proline was used to determine proline concentration (μmol of L-proline) of samples.
Measurement of leaf chlorophyll: Leaf chlorophyll content of fenugreek plants were measured using Chlorophyll meter (Minolta SPAD-502, Japan). The second and third leaves from shoot apex were measured and averages were taken.
Correlation between proline and chlorophyll content: Correlation between Proline and Chlorophyll contents were calculated with the help of a line graph. A trend line was created using the linear equation:
y = ax+b |
where, y represents the proline content (μmolg-1 fresh weight), x represents the chlorophyll quantity (SPAD), a and b are constants. R-squared value (R2) was calculated to represent the proportion of total variability of the ‘y’ values that is accounted for by the independent variable ‘x’.
RESULTS AND DISCUSSION
Effect of salinity on fenugreek seed germination: In present study, the effect of salinity on germination of fenugreek seeds has been investigated. Increase in NaCl concentration in the germination medium significantly influenced germination. The percentage of germination decreased as the salt concentration increased as shown in Table 1. The results indicate that almost 85% accessions germinated satisfactorily because their Germination Stress Index (G.S.I) was more than 80 at lower salinity levels (4 dS m-1). These results are in agreement with previous findings (Kafi and Goldani, 2001; Ashraf and Waheed, 1993; Sekeroglu et al., 1999).
| Table 1: | Germination stress index of 8 genotypes of fenugreek in response to four levels of salinity |
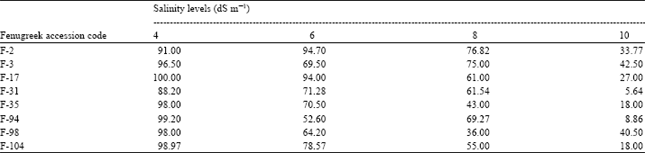 | |
When salt concentration was raised to 6 dS m-1, all accessions exhibited reduction in G.S.I expect in one cultivar (F2) which showed G.S.I value greater than in 4 dS m-1 indicating tolerance to salinity up to certain levels compared to other accessions. At 8 dS m-1, only 30% of the accessions germinated which was even less in 10 dS m-1. Todd (2001) reported similar results in beach peas reporting that delay in germination by increased salt concentration may be explained by the lower osmotic potential of the solution. Katembe et al. (1998) explained that increase in salinity lead to a reduction and/or delay in germination of both halophyte and glycophyte seeds.
Effect of salinity on leaf proline content: Proline accumulation in salt stressed plants is a primary defense response to maintain the osmotic pressure in a cell which is reported in salt tolerant and salt sensitive cultivars of many crops (De Lacerda et al., 2003, 2005; Demiral and Turkan, 2005; Mansour et al., 2005; Misra and Gupta, 2005; Desingh and Kanagaraj, 2007; Koca et al., 2007; Veeranagamallaiah et al., 2007).
In present study, the level of free proline in fenugreek plants were measured during a period of 8 days. Fig. 1 and 2 reveals that under non-stress conditions, the levels of proline were low and relatively similar. But the level of proline started increasing as the concentration of salt increased and plants grown in 8 dS m-1 of sodium chloride showed the highest proline content. Dracup (1991) mentioned that proline levels increase with increasing salt concentrations. After reaching a maximum, the level of proline decreased but remained at a medium value after 8 day. This is consistent with the experiments of Lutts et al. (1999) in rice cultivars that are sensitive to salt stress. In these experiments, proline content in leaves increased in plants exposed to 50 mM of sodium chloride and decreased in plants exposed to 100 mM. Similar results were obtained in experiments made with Sapindus trifoliatus L. somatic embryo using different sodium chloride concentrations (Unnikrishnan et al., 1991). In our experiment proline content decrease after 8 dS m-1 of NaCl have been due to greater proline utilization than synthesis caused by NaCl stress.
Increased proline accumulation under NaCl stress may be due to breakdown of proline-rich protein or de novo synthesis of proline (Tewari and Singh, 1991). It may also be due to prevention of feedback inhibition of the biosynthetic enzyme caused by sequestering proline away from its site of synthesis or by relaxed feedback inhibition of the regulatory step or by decreased activity of proline oxidase involved in degradation of proline. Therefore, accumulation of proline oxidation or diminished incorporation of proline into protein due to impaired protein synthesis and reduced growth. Proline accumulation under salt stress can be explained by the higher inhibitory rate of proline dehydrogenase and proline oxidase (Misra and Gupta, 2006).
Metabolic stress caused by sodium chloride may result in decreased plant growth (Misra and Dwivedi, 2004). Cellular events such as compartmentation and osmotic adjustment in salt tolerant plants may allow continued growth in presence of NaCl (Misra and Gupta, 2005; Volkmar et al., 1998).
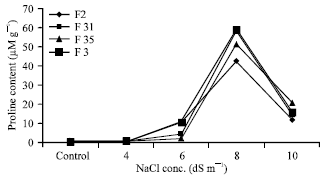 | |
| Fig. 1: | Proline accumulation in 4 accessions of fenugreek after exposure to 4 different NaCl dilutions over a period of 8 days, Values are the mean of five independent determinations |
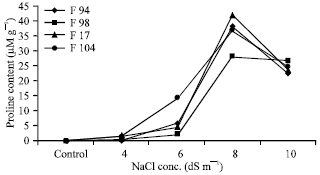 | |
| Fig. 2: | Proline accumulation in 4 accessions of fenugreek after exposure to 4 different NaCl dilutions over a period of 8 days, Values are the mean of five independent determinations |
Proline may also act as an enzyme protectant stabilizing the structure of macromolecules and organelles. Generally proline accumulation appears to be a plant response associated with salt tolerance not a reaction to salt stress damage (Misra and Gupta, 2005; De Lacerda et al., 2003). It has been reported that proline accumulation was not related to salt tolerance or to salt stress acclimation, but probably is just on expression of the seedling reaction to the stress damaging (De Lacerda et al., 2003).
Effect of salinity on leaf chlorophyll content: NaCl caused a marked decrease in the chlorophyll pigment content of fenugreek leaf. Chlorophyll contents in plants exposed to 4 dS m-1 and 6 dS m-1 levels of sodium chloride were similar to or sometimes greater to those observed in controls. Chlorophyll content fell significantly at higher sodium chloride concentration (8 dS m-1, 10 dS m-1) after 15 days (Fig. 3). Similar results were reported with rice and mungbean plants exposed to different concentration of salt (Singh et al., 1973; Garg et al., 1996; Zayed and Zeid, 1997; Asch et al., 1999; Wahid et al., 2004). Plants showed necrosis at 30 days and thus chlorophyll content was affected. This reduction in pigment contents can be due to the inhibitory effect of the accumulated ions (Na and Cl) on the biosynthesis of the different pigment fractions and/or/on their degradation or due to the effect of NaCl on chloroplast structure.
Correlation between Proline and Chlorophyll Content: NaCl concentration 8 dS m-1 affected both proline and chlorophyll content in fenugreek plants. The results indicated that high proline content is 8 dS m-1 positively correlates with the increase of chlorophyll pigment content (Fig. 6).
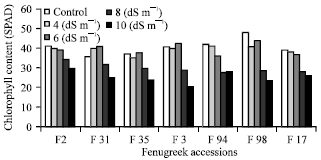 | |
| Fig. 3: | Chlorophyll content of 8 fenugreek accessions in different NaCl dilutions, Values are means of 5 replicates |
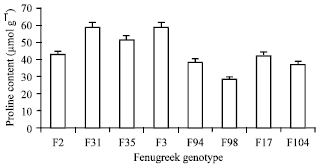 | |
| Fig. 4: | Proline accumulation in 8 accessions of fenugreek after exposure to 8 dS m-1 NaCl for 8 days, Values are Mean±SE |
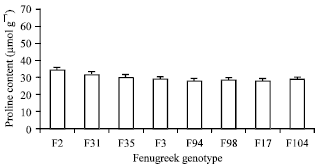 | |
| Fig. 5: | Chlorophyll content of 8 fenugreek cultivars under salt stress (513 mM), Values are Mean±SE |
Figure 4 and 5 represents the proline and chlorophyll content in 8 dS m-1 NaCl which showed highest values for both. Thus there values were used to the find correlation. From the results we can conclude that chlorophyll pigment may be involved with salt tolerance of fenugreek plant that accumulates proline to a high level to prevent damage from osmotic stress. Wanichananan et al. (2003) reported similar findings in salinity study of aromatic rice.
One of the most important mechanisms by higher plants under salt stress is the accumulation of compatible solutes such as proline. The accumulation of proline in salt stressed plants is a primary defense response to maintain the osmotic pressure in a cell. Several reports show a significant role of proline in osmotic adjustment, protecting cell structure and its function in plants in salt-tolerant and salt-sensitive cultivars of many crops (Desingh and Kanagaraj, 2007; Koca et al., 2007; Veeranagamallaiah et al., 2007; Turan et al., 2007).
Soil salinity, one of the most serious problems in many agricultural areas of Oman, has the most obstructive impact on crop production in other parts of the world. This crisis problem attracts many scientists to overcome this obstruction by improving salt-tolerant lines.
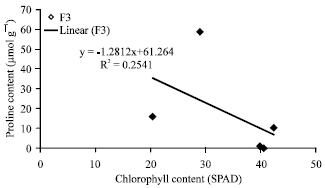 | |
| Fig. 6: | Correlation between proline and chlorophyll content of fenugreek accession F3 influenced by 8 dS m-1 NaCl dilution for 8 days |
The accumulation of proline in plants was first observed in wilted rye grass (Kemble and MacPherson, 1954). Proline is known to be an osmoregulatory solute in plants under hyperosmotic stress (Delauney and Verma, 1993). The hyperosmotic stress due to the drought and salinity is the most important limiting factor for plant growth and crop production (Boyer, 1982). The effect of salt stress on proline accumulation was reported in many plant species such as Beta vulgaris L. (Gzik, 1996), Lycopersicon esculentum (Guerrier, 1995), Brassica napus L. (Trotel et al., 1996), Oryza sativa L. (Lutts et al., 1999; Moons et al., 1995) and Morus alba L. cv. Khonpai (Harinasut et al., 2000).
Chlorophyll, found in all green plants, is the principal pigment for photosynthesis (Nobel, 1999). Chlorophyll degradation has been found affected by many stresses, leading to changes in certain enzyme activities, photosynthetic electron transport, carbon metabolism and photophosphorylation in photosynthesis. During salt stress, salt sensitive plants clearly showed chlorophyll degradation and growth reduction (Harinasut et al., 2000; Taiz and Zeiger, 1991).
ACKNOWLEDGMENT
This study was supported by the HM Research Fund grant number SR/AGR/CROP/07/01.
REFERENCES
- Mansour, M.M.F., K.H.A. Salama, F.Z.M. Ali and A.F. Abou Hadid, 2005. Cell and plant responses to NaCl stress in Zea mays L. cultivars differing in salt tolerance. Gen. Appl. Plant Physiol., 31: 29-41.
Direct Link - Misra, N. and A.K. Gupta, 2005. Effect of salt stress on proline metabolism in two high yielding genotypes green gram. Plant Sci., 169: 331-339.
CrossRefDirect Link - Misra, N. and A.K. Gupta, 2006. Interactive effects of sodium and calcium on proline metabolism in salt tolerant green gram cultivar. Am. J. Plant Physiol., 1: 1-12.
CrossRefDirect Link - Misra, N. and U.N. Dwivedi, 1990. Nitrogen assimilation in germinating Phaseolus aureus seeds under saline stress. J. Plant Physiol., 135: 719-724.
CrossRef - Misra, N. and U.N. Dwivedi, 1995. Carbohydrate metabolism during seeds germination and seedling growth in green gram under saline stress. Plant Physiol. Biochem., 33: 33-38.
Direct Link - Misra, N. and U.N. Dwivedi, 2004. Genotypic difference in salinity tolerance of green gram cultivars. Plant Sci., 166: 1135-1142.
CrossRefDirect Link - Moons, A., G. Bauw, E. Prinsen, M. van Montagu and D. van der Straeten, 1995. Molecular and physiological responses to abscisic acid and salts in roots of salt-sensitive and salt-tolerant Indica rice varieties. Plant Physiol., 107: 177-186.
CrossRef - Morgan, J.M., 1984. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Physiol., 35: 299-319.
CrossRefDirect Link - Redmann, R.E., 1974. Osmotic and specific ion effects on the germination of alfalfa. Can. J. Bot., 52: 803-808.
CrossRef - Sharma, M.L., 1976. Interaction of water potential and temperature effects on germination of three semi-arid plant species. Agron. J., 68: 390-394.
Direct Link - Skriver, K. and J. Mundy, 1990. Gene expression in response to abscisic acid and osmotic stress. Plant Cell, 2: 503-512.
CrossRefPubMedDirect Link - Tewari, T.N. and B.B. Singh, 1991. Stress studies in lentil (Lens esculenta Moench). II. Sodicity-induced changes in chlorophyll, nitrate and nitrite reductase, nucleic acids, proline, yield and yield components in lentil. Plant Soil, 136: 225-230.
CrossRefDirect Link - Trotel, P., A. Bouchereau, M.F. Niogret and F. Larher, 1996. The fate of osmo-accumulated proline in leaf discs of rape (Brassica napus L.) incubated in a medium of low osmolarity. Plant Sci., 118: 31-45.
CrossRef - Turan, M.A., N. Turkmen and N. Taban, 2007. Effect of NaCl on stomacal resistance and proline, chlorophyll, Na, Cl and K concentrations of Lentil Plants. J. Agron., 6: 378-381.
CrossRefDirect Link - Unnikrishnan, S.K., L. Prakash, P.C. Josekutty and A.R. Mehta, 1991. Effect of NaCI salinity on somatic embryo development in Sapindus trifoliatus L. J. Exp. Bot., 42: 401-406.
CrossRef - Veeranagamallaiah, G., P. Chandraobulreddy, G. Jyothsnakumari and C. Sudhakar, 2007. Glutamine synthetase expression and pyrroline-5-carboxylate reductase activity influence proline accumulation in two cultivars of foxtail millet (Setaria italica L.) with differential salt sensitivity. Environ. Exp. Bot., 60: 239-244.
CrossRef - Volkmar, K.M., Y. Hu and H. Steppuhn, 1998. Physiological responses of plants to salinity: A review. Can. J. Plant Sci., 78: 19-27.
CrossRef - Yeo, A., 1998. Molecular biology of salt tolerance in the context of whole-plant physiology. J. Exp. Bot., 49: 915-929.
CrossRefDirect Link - Zayed, M.A. and I.M. Zeid, 1997. Effect of water and salt stresses on growth, chlorophyll, mineral ions and organic solutes contents and enzymes activity in mung bean seedlings. Biol. Plant., 40: 351-356.
CrossRef - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Boyer, J.S., 1982. Plant productivity and environment. Science, 218: 443-448.
CrossRefPubMedDirect Link - Khanna-Chopra, R. and D.S. Selote, 2007. Accumulation to drought stress generates oxidative stress tolerance in drought-resistant than susceptible wheat cultivar under field conditions. Environ. Exp. Bot., 60: 276-283.
CrossRefDirect Link - De Lacerda, C.F., J. Cambraia, A. Oliva and H.A. Ruiz, 2005. Changes in growth and in solute concentrations in sorghum leaves and roots during salt stress recovery. Environ. Exp. Bot., 54: 69-76.
CrossRef - Delauney, A.J. and D.P.S. Verma, 1993. Proline biosynthesis and osmoregulation in plants. Plant J., 4: 215-223.
CrossRef - Demiral, T. and I. Turkan, 2005. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot., 53: 247-257.
CrossRefDirect Link - Desingh, R. and G. Kanagaraj, 2007. Influence of salinity stress on photosynthesis and antioxidative systems in two cotton varieties. Gen. Applied Plant Physiol., 33: 221-234.
Direct Link - Dodd, G.L. and L.A. Donovan, 1999. Water potential and ionic effects on germination and seedling growth of two cold desert shrubs. Am. J. Bot., 86: 1146-1153.
PubMedDirect Link - Dracup, M., 1991. Increasing salt tolerance of plants through cell culture requires greater understanding of tolerance mechanisms. Aust. J. Plant Physiol., 18: 1-15.
CrossRef - Asch, F., M. Dingkuhn, C. Wittstock and K. Doerffling, 1999. Sodium and potassium uptake of rice panicles as affected by salinity and season in relation to yield and yield component. Plant Soil, 207: 133-145.
CrossRefDirect Link - Fowler, J.L., 1991. Interaction of salinity and temperature on germination of crambe. Agron. J., 83: 169-172.
Direct Link - Ghoulam, C., A. Foursy and K. Fares, 2002. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot., 47: 39-50.
CrossRefDirect Link - Girija, C., B.N. Smith and P.M. Swamy, 2002. Interactive effects of sodium chloride and calcium chloride on the accumulation of proline and glycinebetaine in peanut (Arachis hypogaea L.). Environ. Exp. Bot., 47: 1-10.
CrossRefDirect Link - Guerrier, G., 1995. Effect of salt-stress on proline metabolism in calli of Lycopersicon esculentum, Lycopersicon pennellii and their interspecific hybrid. Can. J. Bot., 73: 1939-1946.
CrossRefDirect Link - Gzik, A., 1996. Accumulation of proline and pattern of α-amino acids in sugar beet plants in response to osmotic, water and salt stress. Environ. Exp. Bot., 36: 29-38.
CrossRefDirect Link - Haroun, S.A., 2002. Fenugreek growth and metabolism in response to gibberellic acid and seawater. Bull. Fac. Sci. Assiut Univ., 31: 11-21.
Direct Link - Kafi, M. and M. Goldani, 2001. Effects of water potential and type of gsmoticum on seed germination of three crop species of wheat, sugarbeet and chikpea. Agric. Sci. Technol., 15: 121-133.
Direct Link - Katembe, W.J., I.A. Ungar and J.P. Mitchell, 1998. Effect of salinity on germination and seedling growth of two Atriplex species (Chenopodiaceae). Ann. Bot., 82: 167-175.
CrossRefDirect Link - Kemble, A.R. and H.T. MacPherson, 1954. Liberation of amino acids in perennial rye grass during wilting. Biochem. J., 58: 46-59.
Direct Link - Koca, H., M. Bor, F. Ozdemir and I. Turkan, 2007. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot., 60: 344-351.
CrossRefDirect Link - De Lacerda, C.F., J. Cambraia, M.A. Oliva, H.A. Ruiz and J.T. Prisco, 2003. Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ. Exp. Bot., 49: 107-120.
CrossRefDirect Link - Lutts, S., V. Majerus and J.M. Kinet, 1999. NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant., 105: 450-458.
CrossRef - Hare, P.D., W.A. Cress and J. van Staden, 1998. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ., 21: 535-553.
CrossRefDirect Link - Bohnert, H.J. and R.G. Jensen, 1996. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol., 14: 89-97.
CrossRefDirect Link