
K. Shilpa
Centre for Biotechnology, Anna University, Chennai, Tamilnadu-600025, India
K. Varun
Centre for Biotechnology, Anna University, Chennai, Tamilnadu-600025, India
B. S. Lakshmi
Centre for Biotechnology, Anna University, Chennai, Tamilnadu-600025, India
Journal of Plant Sciences
Year: 2010 | Volume: 5 | Issue: 3 | Page No.: 222-247







ABSTRACT
Plant cells are factories of chemical compounds produced for carrying out biochemical pathways of survival and propagation. Primary metabolism involves biochemical processes for normal anabolic and catabolic pathways which results in assimilation, respiration, transport and differentiation whereas secondary metabolism involves generation of by-products which are the main defense elements of plants against pathogens, herbivore attacks and physical stress such as UV radiation. The secondary metabolites are plant pigments (such as alkaloids, isoprenoids etc.) responsible for the colors, flavors and smell in plants which also act as a source of drugs, fine chemicals, insecticides, dyes, flavours and fragrances. Plant derived secondary metabolites have played an essential role as medicine for thousands of years. Currently, secondary metabolites with bioactivity are being isolated and used either directly or after chemical modification. Their pharmacological value is increasing due to the constant discoveries of their potential roles in healthcare and as lead compounds for new drug development. This review highlights and discusses ways of improving the production of plant secondary metabolites and also focuses on plant cell culture which is considered as a promising alternative for producing bioactive compounds that are difficult to be obtained by chemical synthesis or plant extraction. It would be appropriate to consider the properties of plant cells in culture, in particular the relationship between growth and the expression of the pathways leading to the synthesis of secondary metabolites.
PDF Abstract XML References Citation
Received: March 29, 2010;
Accepted: May 09, 2010;
Published: July 27, 2010
How to cite this article
K. Shilpa, K. Varun and B. S. Lakshmi, 2010. An Alternate Method of Natural Drug Production: Elciting Secondary Metabolite Production Using Plant Cell Culture. Journal of Plant Sciences, 5: 222-247.
DOI: 10.3923/jps.2010.222.247
URL: https://scialert.net/abstract/?doi=jps.2010.222.247
DOI: 10.3923/jps.2010.222.247
URL: https://scialert.net/abstract/?doi=jps.2010.222.247
INTRODUCTION
Plants have been the source for many important drugs because they are able to produce various chemical entities and bioactive molecules through the process known as metabolism. Plant cell carries out both primary and secondary metabolism. Primary meta bolism involves synthesis of polysaccharides, proteins, lipids, RNA and DNA through utilization of sugars, amino acids, common fatty acids and nucleotides whereas secondary metabolism is activated only during particular stages of growth and development or during periods of stress, limitation of nutrients or attack by micro-organisms (Yazaki et al., 2008; Yeoman and Yeoman, 1996).
Secondary metabolites are generally derived from primary metabolites through modifications, such as methylation, hydroxylation and glycosylation. Therefore, secondary metabolites are naturally more complex than primary metabolites and are classified on the basis of chemical structure (e.g., aromatic rings, sugar), composition (containing nitrogen or not), their solubility in various solvents or the pathway by which they are synthesized. They have been categorised into Terpenes (composed entirely of carbon and hydrogen), phenolics (composed of simple sugars, benzene rings, hydrogen and oxygen) and nitrogen and/or sulphur containing compounds (Chinou, 2008) (Table 1). It has been observed that each plant family, genus and species produces a characteristic mix of these metabolites.
All plants produce secondary metabolites which are often specific to an individual species or genus during specific environmental conditions making their extraction and purification difficult. As a result, commercially available secondary metabolites (pharmaceuticals, flavours, fragrances and pesticides) are generally considered high value products as compared to primary metabolites and they are considered to be fine chemicals with their cost ranging from $500 to $10000 per kilogram (Ravishankar and Rao, 2000; Balandrin et al., 1985) (Table 2).
Secondary metabolites have played a significant role in medicine since ancient period. Plants have been directly used as food and herbs in the organised traditional medical systems such as Ayurveda, Unani and traditional Chinese medicine for the treatment of various diseases for thousands of years (Mukherjee et al., 2007; Fabricant and Farnsworth, 2001). Recently, the ethyl acetate extract of Alpinia officinarum has been reported interestingly of possessing dual property of anti-microbial and anti-inflammatory (Subramanian et al., 2008) and the methanol extract of Ocimum Basilicum has been reported for anti-inflamatory activity (Selvakkumar et al., 2007a). Drug discovery pursued using traditional knowledge helps reduce time and developmental cost to identify bioactive molecules (Patwardhan, 2000; Patwardhan et al., 2004; Jachak and Saklani, 2007). An anti-diabetic molecule has been identified from the methanolic extract of Costus pictus and currently investigation is on to isolate an anti-adipogenic molecule from the same extract (Shilpa et al., 2009).
The plant based drug discovery gained interest with the development of anticancer and anti-infectious agents and now contributes to new bioactive molecules which are being isolated for the treatment of various other diseases including metabolic disorders like Diabetes and Obesity (Saklani and Kutty, 2008) such as Metformin, a commercially available anti-diabetic drug derived from Galega officinalis (Oubre et al., 1997).
| Table 1: | Classification of secondary metabolites |
 | |
| Table 2: | Cost estimation for plant-derived secondary metabolites of commercial importance in pharmaceutical industry |
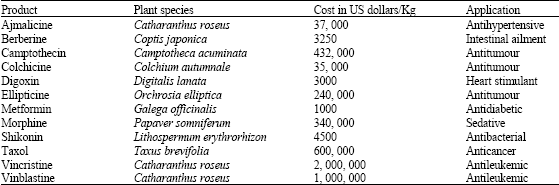 | |
| Ravishankar and Rao (2000) | |
Cinnamic acid, Methyl tetracosanoate, Tannins, Caffeoyl derivatives, 3β-taraxerol, Aloe emodin and Stigmasterol have been isolated showing good anti-diabetic potential and 12-ursene 2-diketone has been isolated which exhibited anti-inflammatory property in in vitro models (Lakshmi et al., 2009; Muthusamy et al., 2008, 2010; Shilpa et al., 2009; Sangeetha et al., 2010; Anand et al., 2010; Sujatha et al., 2009; Gayathri et al., 2007).
The plant as a source for important drug molecules is witnessed through the discoveries of various bioactive molecules such as Taxol, Vincristine, Vinblastine, Metformin, Morphine etc. The synthetic aspirin used by the modern world today is a derivative of a plant-based drug (Raskin and Ripoll, 2004). Many new plant-based drugs such as Prostratin, CAPSOROLS, CCS and over 60 anti-cancer compounds are under active preclinical trials alongwith with the introduction of herbals in the form of nutraceuticals and dietary supplements (Corcoran and Spraul, 2003; Steinbeck, 2004; Ganesan, 2002; Saklani and Kutty, 2008).
Important plant-derived drugs are still obtained commercially by extraction from their whole plant sources. A loss in activity has been observed when these compounds have been attempted to be chemically synthesized; this could be because, the secondary metabolites generally have highly complex structures with many chiral centers which may contribute to their biological activity and such complex compounds are difficult to synthesize economically (Pezzuto, 1995; Kolewe et al., 2008).
Currently, the natural habitats of many plants are disappearing due to environmental and geopolitical instabilities, therefore making it very difficult to acquire important secondary metabolites and in the process leaving many potentially useful compounds left undiscovered. Plant cell culture is considered as a promising alternative for producing bioactive compounds that are difficult to be obtained by chemical synthesis or plant extraction (Kolewe et al., 2008). Plant cell culture studies have been carried out based on the fact that each plant cell in the culture exhibits totipotency, wherein the cell has the full set of genes necessary for all the functions of a plant, including secondary metabolism (Verpoorte et al., 1999). Cell culture systems are useful in large-scale culturing of plant cells, which form a continuous and reliable source of secondary metabolites and can be purified easily due to the absence of significant amounts of pigments. This method removes all seasonal constraints and eliminates the geographic barriers for production of secondary metabolites (Karuppusamy, 2009). Plant tissue culture has been applied for the production of secondary metabolites on a commercial scale since late 1950s, when atropine from the roots of Atropa belladonna was synthesized and accumulated in roots and in callus (West-jr and Mika, 1957).
One Major limitation in the production of secondary metabolites by plant cell culture technology is the low yield of secondary metabolites. This could be improved by: 1) increasing the productivity of cultured plant cells and the necessary metabolite through standardization of culture environment (Ramachadra-Rao and Ravishankar, 2002); 2) manipulation of plant cell cultures to improve productivity of target compounds employing elicitors, abiotic stresses and other approaches, regardless of their mechanisms (Zhong, 2001; Zhao et al., 2005); 3) studying signal transduction pathways underlying various effective strategies leading to the biosynthesis of target secondary metabolites (Memelink et al., 2001; Zhao et al., 2005).
MANIPULATION OF CULTURE ENVIRONMENT FOR IMPROVED BIOMASS AND SECONDARY METABOLITE PRODUCTION
In order to obtain products at high concentrations for commercial manufacturing, standardization of chemical factors like media components, plant growth regulators etc and physical factors like pH, temperature, light, etc. play an important role (Dornenburg and Seydel, 2008; Karuppusamy, 2009). Enhancement of the yield of biomass and secondary metabolite production through plant cell culture also requires selection of proper genotypes and high-yielding clones (Barz and Ellis, 1981; Mulabagal and Tsay, 2004).
Selection of Proper Genotypes and High-Yielding Cell Clone
It has been observed that collection of cells from source explant as inoculum to the production medium is crucial. Cell cultures derived from wild plants have been observed to accumulate large amount of secondary metabolites which could be a good source for the isolation of the bioactive secondary metabolite.
After the cell culture has been established, plant cells undergo a continuous process of epigenetic or genetic changes during culture which represents a heterogeneous population. It is therefore essential to select cell clones which produce high yields as well as accumulate high amount of the desired metabolite (Ogino et al., 1978). This strategy resulted in an increased accumulation of anthocyanins and berberin compared to that of the mother culture in Euphorbia milli and Coptis japonica, respectively (Yamamoto et al., 1982; Yamada and Sato, 1981).
Medium
Medium standardization has been an important parameter in plant cell culture technology since the composition of the culture media is known to influence the biomass yield as well as secondary metabolite production. Medium composition, culture conditions and exogenous phytohormone combinations together influence the metabolite accumulation in the cell (Seabrook, 1980). Although Murashige Skoog (1962) Medium is one of the most widely used medium for rapid growth of callus, it has not been observed as an ideal medium for inducing secondary metabolite accumulation.
Macro and micro nutrient composition in plant tissue culture media has foremost effect on both primary and secondary metabolism of cells (Dougall, 1980) and a medium which leads to rapid cell division and limits early cessation of exponential growth is thought to be helpful for secondary metabolite formation. An overall level of total nitrogen has been found to affect production of secondary metabolites. For example, reduced levels of ammonia and increased nitrate levels were found to promote the production of shikonin and betacyanins, whereas higher ammonia to nitrate ratio was found to increase the production of berberine and ubiquinone (Bohm and Rink, 1988; Nakagawa et al., 1984; Fujita et al., 1981; Ikeda et al., 1977). It has also been observed that reduced levels of total nitrogen improved the production of anthraquinones in Morinda citrifolia and anthocyanins in Vitis species (Yamakawa et al., 1983; Zenk et al., 1975). Apart from the nitrogen source, phosphates were also found to play an important role in increasing the secondary metabolite production. It had been observed that added phosphates in the culture medium resulted in prolonged cell growth in Catharanthus Roseus cultures (Mac-Carthy et al., 1980), similar results were obtained in a separate study for the production of betacyanins in callus cultures of Beta vulgaris (Bohm and Rink, 1988). Increased phosphate also stimulated the production of digitoxin in Digitalis purpurea (Hagimori et al., 1982).
Cell cultures are also influenced with simple sugars as carbon source along with inorganic supply of other nutrients. Sucrose and glucose are most preferred amongst other carbon sources for plant tissue culture media. The carbon source has been known to influence cell growth and yield of secondary metabolites in many cases (Dornenburg, 2008). It has been observed that a range of 2 to 8% of carbon source is appropriate for optimal secondary metabolite production in plant cell cultures (Ramachadra-Rao and Ravishankar, 2002). The maximum yield of rosmarinic acid produced by cell suspension cultures of Salvia officinalis was 3.5 g L-1 when 5% of sucrose was used but it was 0.7 g L-1 in the medium containing 3% sucrose (Whitaker et al., 1984).
Plant Growth Regulators
In a plant cell and/or tissue culture study, growth regulator concentration plays a crucial role in secondary metabolite accumulation (Di-Cosmo and Towers, 1984). The concentration of auxin and cytokinin individually or in combination significantly alters both the growth and the secondary metabolite accumulation in cultured cells (Mantell and Smith, 1984). The growth regulator 2, 4-dichlorophenoxyacetic acid (2, 4-D) has been found to play an important role in callus initiation, but it has been found to inhibit the production of secondary metabolites in a large number of cases. However, there have also been reports where cultures grown in the presence of 2, 4-D have shown secondary metabolite induction for eg. carotenoid and anthocyanin production in callus cultures of Daucus carota (Rajendran et al., 1992) and Oxalis linearis (Meyer and Staden, 1995), respectively. In cases where secondary metabolite production is inhibited, elimination of 2, 4-D or replacement of 2, 4-D with other auxins such as Naphthalene Acetic Acid (NAA) or Indole Acetic Acid (IAA) has been found to enhance the production of nicotine in suspensions of Nicotiana tabacum and shikonin in suspensions of Lithospermum erythrorhizon (Sahai and Shuler, 1984; Fukui et al., 1983). Cytokinins have been observed to have different effects depending on the type of plant species and metabolite. Kinetin has been found to stimulate the production of anthocyanin in Haplopappus gracilus but inhibited the formation of anthocyanins in Populus cell cultures (Seitz and Hinderer, 1988; Mok et al., 1976). Apart from auxins and cytokinins, other plant growth regulators such as Gibberellic acid and abscisic acid have been reported to suppress production of anthocyanins in a number of cultures (Bohm and Rink, 1988; Seitz and Hinderer, 1988).
pH, Light and Temperature
In addition to the necessary salt, sugar and exogenous phytohormone concentrations in media, pH also plays an important role on cell growth and metabolite production. Optimum growth of plant cells in the culture is usually obtained between pH range of 5.5 and 6.0. It has been known for some time that the pH of the culture medium can influence the uptake of nutrients and precursors, the permeability of membranes and release of products from the vacuole to the culture medium. Concentration of hydrogen ions in the medium is known to change during the development of the culture. The medium pH decreases during ammonia assimilation and increases during nitrate uptake (Mc-Donald and Jackman, 1989). Photoautotrophic cell suspension cultures of Chenopodium rubrum L. showed an increase in biomass between pH 5.0 to 6.0 (Husemann et al., 1992).
There are reports of light stimulateSelvakkumard secondary metabolites production (Whitaker et al., 1986) and for each plant species different temperatures favour the production of secondary metabolite. It has been observed that white light has a significant effect on cell growth as well as on chlorophyll and kalata B1 synthesis in Oligodon affinis cell cultures. In callus cultures the optimum irradiation intensity for growth was found to be lower than that required for production of secondary metabolite, which was found to increase with increasing intensities of light (Dornenburg and Seydel, 2008). Light was found to stimulate accumulation of anthocyanin in cell cultures of Daucus carota and Vitis hybrids (Seitz and Hinderer, 1988), on the contrary, elimination of light prompted the accumulation of monoterpenes in callus cultures of Citrus limon (Mulder-Krieger et al., 1988).
Temperature ranging between 17-25°C are normally used for induction of callus and growth of cultured cells but Toivonen et al. (1992) found that lowering the cultivation temperature increased the total fatty acid content per cell of dry weight. Studies have also shown that at 32°C, tobacco cell cultures produced higher yield of ubiquinone when compared to either 24 or 28°C (Ikeda et al., 1977). Cell cultures of Catharanthus roseus produced 12-fold higher crude alkaloids at 16°C as compared to the normal 27°C (Courtois and Guren, 1980).
MANIPULATION OF PLANT CELL CULTURES FOR INCREASED SECONDARY METABOLITE PRODUCTION
Altogether it is the appropriate combination of all the above mentioned parameters, which would aid in a cell culture capable of producing high biomass yield along with increased secondary metabolite production (Table 3). After medium optimization, the subsequent strategies vary based on the type of product needed, like organ cultures, product absorption and secretion, biotransformation, cell immobilization, mutagenesis, metabolic engineering etc.
Organ Culture
It is generally found that in plants high concentrations of secondary metabolites tend to accumulate in specific cell types at specific developmental stages, therefore tissue cultured cells from these plants typically accumulate large amounts of secondary compounds under specific conditions (Robins et al., 1986). Although, a wide range of secondary metabolites are produced through de-differentiated callus, there exist certain metabolites which are organ specific and require differentiated organ cultures (Table 3) when the metabolite is produced only in specialized plant tissues or glands in the wild plant for example, Hypericum perforatum (St. John’s wort), accumulates the anti-depressant hypericins and hyperforins in foliar glands and have no reports of their ability to accumulate phytochemicals in undifferentiated cells (Smith et al., 2002). It has also been observed that Nicotiana tabacum (tobacco) roots are source for biosynthesis of lysine to anabasine, followed by the conversion of anabasine to nicotine in leaves. Herein the callus and shoot cultures of tobacco have been found to produce only trace amounts of nicotine because they lack the organ-specific compound anabasine (Karuppusamy, 2009).
Product Absorption and Secretion
Secondary metabolites produced during cell culture studies have been reported to be accumulated intracellularly and there are also reports of metabolites which are released into the culture medium (Misawa, 1994).
In certain cases the product of interest might be accumulated intracellularly and its production might sometimes inhibit its own synthesis through product inhibition and repression. In some cases higher level of product can be produced when secreted into the medium. In such situations permeabilization of cell membranes has been attempted using permeabilizing agents such as DMSO, Triton X-100, electroporation or ultrasonication, for enhancing the product release, but has achieved limited success.
| Table 3: | In vitro secondary metabolites production from plant cell, tissue and organs cultures using different media and plant growth regulator combinations |
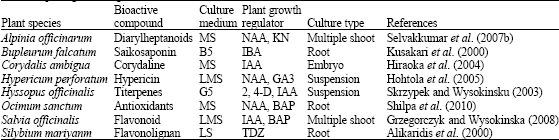 | |
Viability of the cells was observed to be lost in most cases when permeabilizing agents and electroporation were used (Brodelius, 1988). Chenopodium rubrum cultures which secrete amaranthin (red betacyanin pigment) when incubated with low concentrations of chitosan (0.01%) and DMSO (5.7%) for 96 h showed no significant effect on cell viability, but a longer incubation period (196 h) was found to have a deleterious effect (Knorr and Berlin, 1987). Ultrasonication was found to be a better method in comparison wherein Beta vulgaris cells upon ultrasonification for 20 to 60 sec was found to aid the release of betanin with no apparent effect on cell viability (Kilby and Hunter, 1990).
Most secondary metabolites produced in cell cultures are generally accumulated intracellularly, but some metabolites have also been reported to be secreted into the media such as taxol produced by Taxus brevifolia (Collins-Pavao et al., 1996). In such cases XAD-7 (resin) is added in the suspension medium and the metabolites (pigments) are found to be absorbed into it. Addition of XAD-7 (resin) into its suspension culture of Chinchonaledgerina also stimulated the production of anthraquinones with a 15 fold increase compared to the medium without resin (Robins et al., 1986). Similarly, active charcoal in the medium was also found to stimulate the product yield (Misawa, 1994)
Biotransformation
Biotransformation is a process through which the functional groups of organic compounds are modified either stereo specifically or region specifically by living cultures, entrapped enzymes or permeabilized cells to a chemically different products. The advantage of this method includes the production of novel compounds, enhancement in the productivity of desired compound and overcoming the problems associated with chemical synthesis. The production of fine chemicals and pharmaceuticals can be achieved through biological catalysts in the form of enzymes and whole cells (Ravishankar and Rao, 2000; Meyer et al., 1997).
A suitable substrate compound can be biotransformed to obtain a desired product using plant cells. There are two major reasons to choose plant cells for biotransformation purposes. To start with, these cells are generally able to catalyze the reactions stereospecifically resulting in chirally pure products. Secondly, they have the ability to perform regiospecific modifications that can not be easily carried out through chemical synthesis or by microorganisms (Hamada and Furuya, 1996). Plant cells perform reactions such as reduction, oxidation, hydroxylation, acetylation, esterification, glucosylation, isomerization, methylation, demethylation, epoxidation, etc. This process should meet four important prerequisites that the culture must have the necessary enzymes. the substrate or precursor must not be toxic to the culture and that the cellular compartment of the cell and importantly the rate of product formation must be faster than its further metabolism (Table 4).
| Table 4: | Production of pharmaceuticals in plant cell cultures using Biotransformations |
 | |
Immobilization
Immobilization is a technique, which confines a catalytically active cell on a fixed support and prevents cell’s entry into the suspension culture (Yeoman, 1987). This technique involves entrapment of cells in some kind of gel or combination of gels, which are allowed to polymerize around them, e.g., of gels include calcium alginate, agar, agarose, gelatin, carrageenan and polyacrylamide; alternative supports include polyurethane foam and hollow fibre membranes. Amongst them, gels of alginate are most widely used because of their simplicity and lack of toxicity. There is a necessity that immobilized cells should maintain prolonged viability and biosynthetic capability with high rates of secondary metabolite production. Catharanthus roseus had been subjected to two types of immobilization system, one was immobilization in polysaccharide beads (Brodelius and Nilsson, 1980) and the other polyacrylamide sheets (Lambe and Rosevear, 1982), both of which exhibited alkaloid release with no loss in viability. Capsicum frutescens cells immobilized on polyurethane were found to release capsaicin completely into the medium, though other species immobilized by the same method showed retention of their product intracellularly (Lindsey and Yeoman, 1984). Chenopodium rubrum cells when immobilized in alginate beads were found to secrete the red betacyanin pigment, amaranthin into the medium (Knorr and Berlin, 1987). De novo biosynthesis of many important secondary metabolites is carried out through single and/or multistep biotransformation of precursors on immobilized plant cells. Product synthesis should not be strictly growth associated as growth of cells can lead to disintegration of the immobilization matrix, which may lead to interruption of the process and it is essential that a significant amount of product is released into the medium. There has been many more studies conducted using immobilization (Table 5a, b).
Elicitation of Secondary Metabolites
Various studies towards enhancement of secondary metabolite production in cell cultures include treatment with various precursors, elicitors, signal compounds and abiotic stresses.
Precursors
It has been noticed that addition of precursors or related compounds (which intermediate at the beginning of a secondary metabolite biosynthetic route) to the culture media sometimes stimulate secondary metabolite production (Mulabagal and Tsay, 2004). This is thought to occur as any precursor would stand a good chance of increasing the yield of the final product. For example, Salvia officinalis cell suspension cultures and Taxus cultures stimulated rosmarinic acid and taxol production respectively upon addition of phenylalanine (Ellis and Towers, 1970; Fett-Neto et al., 1993). This approach could be advantageous if the precursors are inexpensive.
| Table 5a: | Use of immobilized plant cell systems for production of secondary metabolites |
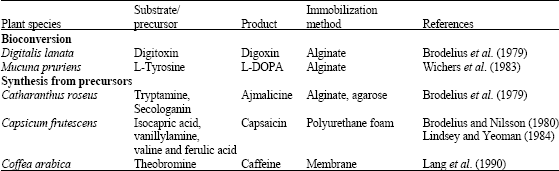 | |
| Table 5b: | Use of immobilized plant cell systems for production of secondary metabolites through De novo synthesis |
 | |
Elicitors
Plant secondary metabolites are majorly produced as a response to protect plants from environmental stresses. The environmental stresses (microbial, physical or chemical factors) leading to an increase in secondary metabolism are known as elicitors.
The use of elicitors in cell cultures has been developed as one of the main strategies to improve the yield of secondary metabolites wherein elicitation is induced by the addition of trace amounts of elicitors (Radman et al., 2003; Roberts and Shuler, 1997). Elicitors are signals triggering the formation of secondary metabolites and are classified based on their nature into abiotic or biotic elicitors (Mulabagal and Tsay, 2004; Namdeo, 2007).
Elicitors of non-biological origin are called abiotic elicitors, which predominantly consists of physical and chemical stresses such as UV radiation extremes of temperature, ethylene, fungicides, antibiotics, salts of heavy metals or high salt concentrations etc. (Table 6).
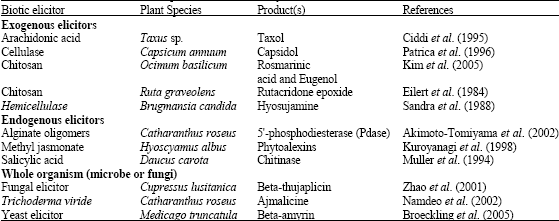
Other class of elicitors are biotic elicitors; these are substances with biological origin such as polysaccharides derived from plant cell walls (pectin or cellulose) and micro-organisms (chitin or glucans) and glycoproteins or intracellular proteins which act by activating or inactivating a number of enzymes or ion channels (Rokem et al., 1984). This class of elicitors are categorized into two types based on their origin, exogenous and endogenous elicitors (Table 7). The elicitors which have originated outside the cell like polysaccharides, polyamines and fatty acids are known as exogenous elicitors whereas endogenous elicitors have originated inside the cell like galacturonide or hepta-β-glucosides etc. (Namdeo, 2007). Evidently, in certain circumstances elicitation can be used to obtain better consequence in increasing product yield and has also been found to have commercial potential.
During the process of elicitor study, it should be noted that cells showing failure to elicit necessary secondary metabolites does not necessarily mean that the metabolic pathway cannot be triggered. Inappropriate combination of medium and elicitor, as well as unsuitable concentration of the elicitor could be a cause for unsuccessful elicitation, which indicates that a successful elicitation is a very challenging process requiring intense standardizations (Namdeo, 2007; Zhao et al., 2005). In several studies it has been observed that elicitor treatments performed at the late log phase result in the high biomass yields along with secondary metabolite production as almost all the elicitors, when used at early log phase showed immediate increase in the secondary metabolites while suppressing the biomass leading to overall low productivity. For commercial production of secondary metabolites, apart from the secondary metabolite enhancement, it is equally important to study the signals involved in the process of elicitation, which might help in choosing the appropriate elicitor (Zhao et al., 2005).
| Table 6: | Abiotic elicitors used for production of secondary metabolites |
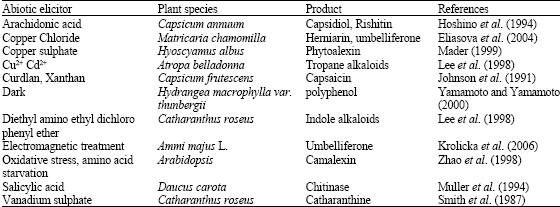 | |
| Table 7: | Biotic elicitors used for the production of secondary metabolites |
 | |
Stress Related Signal Transduction Pathways
Plants subjected to stresses (including various elicitors or signal molecules) lead to an accumulation of a variety of secondary metabolites. It is important to study these stress related signal transduction thereby helping in the development of strategies for commercial production of the target compounds by either activation or suppression of certain metabolic pathways making it a powerful tool to investigate pathway regulation based on gene expression.
Upon challenge by the biotic elicitors, plants generally stage an array of defence or stress responses. The recent studies are focused on studying the recognition of elicitation stimulus and the subsequent triggered defence response. Signal transduction of plant defence response primarily involves the host (cell) recognition to elicitors which initiates the early signalling events such as protein phosphorylation or dephosphorylation, changes in ion fluxes and oxidative burst. These processes stimulate the subsequent transcriptional activation of lant defence genes, which involves activation of enzymes such as Glutathione-S-Transferases (GST), Phenyl Ammonium Lyase (PAL), Chalcone synthase (CHS) etc., biosynthesis of endogenous secondary signals such as salicylic acid etc. and activation of NADPH oxidase leading to complex generation of reactive oxygen species (ROS) such as O2.-, H2O2 and thereby altering the redox status of plant cells affecting the defense signalling. ROS have been found to be generated after any elicitation process. Plants have evolved efficient antioxidant systems to scavenge ROS (Fig. 1). The induction /suppression of antioxidant activities provide evidence for occurrence of oxidative burst and variation of secondary metabolite accumulation (Yang et al., 1997).
The interaction of elicitors with cellular receptors and subsequent transcriptional and postranslational activation of transcription factors thereon lead to the induction of defense genes (Zhu et al., 1996). In addition to eliciting primary defense responses, elicitor signals may be amplified through the generation of secondary plant signalling molecules such as Salicylic Acid (SA) (Durner et al., 1997). Elicitors and secondary endogenous signals generally activate a diverse array of plant defense and protector genes, whose products include glutathione S-transferases (GST), peroxidases, cell wall proteins, proteinase inhibitors, hydrolytic enzymes (e.g., chitinases and ~-1, 3-glucanases), pathogenesis-related (PR) proteins and phytoalexin biosynthetic enzymes, such as Phenylalanine Ammonia Lyase (PAL) and chalcone synthase (CHS) (Hammond-Kosack and Jones, 1996). Phytoalexins are low-molecular-weight, antimicrobial compounds (e.g., phenylpropanoids, terpenoids, etc.), whose synthesis is induced following biotic elicitation. Furthermore, signaling components such as G proteins, NADPH oxidase, H2O2, SA, mitogen-activated protein kinases (MAPK) and Myb transcription factors have been found to participate in several plant defense responses (Fig. 2). The appearance of cell death and production of ethylene also play a regulatory role in the elicitor induced defence responses of plants (Yang et al., 1997).
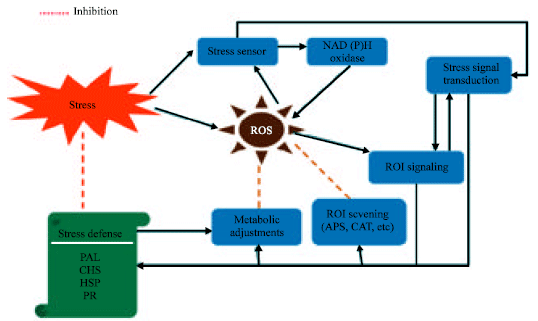 | |
| Fig. 1: | Mechanisms of ROS production and scavenging during biotic or abiotic stresses. Ascorbate peroxidase (APX) and catalase (CAT) are some of the key ROS scavenging enzymes of plants and heat shock proteins (HSPs); pathogenesis related proteins (PR); phenylalanine ammonia-lyaze (PAL), chalcone synthase (CHS); nicotinamide adenine dinucleotide phosphate-oxidase (NAD(P)H oxidase) are some of the important stress related genes |
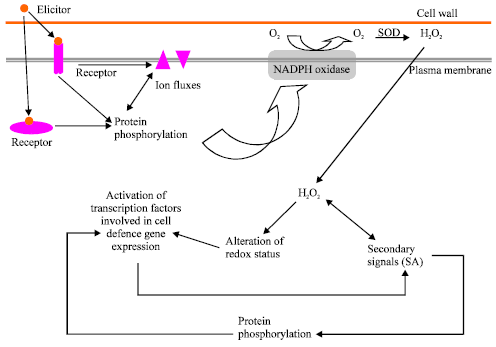 | |
| Fig. 2: | Signal transduction involved in plant defense responses. Cell recognizes elicitors and initiates early signaling events. Subsequent transcriptional and posttranslational activation of transcription factors leads to induction of plant defense genes and biosynthesis of endogenous secondary signals. Additionally, the activated NADPH oxidase complex generates reactive oxygen species (ROS), which alter the redox status of plant cells and affect defense signaling |
Host Recognition of Biotic Elicitors
When cells are incubated with an elicitor, the cell must recognize the elicitor and subsequent defense responses are initiated so as to limit the potential damage that could possibly be caused by the elicitor. Receptor (R) genes present in plants have been found to encode receptors for the recognition of specific elicitors (Gabriel and Rolfe, 1990), but there are many fungal and bacterial oligosaccharides, proteins and glycoproteins that have been found to act as nonspecific elicitors to induce defense responses in the plants cells carrying no specific R genes (Benhamou, 1996; Hahn, 1996).
Role for Protein Phosphorylation in Intracellular Signalling
Protein kinases and phosphatases have been found to play a pivotal role in early as well as later events of plant defense signaling pathways which indicates that phosphorylation and dephosphorylation also play an essential role for defence responses (Yang et al., 1997). Kinase cascades of the Mitogen-Activated Protein Kinase (MAPK) class of enzymes play an extremely important role in plant signalling during various stress conditions. MAPK plays a key role in induction of defence mechanisms by amplifying pathogen-derived signals perceived at membrane receptors and transducing these signals into altered gene expression (Pitzschke et al., 2009) (Fig. 3). Fungal elicitors trigger rapid and transient protein phosphorylation in cell suspension cultures of parsley (Dietrich et al., 1990) and protein kinase inhibitors like K-252a and staurosporine have been found to block these elicitor induced changes in protein phosphorylation and prevent the induction of plant defense responses. In contrast, many protein phosphatase inhibitors such as calyculin A,cantharadin and okadaic acid, imitate elicitor action to activate defense responses (Felix et al., 1994; Levine et al., 1994; Mackintosh et al., 1994).
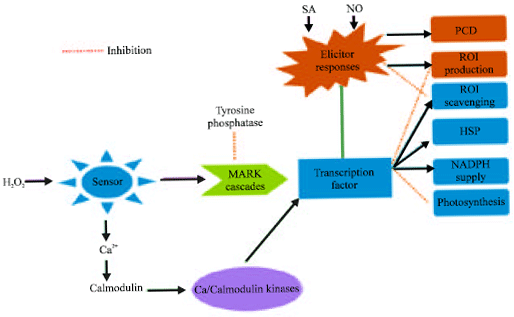 | |
| Fig. 3: | The signaling pathway activated in plants in response to external application of oxidants. SA, salicylic acid; NO, nitric oxide; H2O2, hydrogen peroxide; PCD, programmed cell death; HSP, heat shock protein; MAPK, mitogen-activated protein kinase |
Early Signaling Events
In addition to protein phosphorylation, early signaling events in defense responses involve ion fluxes (ion channels), oxidative burst (ROS), GTP-binding proteins, phospholipases etc.
Oxidative burst is another prominent event occurring early after elicitation. It leads to the rapid production and accumulation of ROS, such as O2.-and H2O2 (Doke et al., 1996; Low and Merida, 1996). Elicitors induced oxidative burst appears to be mediated by multiple signaling cascades, which might be associated with activation of G proteins, Ca2+ influx, H+/K+ exchange, induction of phospholipases and protein phosphorylation (Low and Merida, 1996). ROS generation is apparently dependent on the activation of a plasma membrane NADPH oxidase. Specific inhibitors of NADPH oxidase such as diphenylene iodonium, prevent plant ROS production induced by elicitors. ROS production occurs in two distinct phases which have been observed during cell-elicitor interactions. The first burst occurs within minutes whereas the second, sustained burst occurs after few hours of treatment with elicitors (Laxalt et al., 2007; Manickavelu et al., 2010). Oxidative burst generates superoxide anion and H2O2 and together they play multiple roles in defense responses. Furthermore, they act as a secondary messengers to induce plant defense-related genes (Orozco-Cardenas et al., 2001) and hypersensitive host cell death (Levine et al., 1994; Yoda et al., 2003). The action of ROS is thought to be mediated through changes in cytosolic Ca2+ levels and generation of lipid peroxides. Several studies have shown that Ca2+ plays an important role in defence signalling. Ca2+ channel blockers were shown to inhibit ion fluxes as well as defense responses induced by fungal and bacterial elicitors (Garcia-Brugger et al., 2006; Mc-Anish and Schroeder, 2009) (Fig. 3). ROS is found to activate defense gene expression by altering the redox status of plant cells, thereby modulating the activity of redox-sensitive transcription factors. ROS has been proven to play an important role in plant defense signaling, although their mechanism of action is poorly understood. In few cases, GTP-binding proteins have also found to participate in the induction of ion fluxes and the oxidative burst by some fungal elicitors (Xing et al., 1997; Garcia-Brugger et al., 2006).
Endogenous Secondary Signals in Plant Cell Defense
The elicitor signals are often amplified through the production of secondary signal molecules such as SA, ethylene and jasmonates subsequent to the early signaling events activated by elicitor treatment. SA is known to play an important role in the activation of defense responses. Increase in the levels of SA and its conjugates have been noticed during the activation of defence responses. The increase in SA levels slightly leads to parallel expression of PR genes in cells (Malamy et al., 1990). In addition to SA; ethylene, Jasmonic Acid (JA) and systemin have also been proved to be important secondary signals for plant defense responses (Kotchoni and Gachomo, 2006). Ethylene levels have been found to increase during the HR and have been eminent to induce the expression of PAL and the basic PR genes (Hammond-Kosack and Jones, 1996).
Integration of Signaling Pathways and Activation of Plant Defense Genes
The initial recognition and early signalling events may be different for various elicitors, however, many of these signals are incorporated into one of the various pathways that lead to the transactivation steps involved in the interaction between activated transcription factors and elicitor responsive cis elements in the promoters of defense genes. A single elicitor can activate multiple transcription factors that interact with different cis elements in the same or different promoters, leading to activation of many defense genes (Zhu et al., 1996). Some of these transcription factors are transcriptionally and/or post-translationally activated by elicitor or treatment with secondary signals, such as SA.
Techniques are therefore available for the identification of targets (i.e., genes, proteins, metabolites) as well as for their development. Various tools have been employed to identify unknown genes as well as to characterize secondary metabolite pathway regulation, including precursor feeding, gene over expression, application of metabolic inhibitors and mutant selection.
It is observed that the amount of the enzyme and its activity is directly proportional to the rate limiting step, therefore the amount of product formed is ultimately determined by the flux at the rate limiting step. The flux is generally influenced by substrate availability and end-product inhibition. When the respective biosynthetic pathway is known, then metabolic engineering can be performed to enhance the necessary product by increasing the activity of the enzymes which are limiting in the pathway, regulatory genes expression can be induced; the competitive pathways and catabolism can be blocked.
The former two possibilities require the enhanced expression of genes yielding active enzymes and the latter two approaches need antisense genes for blocking the genes involved in competitive pathway or catabolism. The most important regulatory step in secondary metabolism is transcription of the biosynthetic genes. Concentration of precursors and end products are determining factors in the metabolic control of synthesis and breakdown of the compounds, as all secondary metabolic processes derive their precursors from primary metabolism (Verpoorte et al., 1999).
Genetic methods can be applied to increase the production of the necessary metabolite or a group of related compounds through the expression of genes encoding key enzymes (Dueckershoff et al., 2005; Tian and Dixon, 2006; Inui et al., 2007; Liu et al., 2007) or through functional genomics (Goossens et al., 2003). The end goals of the genetic modifications generally include enhancement in the levels of necessary secondary metabolites along with lowered levels of unwanted or generation of novel compounds. Despite successes, metabolic engineering of the relevant biosynthetic pathways is often hampered by a lack of knowledge of the enzymes and genes involved (Chaudhuri et al., 2009).
Hairy Root Culture
The use of Agrobacterium rhizogenes for production of hairy root has been used as yet another indirect genetic approach for increasing the secondary metabolite production in a variety of species. Agrobacterium rhizogenes inserts the Ri plasmid into wounded tissue and induces the growth of very fine adventitious roots called hairy roots. These roots can be cultured in hormone-free medium and there are several examples of enhanced accumulation of secondary products, relative to non-transformed tissue. This transformation occurs through Ri T-DNA of Agrobacterium rhizogenes, which produces roots that can further be cultured in vitro, which often show high accumulation of secondary metabolites (Guillon et al., 2006). Many dicot species show similar morphological and physiological changes by the Ri T-DNA; thus, Ri T-DNA allows secondary metabolite production in multiple species without any knowledge about the mechanism of production (Chaudhuri et al., 2009) (Table 8).
Apart from the general increase in the secondary metabolite production, Elicitation, precursor feeding, cell permeabilization and immobilization are also considered efficient ways to improvement of secondary metabolite productivity of hairy roots (Table 9). And recently there has been an increased research in the field of hairy root metabolic engineering, where this study is based on integrating genes that encode enzymes of a given pathway between the T-borders of the Ri plasmid, then transferring this construct into plant
| Table 8: | Pharmaceutical products produced using hairy root cultures |
 | |
| Table 9: | Increased metabolite production using various elicitors in hairy root cultures of various plant species |
 | |
| genome (Palazon et al., 2003; Moyano et al., 2003; Hughes et al., 2004; Peebles et al., 2005; Hong et al., 2006). | |
CONCLUSIONS
Many plant derived drugs are still obtained commercially by extraction from their whole plant sources. However, their over-utilization is depleting many important plant sources, leading to serious ecological problems. Plant cell culture provides a continuous, reliable source of natural products. There are only few drugs which are produced commercially using plant cell culture, due to the limitations such as low yields of desired metabolites and biochemical and genetic instability of the plant cells. The factors playing an important role in overcoming the limitations include standardization of chemical and physical parameters for producing high yield of cell mass as well as the secondary metabolite. Secondly, plant cells are subjected to precursors and elicitors, so as to increase the secondary metabolite production. Finally, it becomes important to understand the mechanism by which the secondary metabolites are triggered or produced during the above mentioned conditions.
Despite the increasing demands of plant secondary metabolites, much of plant secondary metabolism is still poorly understood. As secondary metabolites are expressed in response to a variety of stresses, therefore it becomes important to focus on regulatory machinery of plant defense responses. Plants utilize multiple signaling pathways, such as jasmonic acid, ethylene, salicylic acid signaling pathways, to regulate defense response against elicitor or various stresses. Therefore, quantitative profiling of signal in response to various abiotic or biotic elicitors may help detect the initiation of elicitation of secondary metabolites. Elucidation of the signaling network would help in specific and efficient engineering of the production of target secondary metabolites. Molecular biology techniques are being used recently to produce transgenic cultures and to effect the expression and regulation of biosynthetic pathways. This would be a significant step towards commercial production of secondary metabolites using cell cultures. Apart from these methods, there exists another technology called the hairy root technology. Hairy roots grow easily upon infection with a bacterium and are also easy to transform. Their genetic and biochemical stability offer substantial advantages over cell suspensions. Goossensnerally, hairy roots accumulate increased amounts of secondary metabolite. Therefore, using elicitors or by performing metabolic engineering to the hairy roots can produce increased amount of secondary metabolites.
Various studies have shown that plant cell cultures, including both suspension cultures and hairy root cultures, have proven to be an extremely useful platform for production of secondary metabolites and further research needs to be performed to develop fast growing; renewable and cheap source of material for production of economically viable, complex structured pharmaceutically important secondary metabolites.
REFERENCES
- Alikaridis, F., D. Papadakis, K. Pantelia and T. Kephalas, 2000. Flavonolignan production from Silybum marianum transformed and untransformed root cultures. Fitotrapia, 71: 379-384.
CrossRef - Anand, S., V.S. Muthusamy, S. Sujatha, K.N. Sangeetha and R. Bharathi Raja et al., 2010. Aloe emodin glycosides stimulates glucose transport and glycogen storage through PI3K dependent mechanism in L6 myotubes and inhibits adipocyte differentiation in 3T3L1 adipocytes. FEBS Lett., 584: 3170-3178.
CrossRef - Ayadi, R. and J. Tremouillaux-Guiller, 2003. Root formation from transgenic calli of Ginkgo biloba. Tree Physiol., 23: 713-718.
Direct Link - Bais, H.P., R. Vepachedu and J.M. Vivanco, 2003. Root specific elicitation and exudation of fluorescent [β]-carbolines in transformed root cultures of Oxalis tuberosa. Plant Physiol. Biochem., 41: 345-353.
CrossRef - Balandrin, M.F., J.A. Klocke, E.S. Wurtele and W.H. Bollinger, 1985. Natural plant chemicals: Sources of industrial and medicinal materials. Science, 228: 1154-1160.
CrossRefPubMedDirect Link - Benhamou, N., 1996. Elicitor-induced plant defence pathways. Trends Plant Sci., 1: 233-240.
CrossRefDirect Link - Brodelius, P., 1988. Permeabilization of plant cells for release of intracellulary stored products: Viability studies. Applied Microbial. Biotechnol., 27: 561-566.
Direct Link - Brodelius, P., B. Deus, K. Mosbach and M.H. Zenk, 1979. Immobilized plant cells for the production of natural products. FEBS. Lett., 103: 93-97.
PubMed - Broeckling, C.D., D.V. Huhman, M.A. Farag, J.T. Smith and G.D. May et al., 2005. Metabolic profiling of Medicago truncatula cell cultures reveals the effects of biotic and abiotic elicitors on metabolism. J. Exp. Bot., 56: 323-336.
PubMed - Chaudhuri, K., S. Das, M. Bandyopadhyay, A. Zalar, A. Kollmann, S. Jha and D. Tepfer, 2009. Transgenic mimicry of pathogen attack stimulates growth and secondary metabolite accumulation. Transgenic Res., 18: 121-134.
CrossRef - Akimoto-Tomiyama, C., H. Aoyagi, T. Ozawa and H. Tanaka, 2002. Production of 5'-phosphodiesterase by Catharanthus roseus cells promoted by heat-degraded products generated from uronic acid. J. Biosci. Bioeng., 94: 154-159.
CrossRef - Collins-Pavao, M., C.K. Chin and H. Pedersen, 1996. Taxol partitioning in two-phase plant cell cultures of Taxus brevifolia. J. Biotechnol., 49: 95-100.
CrossRef - Corcoran, O. and M. Spraul, 2003. LC-NMR-MS in drug discovery. Drug Discovery Today, 8: 624-631.
CrossRef - Courtois, D. and J. Guern, 1980. Temperature response of Catharanthus roseus cells cultivated in liquid medium. Plant Sci. Lett., 17: 473-482.
CrossRef - DiCosmo, F. and M. Misawa, 1985. Eliciting secondary metabolism in plant cell cultures. Trends Biotechnol., 3: 318-322.
CrossRef - Dietrich, A., J.E. Mayer and K. Hahlbrock, 1990. Fungal elicitor triggers rapid, transient, and specific protein phosphorylation in parsley cell suspension cultures. J. Biol. Chem., 265: 6360-6368.
Direct Link - Doke, N., Y. Miura, L.M. Sanchez, H.J. Park, T. Noritake, H. Yoshioka and K. Kawakita, 1996. The oxidative burst protects plants against pathogen attack: Mechanism and role as an emergency signal for plant bio-defence: A review. Gene, 179: 45-51.
CrossRefPubMedDirect Link - Dornenburg, H., 2008. Plant cell culture technology-harnessing a biological approach for competitive cyclotides production. Biotechnol. Lett., 30: 1311-1321.
PubMed - Dornenburg, H. and P. Seydel, 2008. Effect of irradiation intensity on cell growth and kalata B1 accumulation in Oldenlandia affinis cultures. Plant Cell Tiss. Org. Cult., 92: 93-99.
CrossRef - Dueckershoff, K., M. Unger, A. Frank, E.M. Gillam, F.P. Guengerich and H. Warzecha, 2005. Modified nicotine metabolism in transgenic tobacco plants expressing the human cytochrome P450 2A6 cDNA. FEBS. Lett., 579: 2480-2484.
PubMed - Eliasova, A., M. Repcak and A. Pastirova, 2004. Quantitative changes of secondary metabolites of Matricaria chamomilla by abiotic stress. Zeitschrift Naturforschung, 59: 543-548.
Direct Link - Ellis, B.E. and G.H.N. Towers, 1970. Biogenesis of rosamrinic acid in Mentha. J. Biochem., 118: 291-297.
Direct Link - Fabricant, D.S. and N.R. Farnsworth, 2001. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect., 109: 69-75.
CrossRefPubMedDirect Link - Felix, G., M. Regenass, P. Spanu and T. Boller, 1994. The protein phosphatase inhibitor calyculin A mimics elicitor action in plant cells and induces rapid hyperphosphorylation of specific proteins as revealed by pulse labeling with [33p] phosphate. Proc. Natl. Acad. Sci., 91: 952-956.
Direct Link - Fett-Neto, A.G., S.J. Melanson, K. Sakata and F. Di-Cosmo, 1993. Improved growth and taxol yield in developing calli of Taxus cuspidata by medium composition modification. Nat. Biotechnol., 11: 731-734.
Direct Link - Fujita, Y., Y. Hara, T. Ogino and C. Suga, 1981. Production of shikonin derivatives by cell suspension cultures of Lithospermum erythrorhizon. Plant Cell Rep., 1: 59-60.
CrossRef - Fukui, H., N. Yoshikawa and M. Tabata, 1983. Induction of shikonin formation by agar in Lithospermum erythrorhizon cell suspension cultures. Phytochemistry, 22: 2451-2453.
CrossRef - Gabriel, D.W. and B.G. Rolfe, 1990. Working models of specific recognition in plant-microbe interactions. Annu. Rev. Phytopathol., 28: 365-391.
CrossRef - Ganesan, A., 2002. Recent developments in combinatorial organic synthesis. Drug Discovery Today, 7: 47-55.
PubMed - Garcia-Brugger, A., O. Lamotte, E. Vandelle, S. Bourque and D. Lecourieux et al., 2006. Early signaling events induced by elicitors of plant defenses. MPMI, 19: 711-724.
PubMedDirect Link - Gayathri, B., N. Manjula, K.S. Vinaykumar, B.S. Lakshmi and A. Balakrishnan, 2007. Pure compound from Boswellia serrata extract exhibits anti-inflammatory property in human PBMCs and mouse macrophages through inhibition of TNF alpha IL-1 beta NO and MAP kinases. Int. Immunopharmacol., 7: 473-482.
CrossRef - Goossens, A., S.T. Hakkinen, I. Laakso, T. Seppanen-Laakso and S. Biondi et al., 2003. A functional genomics approach toward the understanding of secondary metabolism in plant cells. Proc. Natl. Acad. Sci. USA., 100: 8595-8600.
Direct Link - Grzegorczyk, I. and H. Wysokinska, 2008. Liquid shoot culture of Salvia officinalis L. for micropropagation and production of antioxidant compounds: Effect of triacontanol. Acta Soc. Botanicorum Poloniae, 73: 99-104.
Direct Link - Guillon, S., J. Tremouillaux-Guiller, P.K. Pati, M. Ridean and P. Gantet, 2006. Hairy root research: Recent scenario and exciting prospects. Curr. Opin. Plant Biol., 93: 341-346.
CrossRefPubMedDirect Link - Hagimori, M., T. Matsumoto and Y. Obi, 1982. Studies on the production of Digitalis purpurea cardenolides by plant tissue culture. Plant Physiol., 69: 653-656.
Direct Link - Hahn, M.G., 1996. Microbial elicitors and their receptors in plants. Annu. Rev. Phytopathol., 34: 387-412.
CrossRefPubMedDirect Link - Hamada, H., K. Sanada, T. Furuya, S. Kawabe and M. Jaziri, 1996. Biotransformation of paclitaxel by cell suspension cultures of Eucalyptus perriniana. Nat. Prod. Lett., 9: 47-52.
CrossRef - Hammond-Kosack, K.E. and J.D.G. Jones, 1996. Resistance gene-dependent plant defense responses. Plant Cell, 8: 1773-1791.
CrossRefDirect Link - Hiraoka, N., I.D. Bhatt, Y. Sakurai and J.I. Chang, 2004. Alkaloid production by somatic embryo cultures of Corydalis ambigua. Plant Biotechnol., 21: 361-366.
Direct Link - Hohtola, A., J. Jalonen, A. Tolnen, L. Jaakola and T. Kamarainen et al., 2005. Natural Product Formation by Plants, Enhancement, Analysis, Processing and Testing. In: Sustainable Use Renewable Natural Resources from Principles to Practices, Jalkanen, A. and P. Nygren, (Eds.). University of Helsinki Publication, United Kingdom.
- Hong, S.B., C.A. Peebles, J.V. Shanks, K.Y. San and S.I. Gibson, 2006. Expression of the arabidopsis feedback-insensitive anthranilate synthase holoenzyme and tryptophan decarboxylase genes in Catharanthus roseus hairy roots. J. Biotechnol., 122: 28-38.
PubMed - Hoshino, T., M. Chida, T. Yamaura, Y. Yoshizawa and J. Mizuatani, 1994. Phytoalexin induction in green pepper cultures treated with arachidonic acid. Phytochemistry, 36: 1417-1419.
CrossRef - Ikeda, T., T. Matsumoto and M. Noguchi, 1977. Effects of inorganic nitrogen source and physical factors on the formation of ubiquinone by tobacco plant cells in suspension culture. Agric. Biol. Chem., 41: 1197-1201.
Direct Link - Inui, T., K. Tamura, N. Fujii, T. Murishige and F. Sato, 2007. Overexpression of Coptis japonica norcoclaurine 6-Omethyltransferase overcomes the rate-limiting step in Benzylisoquinoline alkaloid biosynthesis in cultured Eschscholzia californica. Plant Cell Physiol., 48: 252-262.
CrossRef - Ishida, B.K., 1988. Improved diosgenin production in Dioscorea deltoidea cell cultures by immobilized in polyurethane foam. Plant Cell Rep., 7: 270-273.
CrossRef - Jachak, S.M. and A. Saklani, 2007. Challenges and opportunities in drug discovery from plants. Curr. Sci., 92: 1251-1257.
Direct Link - Karuppusamy, S., 2009. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res., 3: 1222-1239.
CrossRefDirect Link - Kilby, N.J. and C.S. Hunter, 1990. Towards optimisation of the use of 1.02-MHz ultrasound to harvest vacuole-located secondary product from in vitro grown plant cells. Applied Microbiol. Biotechnol., 34: 478-480.
CrossRef - Kim, H.J., F. Chen, X. Wang and N.C. Rajapakse, 2005. Effect of chitosan on the biological properties of sweet basil (Ocimum basilicum L.). J. Agric. Food Chem., 53: 3696-3701.
CrossRefPubMedDirect Link - Knorr, D. and J.J. Berlin, 1987. Effects of immobilization and permeabilization procedures on growth of Chenopodium rubrum cells. J. Food Sci., 52: 1397-1400.
Direct Link - Kobayashi, Y., H. Fukui and M. Tabata, 1987. An immobilized cell culture system for berberine production by Thalictrum minus cells. Plant Cell Rep., 6: 185-186.
CrossRef - Kolewe, M.E., V. Gaurav and S.C. Roberts, 2008. Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol. Pharm., 5: 243-256.
PubMed - Komaraiah, P., G.V. Reddy, P.S. Reddy, A.S. Raghavendra, S.V. Ramakrishna and P. Reddanna, 2003. Enhanced production of antimicrobial sesquiterpenes and lipoxygenase metabolites in elicitor-treated hairy root cultures of Solanum tuberosum. Biotechnol. Lett., 25: 593-597.
CrossRef - Kotchoni, S.O. and E.W. Gachomo, 2006. The reactive oxygen species network pathways: An essential prerequisite for perception of pathogen attack and the acquired disease resistance in plants. J. Biosci., 31: 389-404.
PubMed - Krolicka, A., R. Kartanowicz, S.A. Wosinski, A. Szpitter, M. Kaminski and E. Lojkowska, 2006. Induction of secondary metabolite production in transformed callus of Ammi majus L. grown after electromagnetic treatment of the culture medium. Enzyme Microbial Technol., 39: 1386-1391.
CrossRef - Kuroyanagi, M., T. Arakava, Y. Mikami, K. Yoshida, N. Kawahar, T. Hayashi and H. Ishimaru, 1998. Phytoalexins from hairy root culture of Hyoscyamus albus treated with methyl jasmonate. J. Nat. Prod., 61: 1516-1519.
CrossRef - Kusakari, K., M. Yokoyama and S. Inomata, 2000. Enhanced production of saikosaponins by root culture of Bupleurum falcatum L. using two step control of sugar concentration. Plant Cell Rep., 19: 1115-1120.
CrossRef - Lakshmi, B.S., S. Sujatha, S. Anand, K.N. Sangeetha and R.B. Narayanan et al., 2009. Cinnamic acid, from the bark of Cinnamomum cassia, regulates glucose transport via activation of GLUT4 on L6 myotubes in a phosphatidylinositol 3-kinase-independent manner. J. Diabetes, 1: 99-106.
CrossRef - Laxalt, A.M., N. Raho, A.T. Have and L. Lamattina, 2007. Nitric oxide is critical for inducing phosphatidic acid accumulation in xylanase-elicited tomato Cells. J. Biol. Chem., 282: 21160-21168.
Direct Link - Flem-Bonhomme, V.L., D. Laurain-Mattar and M.A. Fliniaux, 2004. Hairy root induction of Papaver somniferum var. album, a difficult-totransform plant, by A. rhizogenes LBA 9402. Planta, 218: 890-893.
CrossRef - Lee, T.K., T. Yamakawa, T. Kodama and K. Shimomura, 1998. Effects of chemicals on alkaloid production by transformed roots of Atropa belladona. Phytochemistry, 49: 2343-2347.
CrossRef - Levine, A., R. Tenhaken, R. Dixon and C. Lamb, 1994. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell, 79: 583-593.
CrossRefPubMedDirect Link - Lin, H.W., K.H. Kwok and P.M. Doran, 2003. Development of Linum flavum hairy root cultures for production of coniferin. Biotechnol. Lett., 25: 521-525.
CrossRef - Lindsey, K. and M.M. Yeoman, 1984. The synthetic potential of immobilised cells of Capsicum frutescens Mill. cv. Annuum. Planta, 162: 495-501.
CrossRefDirect Link - Liu, R., Y. Hu, J. Li and Z. Lin, 2007. Production of soybean isoflavone genistein in non-legume plants via genetically modified secondary metabolism pathway. Metab. Eng., 9: 1-7.
CrossRef - Lorence, A., F. Medina-Bolivar and C.L. Nessler, 2004. Camptothecin and 10-hydroxycamptothecin from Camptotheca acuminate hairy roots. Plant Cell Rep., 22: 437-441.
CrossRef - Low, P.S. and J.R. Merida, 1996. The oxidative burst in plant defense: Function and signal transduction. Physiol. Plant, 96: 533-542.
CrossRef - Mac-Carthy, J.J., D.R. Ratcliff and H.E. Street, 1980. The effect of nutrient medium composition on the growth cycle of Catharanthus roseus G. Don cells grown in batch culture. J. Exp. Bot., 31: 1315-1325.
Direct Link - Mackintosh, C., G.D. Lyon and R.W. Mackintosh, 1994. Protein phosphatase inhibitors activate anti-fungal defense responses of soybean cotyledons and cell cultures. Plant J., 5: 137-147.
Direct Link - Mader, C.J., 1999. Effects of jasmonic acid, silver nitrate and L-AOPP on the distribution of free and conjugated polyamines in roots and shoots of Solanum tuberosum, in vitro. J. Plant Physiol., 154: 79-88.
Direct Link - Malamy, J., J.P. Carr, D.F. Kessig and I. Raskin, 1990. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science, 250: 1002-1004.
CrossRefDirect Link - Manickavelu, A., K. Kawaura, K. Oishi, T. Shin-I and Y. kohara et al., 2010. Comparative gene expression analysis of susceptible and resistant near-isogenic lines in common wheat infected by Puccinia triticina. DNA Res.
CrossRef - Mc-Donald, K.A. and A.P. Jackman, 1989. Bioreactor studies of growth and nutrient utilization in Alfalfa suspension cultures. Plant Cell. Rep., 8: 455-458.
CrossRef - Memelink, J., J.W. Kijne, R.V.D. Heijden and R. Verpoorte, 2001. Genetic modification of plant secondary metabolite pathways using transcriptional regulators. Adv. Biochem. Eng. Biotechnol., 72: 103-125.
Direct Link - Meyer, H.P., A. Kiener, R. Imwinkelried and N. Shaw, 1997. Biotransformations for fine chemical production. CHIMIA. Int. J. Chemis., 51: 287-289.
Direct Link - Moyano, E., K. Jouhikainen, P. Tammela, J. Palazon and R.M. Cusido et al., 2003. Effect of pmt gene overexpression on tropane alkaloid production in transformed root cultures of Datura metel and Hyoscyamus muticus. J. Exp. Bot., 54: 203-211.
Direct Link - Mukherjee, P.K., S. Rai, V. Kumar, K. Mukherjee, P.J. Hylands and R.C. Hider, 2007. Plants of Indian origin in drug discovery. Expert Opin. Drug Discovery, 2: 633-657.
CrossRef - Muller, S.S., F. Kurosaki and A. Nishi, 1994. Role of salicylic acid and intrercelluar Ca2+ in the induction of chitinase activity in carrot suspension culture. Physiol. Mol. Plant Pathol., 45: 101-109.
CrossRef - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Muthusamy, V.S., C. Saravanababu, M. Ramanathan, R. Bharathi Raja, S. Sudhagar, S. Anand and B.S. Lakshmi, 2010. Inhibition of protein tyrosine phosphatase 1B (PTP1B) and regulation of insulin signalling markers by caffeoyl derivatives of Chicory (Cichorium intybus) salad leaves. Br. J. Nutr., 6: 1-11.
PubMed - Muthusamy, V.S., S. Anand, K.N. Sangeetha, S. Sujatha, B. Arun and B.S. Lakshmi, 2008. Tannins present in Cichorium intybus enhance glucose uptake and inhibit adipogenesis in 3T3-L1 adipocytes through PTP1B inhibition. Chemico-Biol. Interact., 174: 69-78.
CrossRefPubMedDirect Link - Nakagawa, K., A. Konagai, H. Fukui and M. Tabata, 1984. Release and crystallization of berberine in the liquid medium of Thalictrum minus cell suspension cultures. Plant Cell Rep., 3: 254-257.
CrossRef - Namdeo, A.G., 2007. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev., 1: 69-79.
Direct Link - Namdeo, A., S. Patil and D.P. Fulzele, 2002. Influence of fungal elicitors on production of ajmalicine by cell cultures of Catharanthus roseus. Biotechnol. Prog., 18: 159-162.
Direct Link - Ogino, T., N. Hiraoka and M. Tabata, 1978. Selection of high nicotine producing cell lines of Tobacco callus by single cell cloning. Phytochemistry, 17: 1907-1910.
CrossRef - Orozco-Cardenas, M.L., J. Narvaez-Vasquez and C.A. Ryan, 2001. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin and methyl jasmonate. Plant Cell, 13: 179-191.
PubMed - Oubre, A.Y., T.J. Carlson, S.R. King and G.M. Reaven, 1997. From plant to patient: An ethnomedical approach to the identification of new drugs for the treatment of NIDDM. Diabetologia, 40: 614-617.
PubMed - Palazon, J., E. Moyano, R.M. Cusido, M. Bonfill, K.M.O. Caldentey and M.T. Pinol, 2003. Alkaloid production in duboisia hybrid hairy roots and plants overexpressing the h6h gene. Plant Sci., 165: 1289-1295.
CrossRef - Patwardhan, B., A.D.B. Vaidya and M. Chorghade, 2004. Ayurveda and natural products drug discovery. Curr. Sci., 86: 789-799.
Direct Link - Peebles, C.A., S.B. Hong, S.I. Gibson, J.V. Shanks and K.Y. San, 2005. Transient effects of overexpressing anthranilate synthase alpha and beta subunits in Catharanthus roseus hairy roots. Biotechnol. Prog., 21: 1572-1576.
PubMed - Pitzschke, A., A. Schikora and H. Hirt, 2009. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol., 12: 1-6.
Direct Link - Pras, N., H.J. Woerdenbag, S. Batterman, J.F. Visser and V.W. Uden, 1993. Mucuna pruriens: Improvement of the biotechnological production of the anti-Parkinson drug L-DOPA by plant cell selection. Pharm. World Sci., 15: 263-268.
PubMed - Radman, R., T. Saez, C. Bucke and T. Keshavarz, 2003. Elicitation of plant and microbial systems. Biotechnol. Applied Biochem., 37: 91-102.
PubMed - Rajendran, L., G.A. Ravishankar, L.V. Venkataraman and K.R. Prathiba, 1992. Anthocyanin production in callus cultures of Daucuscarota L. as influenced by nutrient stress and osmoticum. Biotechnol. Lett., 14: 707-712.
CrossRef - Rao, S.R. and G.A. Ravishankar, 2002. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv., 20: 101-153.
CrossRefPubMedDirect Link - Rao, S.R., U. Tripathi and G.A. Ravishankar, 2002. Biotransformation of digitoxin to digoxin in cell cultures of Capsicum frutescens in the presence of b-cyclodextrin. Biocatalysis Biotransformation, 20: 137-143.
CrossRef - Rao, S.R., U. Tripathi and G.A. Ravishankar, 1999. Biotransformation of codeine to morphine in free and immobilized cell cultures of Spirulina platensis. World J. Microbiol. Biotechnol., 15: 465-469.
CrossRef - Raskin, I. and C. Ripoll, 2004. Can an apple a day keep the doctor away? Curr. Pharm. Des., 10: 3419-3429.
CrossRefDirect Link - Ravishankar, G.A. and S.R. Rao, 2000. Biotechnological production of phyto-pharmaceuticals. J. Biochem. Mol. Biol. Biophys., 4: 73-102.
Direct Link - Sangeetha, K.N., S. Sujatha, V.S. Muthusamy, S. Anand and N. Nirmal et al., 2010. 3β-taraxerol of Mangifera indica, a PI3K dependent dual activator of glucose transport and glycogen synthesis in 3T3-L1 adipocytes. BBA-Gen. Subjects, 1800: 359-366.
CrossRef - Selvakkumar, C., A. Balakrishnan and B.S. Lakshmi, 2007. Rapid in vitro micropropagation of Alpinia officinarum hance, an important medicinal plant, through rhizome bud explants. Asian J. Plant Sci., 6: 1251-1255.
CrossRefDirect Link - Selvakkumar, C., B. Gayathri, K.S. Vinaykumar, B.S. Lakshmi and A. Balakrishnan, 2007. Potential Anti-inflammatory properties of crude alcoholic extract of Ocimum Basilicum L. in human peripheral blood mononuclear cells. J. Health Sci., 53: 500-505.
Direct Link - Shilpa, K., K.N. Sangeetha, V.S. Muthusamy, S. Sujatha, B.S. Lakshmi, 2009. Probing key targets in insulin signaling and adipogenesis using methanolic extract of Costus pictus and its bioactive molecule methyl tetracosanoate. Biotechnol. Lett., 31: 1837-1841.
CrossRef - Shilpa, K., C. Selvakkumar, R.S.A. Kumar and B.S. Lakshmi, 2010. In vitro root culture of Ocimum sanctum L. and evaluation of its free radical scavenging activity. Plant Cell Tissue Organ Cult., 101: 105-109.
CrossRefDirect Link - Skrzypek, Z. and H. Wysokinsku, 2003. Sterols and triterpenes in cell cultures of Hyssopus officinalis L. Z. Naturforsch, 58: 308-312.
PubMed - Smith, M.A.L., H. Kobayashi, M. Gawienowski and D.P. Briskin, 2002. An in vitro approach to investigate chemical synthesis by three herbal plants. Plant Cell Tissue Organ Cult., 70: 105-111.
CrossRef - Steinbeck, C., 2004. Recent development in automated structure elucidation of natural products. Nat. Prod. Rep., 21: 512-518.
PubMed - Subramanian, K., C. Selvakkumar, S. Meenakshisundaram, A. Balakrishnan and B.S. Lakshmi, 2008. Extract of Alpinia officinarum suppresses enteropathogenic Escherichia coli (EPEC) Lipopolysaccharide (LPS) induced inflammation in J774 A.1 Macrophages. J. Health Sci., 54: 112-117.
Direct Link - Sudha, C.G., B.O. Reddy, G.A. Ravishankar and S. Seeni, 2003. Production of ajmalicine and ajmaline in hairy root cultures of Rauvolfia micrantha Hook f., a rare and endemic medicinal plant. Biotechnol. Lett., 25: 631-636.
PubMed - Sujatha, S., S. Anand, K. N. Sangeetha, K. Shilpa, J. Lakshmi, A. Balakrishnan, B.S. Lakshmi, 2009. Biological evaluation of (3β)-stigmast-5-en-3-ol as potent anti-diabetic agent in regulating glucose transport using in vitro model. Int. J. Dibetes Mel.
CrossRef - Tian, L. and R.A. Dixon, 2006. Engineering isoflavone metabolism with an artificial bifunctional enzyme. Planta, 224: 496-507.
CrossRef - Van-Uden, W., A.S. Bouma, J.F.S. Waker, O. Middel and H.J. Wichers et al., 1995. The production of podophyllotoxin and its 5-methoxy derivatives through bioconversion of cyclodextrin complexed desoxypodophyllotoxin by plant cell cultures. Plant Cell Tissue Org. Cult., 42: 73-79.
CrossRef - Ciddi, V., V. Srinivasan and M.L. Shuler, 1995. Elicitation of Taxus Sp. cell cultures for production of taxol. Biotechnol. Lett., 17: 1343-1346.
CrossRef - West-jr, F.R. and E.S. Mika, 1957. Synthesis of atropine by isolated roots and root-callus cultures of Belladonna. Bot. Gaz., 119: 50-54.
Direct Link - Wilhelm, R. and M.H. Zenk, 1997. Biotransformation of thebaine by cell cultures of Papaver somniferum and Mohonia nervosa. Phytochemicals, 46: 701-708.
CrossRef - Yamada, Y. and F. Sato, 1981. Production of berberine in cultured cells of Coptis japonica. Phytochemistry, 20: 545-547.
CrossRef - Yamakawa, T., S. Kato, K. Ishida, T. Kodama and Y. Minoda, 1983. Production of anthocyanins by Vitis cells in suspension culture. Agric. Biol. Chem., 47: 2185-2191.
Direct Link - Yamamoto, H. and M. Yamamoto, 2000. Effects of various culture conditions on callus growth and polyphenol contents of Hydrangea macrophylla var. Thunbergii Callus cultures. Bull. Hokuriku Univ., 24: 25-33.
Direct Link - Yamamoto, Y., R. Mizuguchi and Y. Yamada, 1982. Selection of a high stable pigment producing strain in cultured Euphorbia milli cells. Theor. Applied Genet., 61: 113-116.
CrossRef - Yang, Y., J. Shah and D.F. Klessig, 1997. Signal perception and transduction in plant defense responses. Genes Dev., 11: 1621-1639.
PubMed - Yaoya, S., H. Kanho, Y. Mikami, T. Itani, K. Umehara and M. Kuroyanagi, 2004. Umbelliferone released from hairy root cultures of Pharbitis nil treated with copper sulfate and its subsequent glucosylation. Biosci. Biotechnol. Biochem., 68: 1837-1841.
PubMed - Yazaki, K., A. Sugiyama, M. Morita and N. Shitan, 2008. Secondary transport as an efficient membrane transport mechanism for plant secondary metabolites. Phytochem. Rev., 7: 513-524.
CrossRefDirect Link - Yeoman, M.M. and C.L. Yeoman, 1996. Tansley review no 90. Manipulating secondary metabolism in cultured plant cells. New Phytol., 134: 553-569.
Direct Link - Yoda, H., Y. Yamaguchi and H. Sano, 2003. Induction of hypersensitive cell death by hydrogen peroxide produced through polyamine degradation in tobacco plants. Plant Physiol., 132: 1973-1981.
Direct Link - Zenk, M.H., H. EI-Shagi and U. Schulte, 1975. Anthraquinone production by cell suspension cultures of Morinda citrifolia. Planta Med., 1975: 79-101.
PubMed - Zhao, D., C. Fu, Y. Chen and F. Ma, 2004. Transformation of Saussurea medusa for hairy roots and jaceosidin production. Plant Cell Rep., 23: 468-474.
CrossRef - Zhao, J., L.C. Davis and R. Verpoorte, 2005. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv., 23: 283-333.
CrossRefPubMedDirect Link - Zhong, J.J., 2001. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. Adv. Biochem. Eng. Biotechnol., 72: 1-26.
PubMed - Zhu, Q., W. Droge-Laser, R.A. Dixon and C. Lamb, 1996. Transcriptional activation of plant defense genes. Curr. Opin. Genet. Dev., 6: 624-630.
PubMed - Johnson, T.S., G.A. Ravishankar and L.V. Venkataraman, 1991. Elicitation of capsaicin production in freely suspended cells and immobilized cell cultures of Capsicum frutescens mill. Food Biotechnol., 5: 197-205.
CrossRefDirect Link