
J. W. Muthomi
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
G. M. Riungu
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J. K. Ndung`u
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
R. D. Narla
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J. K. Gathumbi
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
J. M. Wagacha
Department of Plant Science and Crop Protection, University of Nairobi, P.O. Box 30197, 00100 GPO, Nairobi, Kenya
Journal of Plant Sciences
Year: 2008 | Volume: 3 | Issue: 1 | Page No.: 52-60







ABSTRACT
The study was carried out during the 2006 cropping season in Nakuru district, Kenya. Incidence and severity of head blight were determined and pathogens isolated from diseased wheat heads, wheat and maize kernels. Mycotoxin deoxynivalenol content in grain was determined by direct competitive Enzyme-Linked Immunosorbent Assay (ELISA). Pathogenicity of different Fusarium species isolated from wheat was determined by inoculation onto wheat ears in greenhouse. Head blight was highly prevalent (90-100%) and mean incidence and severity ranged from 4 to 9% and 15 to 37%, respectively. Fusarium was most prevalent in infected wheat heads while Epicoccum was most prevalent in harvested wheat grain. Only Fusarium spp. and Penicillium spp. contaminated harvested maize grain. The most frequently isolated Fusarium species were F. poae, F. graminearum and F. chlamydosporum in wheat and F. verticilloides in maize. Most wheat and maize grain samples were contaminated with mycotoxin (DON), with concentration ranging from 0-1,200 and 0-4,600 µg kg-1, respectively. Fusarium graminearum isolates were highly pathogenic, significantly reducing kernel weight. The results suggest that head blight in Nakuru district is due to a complex of Fusarium species with F. graminearum being the major pathogen. Cross-contamination of wheat and maize is implied, indicating possible contamination of wheat maize products with deoxynivalenol mycotoxin.
PDF Abstract XML References Citation
How to cite this article
J. W. Muthomi, G. M. Riungu, J. K. Ndung`u, R. D. Narla, J. K. Gathumbi and J. M. Wagacha, 2008. Head Blight of Wheat in Kenya and Contamination of Grain with Mycotoxin Producing Fusarium Species. Journal of Plant Sciences, 3: 52-60.
DOI: 10.3923/jps.2008.52.60
URL: https://scialert.net/abstract/?doi=jps.2008.52.60
DOI: 10.3923/jps.2008.52.60
URL: https://scialert.net/abstract/?doi=jps.2008.52.60
INTRODUCTION
Fusarium head blight (FHB, scab) is a significant disease of small-grain cereals throughout the world where wheat is grown. Up to 17 causal organisms have been associated with the disease (Parry et al., 1995). Fusaria like F. avenaceum, F. culmorum, F. graminearum, F. poae (Kriel, 2006) and Microdochium nivale are the species most associated with FHB (Simpson et al., 2004). These fungi rarely exist in isolation, but occur as a complex with each other and with other Fusaria and other fungal genera. Climatic conditions will influence competition among and the predominance of different fungi within this complex (Doohan et al., 2003). Similar Fusaria species to those causing FHB are thought to cause ear rot of maize. However, three species of Fusarium are predominantly responsible for the disease: F. graminearum, F. verticillioides (moniliforme), F. subglutinans (Reid et al., 1999; Fandohan, 2005). Fusarium verticillioides is likely to be the most common species isolated worldwide from diseased maize (Logrieco et al., 2002; Oren et al., 2003).
Fusarium head blight may be epidemic over large areas in some seasons but more commonly varies in severity from field to field or between local areas. The prevalence of infection may vary from a trace to virtually 100% of the heads in the field and losses vary correspondingly (Brennan et al., 2007). Depending on the species involved Fusarium grain infection may result in yield reductions (Henriksen and Ellen, 2005). The extent to which yield is affected is influenced by the Fusarium species involved. In inoculation trials with F. graminearum on Kenyan varieties the yield was reduced by between 23 and 57% and head susceptibility of between 29 and 68% (Muthomi, 2001). Fusarium infections can also lead to mycotoxin contamination of the grains thus threatening the health of humans and livestock (Dohlman, 2004; Heier et al., 2005). Most frequently found mycotoxins are Deoxynivalenol (DON, also known as vomitoxin) and zearalenone (ZEA) (Llorens et al., 2006; Goyarts et al., 2007).
Earlier studies in Kenya (Muthomi, 2001; Muthomi et al., 2007a; Ndungu, 2006) have found different levels of Fusarium and mycotoxin contamination in wheat grains from different parts of the country. There is however, no documentation of the actual spread and the severity of the disease in farmer fields and the DON contamination in grain from different agro-ecological zones. Therefore, this study was carried out to determine the incidence and severity of FHB in farmers’ fields in different agro-ecological zones of Nakuru district and to determine the major species involved in infected wheat ears and grains of both wheat and maize.
MATERIALS AND METHODS
Determination of Incidence and Severity of Head Blight in Farmers’ Fields
The study was conducted during 2006 cropping season (May-December) in Nakuru district of Kenya. Five agro-ecological zones in which wheat is commonly grown in Nakuru district were considered: Upper midland 4 (UM4), Lower highland 2 (LH2), Upper highland 2 (UH2), Lower highland 3 (LH3) and Lower highland 4 (LH4). Ten farmers were systematically sampled from each of the five agro-ecological zones. Both large and small-scale farms were targeted. Head blight incidence and severity were determined at soft dough stage (GS85, Zadoks et al., 1974). Incidence was determined by marking out a 10 m2 quadrants and counting the number of blighted heads in the quadrant out of the total number of heads. Three quadrants were assessed per farm. Disease severity was measured as the average proportion of the heads that were blighted and was determined using the scale by Miedaner et al. (1996): -1% = no symptoms, 2 = <5%, 3 = 5-15%, 4 = 16-25%, 5 = 26-45%, 6 = 46-65%, 7 = 66-85%, 8 = 86-95%, 9 = 96-100%. Diseased heads were collected for isolation of causal agents. At harvest, 1-2 kg of the freshly harvested wheat and maize kernels were collected for mycological and mycotoxin analysis. The samples were stored at 4°C until analyzed.
Isolation and Identification of Fusarium species
The infected wheat heads were cut into small pieces (0.5 cm long). For the kernels, sub-samples were taken randomly. These were sterilized in 3% sodium hypochlorite for three minutes and rinsed off thrice in sterile distilled water. Plating was on low strength potato dextrose agar amended with mineral salts and antimicrobial agents (Muthomi, 2001): (PDA 17 g, KH2PO4 1.0 g, KNO3 1.0 g, MgSO4 0.5 g, Agar 10 g). Antibiotics penicillin, tetracycline, streptomycin and pentachloronitrobenzene (PCNB) were added (50 mg) after the media cooled to 45°C. A total of 100 kernels and 50 pieces for the diseased heads were plated per farm. Incubation was at 25°C for 5 to 14 days under 12 h daylight and 12 h darkness cycles.
Fungal colonies were identified based on cultural and morphological characteristics like mycelial colour, pigmentation, spore shape, septation and sporophores. Fusarium colonies were sub-cultured onto PDA and Synthetic Nutrient Agar (SNA) (Nirenberg, 1981): (KH2PO4 1.0 g, KNO3 1.0 g, MgSO4 0.5 g, KCl 0.5 g, Glucose 0.2 g, Agar 20 g) and incubated under-near UV light for seven days to induce sporulation. The cultures were then identified to species level according to Nelson et al. (1983) and Seifert (1996).
Determination of Deoxynivalenol Content in Grains
Deoxynivalenol content in the wheat and maize grain was analyzed by direct competitive Enzyme-Linked Immunosorbent Assay (ELISA), (AOAC, 1995; Gathumbi et al., 2001). Each sample was homogenized and 100 g ground to fine powder. Five grams of the ground sample was extracted with 25 mL of methanol:water (50:50v/v). The extract was de-fatted with 10 mL hexane and 4 mL of the methanolic layer taken and diluted to 10% using Phosphate Buffer Solution (PBS). Microtitre polystyrene plates were coated with 100 μL of anti-deoxynivalenol antiserum DON143/16 (Usleber et al., 1992) in bicarbonate buffer (pH 9.6) per well. Absorbance was determined using spectrophotometer Elisa reader at 450 nm. A calibration curve for the standards for each toxin dilutions were plotted using log10 of standards concentration against the percentage inhibition of the standards.
Pathogenicity of Fusarium Species
Isolates of different Fusarium species from wheat kernels were inoculated onto spikes of ‘Mbuni’, a highly susceptible wheat cultivar (Muthomi et al., 2002) under greenhouse conditions. Each isolate was cultured separately at 22±5°C on mung bean medium (Bai and Shaner, 1996) for 14 days. Conidia suspension of each isolate was harvested and adjusted to 5x105 conidia mL-1. Three drops (0.01%) of Tween 20 was added to ensure uniform conidia dispersion. Wheat ears were inoculated at 50% flowering (GS65, Zadoks et al., 1974) by spraying with hand sprayer, exposing all spikelets to the inoculum. Controls were treated similarly with distilled water. After inoculation, the ears were incubated under polythene bags for 48 h to ensure high relative humidity for optimal infection. Each isolate was inoculated separately and replicated four times. Head blight severity was visually assessed using a 1-9 scale (Miedaner et al., 1996) as proportion of bleached spikelets after every 5 days on 10 average sized ears per replicate.
Data Analysis
All data were subjected to analysis of variance (ANOVA) using the PROC ANOVA procedure of Genstat (Lawes Agricultural Trust Rothamsted Experimental station 2006, version 9) and differences among the treatments means were compared using the Fisher’s protected LSD test at 5% probability level.
RESULTS
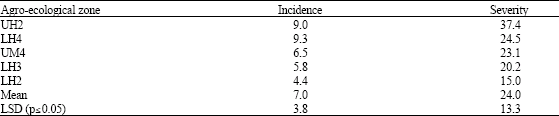
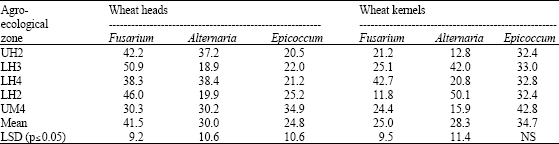
Fusarium head blight was found in most farms (97%) but there were significant differences (p≤0.05) among the agro-ecological zones in both head blight incidence and severity (Table 1). Fusarium, Alternaria and Epicoccum were isolated from both diseased heads and wheat kernels (Table 2). However, Fusarium was the major contaminant in the heads (41%) while Epicoccum was the main contaminant in wheat kernels (34%). There were significant differences (p≤0.05) among the agro-ecological zones. Only Fusarium and Penicillium were isolated from the maize kernels.
| Table 1: | Prevalence and severity of Fusarium head blight on wheat ears in farmer’s fields in different agro-ecological zones of Nakuru district |
 | |
| Table 2: | Percentage isolation frequency of major fungi contaminating wheat ear and kernels in different agro-ecological zones of Nakuru district |
 | |
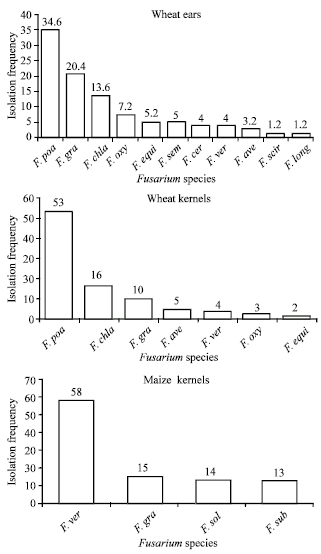 | |
| Fig. 1: | Fusarium species isolated from head blight infected wheat ears, harvested wheat kernels and harvested maize kernels from Nakuru district (F. gra = F. graminearum; F. poa = F. poae; F. chla = F. chlamydosporum; F. oxy = F. oxysporum; F. equi = F. equiseti; F. sem = F. semitectum; F. cer = F. cerealis; F. ver = F. verticillioides; F. ave = F. avenaceum; F. scir = F. scirpi; F. long = F. longipes; F. sub. F. subglutinans; F. sol = F. solani) |
The infected wheat heads, wheat and maize kernels differed in the spectrum of Fusarium species isolated (Fig. 1). However, the highest number of Fusarium species was isolated from the diseased wheat heads while maize kernels had the least number of species isolated.
| Table 3: | Deoxynivalenol contamination of wheat and maize grain from different agro-ecological zones of Nakuru district |
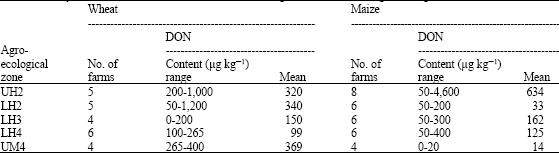 | |
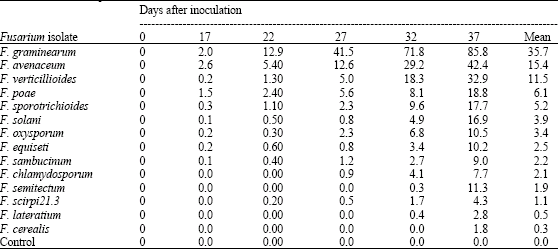
| Table 4: | Average disease severity over time on wheat ears inoculated with Fusarium species isolated from wheat kernels Fusarium species |
 | |
The most frequently isolated Fusarium species from wheat heads and kernels were F. poae, F. chlamydosporum, F. graminearum, F. avenaceum and F. equiseti. Fusarium verticilloides, F. graminearum, F. solani and F. subglutinans were most isolated from maize kernels. Fusarium graminearum was found to be a major contaminant in wheat heads, kernels and maize.
Most of the wheat and maize samples were contaminated with mycotoxin deoxynivalenol (DON) but the different agro-ecological zones differed the amount of the toxin (Table 3). Wheat samples had higher mean levels of DON (249 μg kg-1) compared to maize (mean 221 μg kg-1). However, the highest DON content was detected in maize samples with up to 4,600 μg kg-1 compared to 1,200 μg kg-1 in wheat. The different Fusarium species significantly differed (p≤0.05) in severity of head blight induced on inoculated wheat ears (Table 4). However, isolates of Fusarium graminearum were the most pathogenic, resulting in the highest disease severity of up to 87% of spikelets bleached.
DISCUSSION
Fusarium head blight was found in all the agro ecological zones surveyed (AEZS) with 100% prevalence. The disease severity was highly positively co-related to incidence (r = 0.647, p≤0.001). The varying severity and incidence over the different AEZS could be due to the different environmental conditions that impact on numerous aspects of FHB epidemiology (Doohan et al., 2003; Brennan et al., 2005; Chen et al., 2006; Klahr et al., 2007a). The major wheat varieties grown in Kenya have been found to be susceptible to FHB (Muthomi et al., 2002, 2007; Ndung'u, 2006) Currently, no source of complete resistance is known but varieties with partial resistance have been reported (Wisniewska and Kowalczk, 2005; Browne, 2007; Semagn et al., 2007).
Diseased wheat heads and kernels were contaminated with high levels of Alternaria and Epicoccum. Epicoccum and Alternaria species are mainly saprophytes or weak pathogens that grow on senescencing plant tissues (Vincent et al., 2006). However, Alternaria alternata is one of the most common wheat pathogens, causing huge reductions in yield worldwide (Williamson, 1997). The co-occurrence of several fungi could have synergistic effect on disease severity and higher reductions of wheat yield. In addition, some species of Alternaria are known to produce mycotoxins altanariol, altenuene, tenuazonic acid and altertoxin I-III (Weidenbörner, 2001; Sab et al., 2007). It was noted where the isolation frequency of Alternaria or Epicoccum was high that of Fusarium was low similar to results by Kosiak et al. (2004), Ndung’u (2006) and Sab et al. (2007).
Over 10 different Fusarium species were isolated from infected wheat ears and harvested kernels, indicating that head blight is due to a complex of Fusarium species. Species isolated at high frequencies were F. poae, F. oxysporum, F. graminearum and F. chlamydosporum. However, results from inoculation studies showed that only F. graminearum was highly pathogenic, suggesting that this species could be implicated in the possible head blight of wheat in Kenya. Fusarium graminearum is known to cause severe blighting of wheat heads resulting in visually damaged kernels (Brennan et al., 2005). Other species including Fusarium poae, F. avenaceum, F. equiseti and F. sporotrichioides do not blight the heads but sometimes cause damage to the spikelets, resulting in low disease severity and latent seed infection. According to Marasas (1991) Fusarium equiseti, F. graminearum, F. poae, F. verticillioides and F. sporotrichioides are considered the most toxic Fusarium species.
Although wheat and maize differed in the spectrum of Fusarium species isolated, both were contaminated with F. graminearum and F. verticilloides. Maize is the staple grain in Kenya and it is grown alongside wheat by most farmers. Wheat fields are surrounded by fields used for different crops, usually maize, sorghum or barley wheat are therefore potentially exposed to cross-contamination by other plants (Vincent et al., 2006). Fusarium graminearum the predominant causal agent of head blight (Kriel, 2006) is also a major pathogen in ear rot of maize (Parry et al., 1995; Silva et al., 2007). The co-occurrence of Fusarium species in maize and wheat kernels suggests co-occurrence of mycotoxins, F. graminearum and F. poae produce type B trichothecenes such as nivalenol, deoxynivalenol, zearalenone and Fusarenon-x (Geraldo et al., 2006; Kriel, 2006) whereas F. verticilloides produces fumonisins and moniliformin (Marasas, 2001; Ramos et al., 2006). Ndung’u (2006) reported co-occurrence of mycotoxins deoxynivalenol, zearalenone and aflatoxin B1 in harvested wheat in Kenya. Deoxynivalenol was the major mycotoxin while the rest were at low levels. These mycotoxins cause a wide range of acute and chronic effects in humans and animals (Tiemann and Daenicke, 2007). Sab et al. (2007) suggested a possible role played by zealarenone in competitive interactions between F. graminearum and A. alternata. Contamination of maize with fumonisins has been reported in Kenya (Kedera, 1999). Consumption of fumonisin has been associated with elevated human esophageal cancer incidence in various parts of Africa, Central America and Asia (Marasas et al., 2004). Some correlation studies have suggested a link between the consumption of maize with high incidence of F. verticillioides and fumonisins and the high incidence of human oesophageal carcinoma in certain parts of S. Africa (Yoshizawa et al., 1994; Anonymous, 2000). Fusarium verticilloides is also known to infect both maize and wheat (Dill-Macky and Jones, 2000; Ramos et al., 2006).
The study indicated that FHB is present in significant levels in Kenya and that it is due to a complex of different Fusarium species. However, F. graminearum and F. avenaceum could be main pathogenic species involved in pathogenicity. The presence of F. graminearum in both wheat and maize indicated that maize is a reservoir for the head blight pathogens. This could be due to the growing of wheat and maize in the same or neighbouring fields and lack of proper rotation programmes. Due to the health risks posed by mycotoxin contamination, there is need for continuous surveillance for the major Fusarium mycotoxins, especially deoxynivalenol to prevent carry over to food and animal feeds. Further characterization of F. graminearum isolates from maize and wheat would be necessary to determine whether similar population infect both crops. In addition, the resulting yield losses and possible management strategies for FHB need to be determined.
ACKNOWLEDGMENT
This study was funded by the International Foundation for Science (IFS) grant No.E/3654-1.
REFERENCES
- AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Bai, G.H. and G. Shaner, 1996. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis., 80: 975-979.
CrossRefDirect Link - Brennan, J.M., D. Egan, B.M. Cooke and F.M. Doohan, 2005. Effect of temperature on head blight of wheat caused by Fusarium culmorum and F. graminearum. Plant Pathol., 54: 156-160.
CrossRefDirect Link - Brennan, J.M., G. Leonard, B. Fagan, B.M. Cooke and A. Ritieni et al., 2007. Comparison of commercial European wheat cultivars to Fusarium infection of head and seedling tissue. Plant Pathol., 56: 55-64.
Direct Link - Browne, R.A., 2007. Components of resistance to Fusarium head blight (FHB) in wheat detected in a seed-germination assay with Microdochium majus and the relationship to FHB disease development and mycotoxin accumulation from Fusarium graminearum infection. Plant Pathol., 56: 65-72.
Direct Link - Chen, C.A., M.A. Griffey, M. Saghai, E.L. Stromberg and R.M. Biyashev et al., 2006. Validation of two major quantitative trait loci for Fusarium head blight resistance in Chinese wheat line W14. Plant Breed., 125: 99-101.
Direct Link - Dill-Macky, R. and R.K. Jones, 2000. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis., 84: 71-76.
CrossRefDirect Link - Doohan, F.M., J. Brennan and B.M. Cooke, 2003. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol., 109: 755-768.
Direct Link - Fandohan, P., B. Gnonlonfin, K. Hell, W.F.O. Marasas and M.J. Wingfield, 2005. Natural occurrence of Fusarium and subsequent fumonisin contamination in preharvest and stored maize in Benin, West Africa. Int. J. Food Microbiol., 99: 173-183.
Direct Link - Gathumbi, J.K., E. Usleber and E. Mrtlbauer, 2001. Production of ultrasensitive antibodies against aflatoxin B1. Lett. Applied Microbiol., 32: 349-351.
CrossRef - Geraldo, M.R.F., D.J. Tessmann and C. Kemmelmeier, 2006. Production of mycotoxins by Fusarium graminearum isolated from small cereals (wheat, triticale and barley) affected with scab disease in Southern Brazil. Brazilian J. Microbiol., 37: 58-63.
CrossRefDirect Link - Goyarts, T., S. Dänicke, H. Valenta and K.H. Ueberschär, 2007. Carry-over of Fusarium toxins (deoxynivalenol and zearalenone) from naturally contaminated wheat to pigs. Food Additives and Contaminants, 24: 369-380.
Direct Link - Heier, T., S.K. Jain, K.H. Kogel and J. Poris Kuhneman, 2005. Influence of N fertilization and fungicide strategies on Fusarium head blight severity and mycotoxin content in winter wheat. J. Phytopathol., 153: 551-557.
Direct Link - Henriksen, B. and O. Ellen, 2005. Natural Fusarium grain infection level in wheat, barley and oat after early application of fungicides and herbicides. J. Phytopathol., 153: 214-220.
Direct Link - Kedera, C.J., R.D. Plattner and A. Desjardins, 1999. Incidence of Fusarium spp. and levels of fumonisins B1 in maize in Western Kenya. Applied Environ. Microbiol., 65: 41-44.
PubMed - Klahr, A., Z.G. Gerhard, W. Gerhard and M. Volker, 2007. Effects of environment, disease progress, plant height and heading date on the detection of QTLs for resistance to Fusarium head blight in an European winter wheat cross. Euphytica, 154: 17-28.
Direct Link - Kosiak, B., M. Torp, E. Skjerve and B. Andersen, 2004. Alternaria and Fusarium in Norwegian grains of reduced quality-a matched pair sample study. Int. J. Food Microbiol., 93: 51-62.
CrossRef - Llorens, A., M.J. Hinojo, R. Mateo, M.T. Gonzalez-Jaen, F.M. Valle-Algarra, A. Logrieco and M. Jimenez, 2006. Characterization of Fusarium spp. isolates by PCR-RFLP analysis of the intergenic spacer region of the rRNA gene (rDNA). Int. J. Food Microbiol., 106: 287-306.
CrossRefPubMedDirect Link - Logrieco, A., G. Mule, A. Moretti and A. Bottalico, 2002. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol., 108: 597-609.
Direct Link - Marasas, W.F.O., 1991. Toxigenic Fusaria. In: Mycotoxins and Animal Foods, Smith, J.E. and R.S. Henderson (Eds.). Chapter 6, CRC Press, Boca Raton, FL., pp: 119-139.
Direct Link - Marasas, W.F.O., R.T. Riley, K.A. Hendricks, V.L. Stevens and T.W. Sadler et al., 2004. Fumonisins disrupt sphingolipid metabolism, folate transport and neural tube development in embryo culture and in vivo: A potential risk factor for human neural tube defects among populations consuming fumonisin-contaminated maize. J. Nutr., 134: 711-716.
Direct Link - Miedaner, T., G. Gang and H.H. Geiger, 1996. Quantitative-genetic basis of aggressiveness of 42 isolates of Fusarium culmorum for winter rye head blight. Plant Dis., 80: 500-504.
Direct Link - Muthomi, J.W., E.C. Oerke, H.W. Dehne and E.W. Mutitu, 2002. Susceptibility of Kenyan wheat varieties to head blight, fungal invasion and deoxynivalenol accumulation inoculated with Fusarium graminearum. J. Phytopathol., 150: 30-36.
CrossRefDirect Link - Muthomi, J.W., J.K. Ndung`u , G.N. Chemining`wa and J.M. Wagacha, 2007. Reaction of some Kenyan wheat cultivars to head blight after inoculation with Fusarium graminearum. Asian J. Plant Sci., 6: 585-591.
CrossRefDirect Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Nirenberg, H.I., 1981. A simplified method for identifying Fusarium spp. occurring on wheat. Can. J. Bot., 59: 1599-1609.
CrossRefDirect Link - Oren, L., S. Ezrati, D. Cohen and A. Sharon, 2003. Early events in the Fusarium verticillioides-maize interaction characterized by using a green fluorescent protein-expressing transgenic isolate. Applied Environ. Microbiol., 69: 1695-1701.
Direct Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (Scab) in small grain cereals-A review. Plant Pathol., 44: 207-238.
CrossRefDirect Link - Ramos, A.J., D. Cuevas and V. Sanchis, 2006. Fusarium verticillioides and Fusarium graminearum infection and Fumonisin B1 and Zearalenone accumulation in resveratrol-treated corn. Food Sci. Technol. Int., 12: 353-359.
CrossRefDirect Link - Reid, L.M., R.W. Nicol, T. Ouellet, M. Savard and J.D. Miller et al., 1999. Interaction of Fusarium graminearum and F. moniliforme in maize ears: Disease progress, fungal biomass and mycotoxin accumulation. Phytopathology, 89: 1028-1037.
Direct Link - Sass, V., J. Milles, J. Kramer and A. Prange, 2007. Competitive interactions of Fusarium graminearum and Alternaria alternata in vitro in relation to deoxynivalenol and zearalenone production. Int. J. Food Agric. Environ., 5: 257-261.
Direct Link - Semagn, K., H. Skinnes, A. Bjornstad, A.G. Maroy and Y. Tarkegne, 2007. Quantitative trait loci controlling Fusarium head blight resistance and low deoxynivalenol content in hexaploid wheat population from Arina and NK93604. Crop Sci., 47: 294-303.
Direct Link - Silva, E., E.A. Mora, A. Medina, J. Vasquez, D. Valdez, D.L. Danial and J.E. Parlevliet, 2007. Fusariumear rot and how to screen for resistance in open pollinated maize in the Andean regions. Euphytica, 153: 329-337.
Direct Link - Simpson, D.R., M.A. Thomsett and P. Nicholson, 2004. Competitive interactions between Microdochium nivale var. majus, M. nivale var. nivale and Fusarium culmorum in planta and in vitro. Environ. Microbiol., 6: 79-87.
CrossRefDirect Link - Tiemann, U. and S. Dänicke, 2007. in vitro and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs. Food Additives Contaminants, 24: 306-314.
Direct Link - Usleber, E., V. Renz, E. Martlbauer and G. Terplan, 1992. Studies on the application of enzyme immunoassays for the Fusarium mycotoxins deoxynivalenol, 3-acetyldeoxynivalenol and zearalenone. Zoonoses Public Health, 39: 617-627.
CrossRefDirect Link - Phalip, V., D. Hatsch, B. Laugel and J.M. Jeltsch, 2006. An overview of fungal community diversity in diseased hop plantations. FEMS Microbiol. Ecol., 56: 321-329.
CrossRefDirect Link - Williamson, P.M., 1997. Black point of wheat: In vitro production of symptoms, enzymes involved, and association with Alternaria alternata. Aust. J. Agric. Res., 48: 13-19.
CrossRefDirect Link - Wisniewska, H. and K. Kowalczyk, 2005. Resistance of cultivars and breeding lines of spring wheat to Fusarium culmorum and powdery mildew. J. Applied Genet., 46: 35-40.
Direct Link - Yoshizawa, T., A. Yamashita and Y. Luo, 1994. Fumonisin occurrence in corn from high-and low-risk areas for human esophageal cancer in China. Applied Environ. Microbiol., 60: 1626-1629.
Direct Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link