
A. Vikram
Department of Plant Science, McGill University, Ste-Anne-De-Bellevue, Quebec, Canada
H. Hamzehzarghani
Department of Plant Science, McGill University, Ste-Anne-De-Bellevue, Quebec, Canada
A .R. Alagawadi
Department of Agricultural Microbiology, University of Agricultural Sciences, Dharwad, India
P. U. Krishnaraj
Department of Agricultural Biotechnology, University of Agricultural Sciences, Dharwad, India
B. S. Chandrashekar
Department of Agricultural Biotechnology, University of Agricultural Sciences, Dharwad, India
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 3 | Page No.: 326-333







ABSTRACT
Phosphate Solubilizing Bacteria (PSB) isolated from the crops grown in vertisols were tested for the production of Plant Growth Promoting Substances (PGPS) such as Indole Acetic Acid (IAA), Gibberellic Acid (GA) and organic acids. All the 30 isolates of PSB were able to produce both IAA and GA and the amount ranged from 1.1 to 28.0 µg/25 and from 0.6 µg to 9.8 µg/25 mL of broth, respectively. The strains PSBV-4 and PSBV-5 produced the maximum amounts of IAA and GA, respectively. The organic acids produced were gluconic acid, tartaric acid, citric acid, maleic acid, succinic acid, glyoxalic acid and a few unidentified acids. The strain PSBV-9 and the standard Pseudomonas striata produced six organic acids while strains PSBV-10, PSBV-25 and PSBV-16 produced only five organic acids. Hierarchical cluster analysis was used to separate PSBs according to IAA and GA production and the PSBs were classified into three major groups. Correspondence analysis was used to find any association between PSB isolates and organic acid production and it showed a significant association between organic acid production and PSB isolates. The PSB strains isolated here have the potential to promote plant growth, as demonstrated here based on the production of PGPS and organic acids. However, their ability to promote plant growth under greenhouse and field conditions has to be validated.
PDF Abstract XML References Citation
How to cite this article
A. Vikram, H. Hamzehzarghani, A .R. Alagawadi, P. U. Krishnaraj and B. S. Chandrashekar, 2007. Production of Plant Growth Promoting Substances by Phosphate Solubilizing Bacteria Isolated from Vertisols. Journal of Plant Sciences, 2: 326-333.
DOI: 10.3923/jps.2007.326.333
URL: https://scialert.net/abstract/?doi=jps.2007.326.333
DOI: 10.3923/jps.2007.326.333
URL: https://scialert.net/abstract/?doi=jps.2007.326.333
INTRODUCTION
Plant growth stimulation following inoculation of Phosphate Solubilizing Bacteria (PSB) to crops in soils containing low levels of phosphorus has been reported (Datta et al., 1982; Domey and Lippmann, 1989; De Freitas et al., 1997). PSB application has promoted P-uptake as well as the yields in several crops (Tomar, 1998; Khalid et al., 2004). The plant growth promoting effects of PSB is considered to be related to their ability to synthesize plant growth regulating substances (Barea et al., 1976; Sattar and Gaur, 1987). PSB is known to solubilize insoluble phosphates and make it available to plants (Rodriguez and Fraga, 1999; Vazquez et al., 2000; Kuklinsky-Sobral et al., 2004). They are also capable of producing phytohormones and growth promoting substances. The production of Indole Acetic Acid (IAA), gibberellins and cytokinins by PSB have been reported earlier by several workers (Barea et al., 1976; Sattar and Gaur, 1987; Khalid et al., 2004; Kuklinsky-Sobral et al., 2004).
The major mechanism for solubilization of insoluble inorganic phosphates by microorganisms is through production of organic acids. The production of organic acids including lactic, glycolic, citric, maleic, oxalo acetic, tartaric and succinic acids have been reported by various workers (Bajpai and Sunadara Rao, 1971; Illmer and Schinner, 1992). Thirty PSB strains isolated from the rhizosphere of crop plants grown in vertisols were found to release P from insoluble phosphates (Vikram, 2001). The objectives of this study were to test the ability of these 30 strains for production of plant growth promoting substances such as IAA, gibberellins and organic acids. A standard PSB, Pseudomonas striata was also used as a control. This study will aid in selecting effective PSB with the ability to produce growth promoting substances and organic acids which can be later tested for their growth promotional potential on crops.
MATERIALS AND METHODS
PSB Strains Used in the Study
The 30 strains of PSB used in the study were PSBV-1 to PSBV-30. Pseudomonas striata obtained from the culture bank of Department of Agricultural Microbiology, University of Agricultural Sciences, Dharwad was also used in the experiment.
Assaying for Plant Growth Promoting Substances Produced by PSB
The isolates of PSB were inoculated to Czapek’s solution (Mahadevan and Sridhar, 1984) supplemented with 0.005 M L-tryptophan. They were incubated at 37°C for seven days in dark. After incubation, the cultures were centrifuged at 6000 rpm to remove the bacterial cells and the supernatant was collected in a conical flask. The amount of Indole Acetic Acid and Gibberellic Acid (GA) were determined using the supernatant.
Estimation of IAA
Indole Acetic Acid was estimated according to the procedure of Gordon and Paleg (1957). Twenty-five milliliter of the supernatant was collected and the pH was adjusted to 2.8 using 1 N HCl in a 100 mL conical flask. Equal volume of diethyl ether was added to it and incubated in dark for 4 h. Indole Acetic Acid extraction was done at 4°C in a separating funnel using diethyl ether. The organic phase was discarded and the solvent phase was pooled and evaporated to dryness. To the dried material, 2 mL of methanol was added, pooled and the IAA present in the methanol extract was determined according to Gordon and Paleg (1957). To 0.5 mL of methanol extract, 1.5 mL of double distilled water and 4 mL of Sapler’s reagent (1 mL of 0.5 M FeCl3 in 50 mL of 35% perchloric acid) were added and incubated in dark for 1 h. The intensity of pink color developed was read at 535 nm in a spectrophotometer. From a standard curve prepared with known concentrations of IAA, the quantity of IAA in the supernatant was determined and expressed as μg/25 mL of the medium.
Estimation of GA
Amount of GA present in the supernatant of the culture was determined by the method of Paleg (1965). Twenty-five milliliter of the supernatant was taken in a test tube to which 2 mL of zinc acetate was added. After 2 min, 2 mL of potassium ferrocyanide was added and centrifuged at 1000 rpm for 15 min. To 5 mL of this supernatant, was added 5 mL of 30% HCl and incubated at 20°C for 75 min. The blank sample was treated with 5% HCl and the absorbance of the sample as well as blank was measured at 254 nm in a spectrophotometer. The amount of GA present in the extract was calculated from the standard curve and expressed as μg/25 mL of the medium. The standard curves of IAA and GA were prepared by using graded concentrations of IAA and GA3.
Detection of Organic Acids
The major mechanism for solubilization of insoluble inorganic phosphates by microorganisms is through production of organic acids. Hence, the organic acid production profile of the PSB was examined, in comparison with the standard Pseudomonas striata, by paper chromatographic method. One milliliter of 24 h old culture of each isolate was inoculated to 50 mL Pikovskaya’s broth and incubated at 28±2°C for 10 days. The broth was centrifuged at 10,000 rpm for 10 min. The supernatant so obtained was concentrated to nearly 1/10th of the original volume in a water bath maintained at 60°C. The concentrated material was then used for determination of organic acids by paper chromatography in comparison with standard organic acid (Gaur, 1990). Standards of organic acids were prepared at 20 mg mL‾1 stock. About 10 μL of standards and 15 μL of culture supernatants were spotted on Whatman No. 1 chromatographic paper and dried with a hair dryer. A descending chromatography was run using a solvent mixture of n-butanol, acetic acid and water in 12:3:5 ratio in a chromatographic chamber pre saturated with solvent for 6 h. The chromatogram was run for 16 h and air dried for 3 days. The air dried paper was sprayed with 0.04% bromocresol green. The paper was dried at room temperature. The Rf values of yellow spots of organic acids developed on a blue background were measured and compared with the Rf values of the standard organic acids for identification.
Correspondence Analysis
Correspondence analysis, a non-parametric chi square based method, was used to explore data on the frequency of occurrence of organic acids in each PSB isolate group and to find any association between PSB isolates and organic acid production. Correspondence analysis is a descriptive/graphical multivariate technique to analyze contingency table data and investigate probable similarity in categorical data and to explore the structure of categorical variables (organic acid production and PSB isolate). In correspondence analysis the categorical data are used to produce two or three dimensions as coordinates of a new reduced space in which categorical variables are projected based on chi-square distances.
Hierarchical Cluster Analysis of Organic Acids
The data on Indole Acetic Acid (IAA) and Gibberellic Acid (GA) was subjected to hierarchical cluster analysis, using cluster procedure of SAS (1999) to identify the similarity of PSB isolates and classify them into clusters with similar IAA and GA production phenotypes. Mahalanobis distance between individuals in a p-dimensional space was used to identify clusters.
The unsupervised produced clusters of PSB isolates was subjected to analysis of variance using GLM procedure of SAS and the means of clusters was compared with LSMEAN method to investigate any significant difference between clusters based on GA and IAA production.
RESULTS
Plant Growth Promoting Substances Produced by Isolates of PSB
All the 30 isolates of PSB tested here produced both IAA and GA. The amount of IAA produced by the strains ranged from 1.1 μg to 28.0 μg/25 mL broth, whereas the amount of GA produced by the strains ranged from 0.6 μg to 9.8 μg/25 mL broth. The strain PSBV-4 produced maximum amount of IAA (28 μg/25 mL broth) and the minimum amount of IAA was produced by the strains PSBV-19 and PSBV-29 (both 1.1 μg/25 mL broth). Similarly, maximum amount of GA was produced by the strain PSBV-5 (9.80 μg/25 mL broth) and the minimum amount of GA was produced by the strains PSBV-20 and PSBV-30 (both 0.60 μg/25 mL broth). The production of IAA and GA by Pseudomonas striata was 23.05 μg and 8.40 μg/25 mL broth, respectively. Among the 30 isolates of PSB, one strain recorded higher IAA than P. striata while about seven of them were able to produce higher content of GA than P. striata.
Classification of PSBs Based on the Hierarchical Cluster Analysis of the Amount of IAA and GA
The PSBs were classified into three clusters based on the hierarchical cluster analysis of the amounts of IAA and GA (Fig. 1). The three clusters were: a) Cluster 1: The PSBs namely PSBV-1, PSBV-4, PSBV-6, PSBV-10, PSBV-13, PSBV-15, PSBV-16, PSBV-17, PSBV-26 and P. striata; b) Cluster 2: PSBV-2, PSBV-5, PSBV-7, PSBV-11, PSBV-22, PSBV-24, PSBV-25 and PSBV-27; c) Cluster 3: PSBV-3, PSBV-8, PSBV-9, PSBV-12, PSBV-14, PSBV-18, PSBV-19, PSBV-20, PSBV-21, PSBV-23, PSBV-28, PSBV-29 and PSBV-30.
The strains of first cluster generated based on unsupervised method grouped tightly together and were separated distinctly from the other two groups. Isolates in the second and third clusters were significantly different from both the first cluster and each other based on IAA production (p<0.0002). According to GA production, the second and third clusters were significantly different from the first cluster however they did not differ from each other significantly (Table 1).
Organic Acids Produced by Isolates of PSB and Classification Based on Correspondence Analysis
The isolates of PSB were assayed for organic acid production profile by paper chromatography. The organic acids produced by PSB included gluconic acid, tartaric acid, citric acid, maleic acid, succinic acid, glyoxalic acid and few unidentified acids. Among the 30 strains and a standard tested, PSBV-9 and the standard Pseudomonas striata produced six organic acids namely gluconic acid, tartaric acid, citric acid, maleic acid, succinic acid and glyoxalic acid while strains PSBV-10, PSBV-25 and PSBV-16 were able to produce five organic acids. Most of the strains produced three to four organic acids. While 23 strains produced maleic acid, 20 produced gluconic acid and 19 produced citric acid and a few produced organic acids that were unidentified.
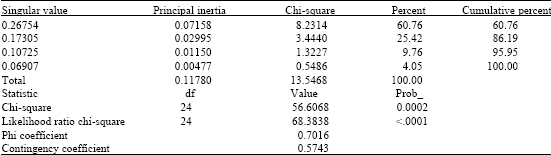
There was a significant association between organic acid production and PSB isolates (Chi square = 56.60, p = 0.0002) (Fig. 2 and Table 2). Dimension one is a vector in chi-square space that separates Xanthomonas and Bacillus from P. striata and Serratia whereas dimension two divides Pseudomonas from others.
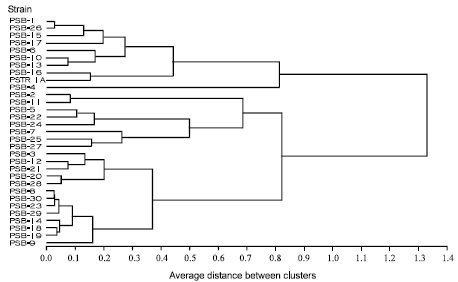 | |
| Fig. 1: | Grouping of 30 PSB and Pseudomonar striata isolates based on production of Indole Acetic Acid (IAA) and Gibberellic Acid (GA). Dendrogram was generated based on Hierarchical cluster analysis of detected amounts of IAA and GA and the scale represents the Euclidean distance of phosphate solubilizing bacterial isolates as an index of their similarity |
| Table 1: | Analysis of variance of three clusters (1, 2 and 3) of PSB isolates according to their ability to produce Indole Acetic Acid and Gibberellic Acid |
 | |
| IAA = Indole Acetic Acid and GA = Gibberellic Acid. In each row of the table the top/bottom values are t-student/P-values for rejection of null hypothesis no significant difference between the two means. Only the H0 of no significant difference between means of GA production of clusters 1 and 2 could not be rejected (labeled as ns = non-significant). G1 = PSBV-1, PSBV-4, PSBV-6, PSBV-10, PSBV-13, PSBV-15, PSBV-16, PSBV-17, PSBV-26 and P. striata; G2 = PSBV-2, PSBV-5, PSBV-7, PSBV-11, PSBV-22, PSBV-24, PSBV-25 and PSBV-27; G3 = PSBV-3, PSBV-8, PSBV-9, PSBV-12, PSBV-14, PSBV-18, PSBV-19, PSBV-20, PSBV-21, PSBV-23, PSBV-28, PSBV-29 and PSBV-30 | |
| Table 2: | Statistics for contingency table of PSB isolates by organic acids and inertia and Chi square decomposition results |
 | |
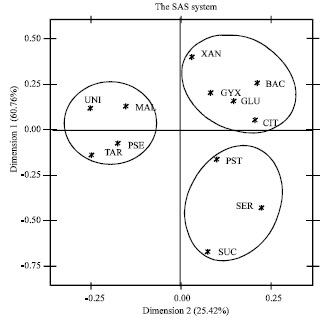 | |
| Fig. 2: | Plot of simple correspondence analysis of a 5χ7 contingency table of organic acid profile and PSB isolates from vertisol (Chi square = 56.60, p = 0.0002). Abbreviations used for bacterial isolates and organic acids are as follows: BAC = Bacillus, PSE = Pseudomonas, PST = P. striata, XAN = Xanthomonas, SER = Serratia, GLU = Gluconic acid, TAR = Tartaric acid, CIT = Citric acid, MAL = Maleic acid, SUC = Succinic acid, GYX = Glyoxalic acid, UNI = Unidentified |
Based on chi-square distance PSB isolates belonging to Pseudomonas produced tartaric acid, maleic acid and unidentified organic acids whereas PSB isolates from Xanthomonas and Bacillus were more glyoxalic acid, gluconic acid and citric acid producers. Pseudomonas striata and Serratia were closer according to Chi square scale used to calculate their distance producing more of succinic acid (Fig. 2). The distance in Fig. 2 is not simply Euclidean distance but a chi-square distance between the data points and is used to refer to the differences between the pattern of relative frequencies for the PSB isolates (rows) across the OA profiles (columns) and vice versa. The closer the data points (PSB isolates for instance) along one dimension the more similar they will be based on organic acid production and vice versa. Inertia is the weight or the amount of contribution of each data point which represents the proportion of the contribution of that point to the overall Chi-square distance. Pseudomonas and Serratia PSB isolates were separated from other PSB isolates along dimension one whereas dimension two separated maleic acid, tartaric acid and unidentified acid from other organic acids. The interpretations of the first two dimensions (with explanation of more than 85% of the inertia or Chi square) showed significant association between organic acid profile and PSB isolates.
DISCUSSION
Production of Plant Growth Promoting Substances (PGPS) like IAA, GA and cytokinins by several beneficial microorganisms including phosphate solubilizers have been quantified and related to enhancement of plant growth. In the present investigation we report the production of PGPS such as IAA, GA and organic acids by 30 isolates of PSB (Defreitas et al., 1997; Dey et al., 2004; Hameeda et al., 2006). The results on production of IAA and GA from the PSBs tested here support the observations of Barea et al. (1976) who reported the production of PGPS like IAA, GA or cytokinins or their combinations from 50 P dissolving bacteria. Reports of similar nature on the production of IAA and GA by PSB have been made by various workers (Gaur, 1990; De Freitas et al., 1997; Bastian et al., 1998; Khalid et al., 2004; Kuklinsky-Sobral et al., 2004). The amounts and range of IAA and GA produced, in general in our study, by the 30 PSBs were comparable to those reported by several others in different PSB strains (Leinhos and Vacek, 1994; Defreitas et al., 1997; Khalid et al., 2004). Production of IAA by all the PSBs tested supports the view that auxin production is common among rhizosphere bacteria (Prikryl et al., 1985; Leinhos and Vacek, 1994; Bastian et al., 1998). The large variation in the amount of IAA and GA produced by different strains has been attributed to the variability in the metabolism of the different strains of phosphate solubilizing rhizobacteria (Leinhos and Vacek, 1994). Auxins produced by the PSBs can influence several plant growth components, including root growth which in tern can improve uptake of essential nutrients thus increasing plant growth (Thimann, 1972). Thus, it is possible that the strains tested here can also provide additional growth promotional activity apart from releasing Pi into the rhizosphere. Hierarchical Cluster Analysis classified the 30 isolates of PSB and P. Striata into 3 separate groups based on their similarity of IAA and GA production which signifies that P. striata and other strains in the first group had relatively high potential of IAA and GA production as compared to the other two groups which produced the highest amounts of both IAA and GA. Although based on IAA production PSB isolates formed three distinct groups, in terms of GA production ANOVA could discriminate only two groups (groups 1 and 2 were not significantly different) (Table 1).
Phosphate solubilizing microorganisms produce a variety of organic acids from simple carbohydrates (Bajpai and Sundara Rao, 1971) by virtue of which they solubilize insoluble inorganic phosphates (Banik and Dey, 1983; Vazquez et al., 2000). Acidification of the periplasmic space by direct oxidation pathway of glucose to gluconic acid as the major cause of mineral phosphate solubilization in gram-negative bacteria has also been shown by Goldstein (1986). In the present study the 30 PSBs produced several kinds of organic acids, indicating the ability of P solubilizers to produce organic acids. Production of different organic acids like lactic, glycolic, succinic, acetic, oxalic, citric and malonic acids by PSB have been reported earlier (Venkateswarlu et al., 1984; Illmer et al., 1995) which are in line with the results of present investigation. In cluster analysis the first cluster had significantly higher GA and IAA production which could have stimulating effect on rooting and consequently P absorption as well as overall plant growth. Correspondence analysis was used to unravel any association between organic acid production and PSB isolates and it showed a strong association (Phi coefficient = 0.79, Table 2) but it could not reveal whether there was any interaction between organic acids and IAA/GA production. Further studies are necessary to illustrate any plausible interaction between production of organic acids and PGPS. Similar to our results, organic acids like gluconic acid, glycolic, oxalic, malonic and succinic acids also have been identified in phosphate solubilizers namely Bacillus firmus, Pseudomonas cepacia and Pseudomonas sp. (Banik and Dey, 1982; Illmer and Schinner, 1992, Goldstein et al., 1993) but this is the first time correspondence analysis was used to associate production of organic acids to bacterial isolates.
Although no generalization in terms of efficiency has been drawn for the number of organic acids produced by an organism, production of 2 keto-gluconic acid in combination with succinic or citric acid by an organism is shown to cause efficient solubilization of insoluble inorganic phosphates (Banik and Dey, 1983). Banik and Dey (1982) observed bacterial isolates producing succinic acid in addition to oxalic or 2-ketogluconic acid to be more efficacious and suggested that organic acids with dissimilar carbon atoms possessed better activity of phosphate solubilization. The production of IAA and GA by the strains of PSB gives an additional benefit of growth promotion in addition to its ability in P solubilization. These strains of PSB can be tested further for their effect on crop growth under greenhouse conditions as they have recorded IAA and GA contents comparable to that produced by the strain Pseudomonas striata. This will ultimately help in development of phosphate solubilizing biofertilizers which can be used in the vertisols where usually P is locked in the soil and is not easily available for plant uptake.
ACKNOWLEDGMENT
Authors are thankful to KSDA, Bangalore, India for providing required funds to undertake this project. Financial support of Shiraz University, Iran to the second author is appreciated.
REFERENCES
- Banik, S. and B.K. Dey, 1982. Available phosphate content of an alluvial soil as influenced by inoculation of some isolated phosphate-solubilizing micro-organisms. Plant Soil, 69: 353-364.
CrossRefDirect Link - Bastian, F., A. Cohen, P. Piccoli, V. Luna, R. Bottini, R. Baraldi and R. Bottini, 1998. Production of indole-3-acetic acid and gibberellins A1 and A3 by Acetobacter diazotrophicus and Herbaspirillum seropedicae in chemically-defined culture media. Plant Growth Regul., 24: 7-11.
CrossRefDirect Link - De Freitas, J.R., M.R. Banerjee and J.J. Germida, 1997. Phosphate-solubilizing rhizobacteria enhance the growth and yield but not phosphorus uptake of canola (Brassica napus L.). Biol. Fertil. Soils, 24: 358-364.
CrossRefDirect Link - Dey, R., K.K. Pal, D.M. Bhatt and S.M. Chauhan, 2004. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res., 159: 371-394.
CrossRefDirect Link - Goldstein, A.H., 1986. Bacterial solubilization of mineral phosphates: Historical perspective and future prospects. Am. J. Altern. Agric., 1: 51-57.
CrossRefDirect Link - Goldstein, A.H., R.D. Rogers and G. Mead, 1993. Mining by microbe. Biol. Technol., 11: 1250-1254.
Direct Link - Hameeda, B., G. Harini, O.P. Rupela, S.P. Wani and G. Reddy, 2006. Growth promotion of maize by phosphate-solubilizing bacteria isolated from composts and macrofauna. Microbiol. Res., 163: 234-242.
CrossRefPubMedDirect Link - Illmer, P. and F. Schinner, 1992. Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem., 24: 389-395.
CrossRefDirect Link - Khalid, A., M. Arshad and Z.A. Zahir, 2004. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol., 96: 473-480.
CrossRefDirect Link - Kuklinsky-Sobral, J., W.L. Araujo, R. Mendes, I.O. Geraldi, A.A. Pizzirani-Kleiner and J.L. Azevedo, 2004. Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ. Microbiol., 6: 1244-1251.
CrossRef - Paleg, L.G., 1965. Physiological effects of gibberellins. Ann. Rev. Plant Physiol., 16: 291-322.
CrossRefDirect Link - Rodriguez, H. and R. Fraga, 1999. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv., 17: 319-339.
CrossRefDirect Link - Sattar, M.A. and A.C. Gaur, 1987. Production of auxins and gibberellins by phosphate dissolving microorganisms. Zentral. Microbiol., 142: 393-395.
Direct Link - Tomar, R.K.S., 1998. Effect of phosphate-solubilizing bacteria and farmyard manure on the yield of blackgram (Phaseolus mungo). Indian J. Agric. Sci., 68: 81-83.
Direct Link - Vazquez, P., G. Holguin, M.E. Puente, A. Lopez-Cortes and Y. Bashan, 2000. Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol. Fert. Soils, 30: 460-468.
CrossRefDirect Link