
R. Ramakrishnan Nair
Indian Institute of Spices Research, Calicut 673012, Kerala, India
S. Dutta Gupta
1Indian Institute of Spices Research, Calicut 673012, Kerala, India
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 3 | Page No.: 283-292







ABSTRACT
A successful encapsulation protocol was developed for somatic embryos of black pepper to produce synthetic seeds. Sodium alginate concentration of 4% (w/v) was ideal at a calcium chloride concentration of 100 mM to produce capsules of perfect morphology and sufficient gel strength. Encapsulation affected the percentage of germination of somatic embryos depending upon the concentration of sodium alginate used, with a drastic reduction at 5% and above. Preliminary studies on storage indicated that the somatic embryos encapsulated in 4% sodium alginate can be successfully stored at culture room temperature on agar-gelled SH basal salt solution up to 45 days. Storage at low temperature of 4°C found to be lethal. Non-encapsulated somatic embryos were not suitable for storage. Possibility of direct sowing of synthetic seeds in the soil was also examined.
PDF Abstract XML References Citation
How to cite this article
R. Ramakrishnan Nair and S. Dutta Gupta, 2007. In vitro Plant Regeneration from Encapsulated Somatic Embryos of Black Pepper (Piper nigrum L.). Journal of Plant Sciences, 2: 283-292.
DOI: 10.3923/jps.2007.283.292
URL: https://scialert.net/abstract/?doi=jps.2007.283.292
DOI: 10.3923/jps.2007.283.292
URL: https://scialert.net/abstract/?doi=jps.2007.283.292
INTRODUCTION
High frequency plant regeneration through somatic embryogenesis is a suitable system for mass propagation of plants. The prospect of promoting synthetic seed technology in agriculturally important species utilizing somatic embryogenesis is a subject of increasing interest (Redenbaugh, 1990). Artificial or synthetic seeds, consisting of somatic embryos enclosed in a protective coating, have been proposed as a low-cost, high-volume propagation system (Redenbaugh et al., 1986). Synthetic seed technology is designed to combine the advantages of clonal propagation with those of seed propagation. By combining the benefits of a vegetative propagation system with the capability of long term storage, synthetic seeds have many diverse applications in agriculture (Gray and Purohit, 1991; Redenbaugh et al., 1991; Redenbaugh, 1993). The potential uses of artificial seeds are numerous, including storage of germplasm and delivery of elite genotypes, hand-pollinated hybrids with reduced seed fertility and genetically engineered plants with sterile or unstable genotypes. The size of the synthetic seed and the coating around the somatic embryo potentially are advantageous for storage, handling, transportation and planting (Redenbaugh et al., 1988). Synthetic seeds represent a potential delivery system for future genetically engineered cultivars that are meiotically unstable (Mariani, 1992). The potential application of synthetic seed technology has been demonstrated for several economically useful plants (Redenbaugh et al., 1986; Bapat and Rao, 1988; Mariani, 1992; Lulsdorf et al., 1993; Padmaja et al., 1995; Onay et al., 1996; Janeiro et al., 1997; Castillo et al., 1998; Ara et al., 1999; Vij et al., 2001; Choi and Jeong, 2002; Prewein and Wilhelm, 2003; Ipekci and Gozukirmizi, 2003; Jung et al., 2004). Prospects and limitations of synthetic seeds were reviewed by Ara et al. (2000) and application of artificial seeds to tropical crops has been featured by Redenbaugh (1990). According to Senaratna (1992) encapsulation of somatic embryo may control the water uptake, release of nutrients and provide mechanical protection required for field planting.
Black pepper (Piper nigrum L., Piperaceae), is one of the most important spice crops grown in India and traded throughout the world and is traditionally propagated by stem cuttings. Propagation of this crop through seeds is cumbersome, uncertain and yields only heterogeneous progenies (Ravindran et al., 2000). Black pepper seeds are recalcitrant and viability is retained only for about a week after harvest (Ravindran et al., 2000) and easily looses viability on storage, as the moisture content is lost (Chaudhury and Chandel, 1994). Thus, standardization of synthetic seed technology has potential application in this crop.
Plant regeneration through direct somatic embryogenesis from the micropylar tissues of the germinating seeds of black pepper and its scaling up through high-frequency cyclic secondary somatic embryogenesis have been described in earlier reports (Nair and Dutta Gupta, 2003, 2005, 2006). Published reports on plant regeneration from encapsulated somatic embryos of black pepper were not available. The present study describes the encapsulation of black pepper somatic embryos in sodium alginate matrix to produce synthetic seeds and successful plant regeneration. Sensitivity of encapsulated somatic embryos to different temperatures and storage media is also tested. Possibility of direct seeding of encapsulated somatic embryos in soil is also examined.
MATERIALS AND METHODS
Establishment of Embryogenic Cultures
Primary somatic embryos of black pepper cultivar Karimunda were induced on the micropylar tissues of seeds undergoing in vitro germination, on agar gelled (0.8%), full-strength, PGR-free SH (Schenk and Hildebrandt, 1972) medium containing 3.0% (W/V) sucrose (SHS30) under darkness, after 90 days of culture, following the method described by Nair and Dutta Gupta (2003 and 2005). High frequency secondary somatic embryo proliferation was achieved subsequently on PGR-free SH medium containing 1.5% sucrose (SHS15) as reported by same authors later (Nair and Dutta Gupta, 2006). Embryogenic suspension cultures were established using the proliferating secondary embryogenic clumps as inoculums and uniformity of cultures was induced by sieving through a mesh of 500 μm (Nair and Dutta Gupta, 2006). Such embryogenic suspension cultures were used as source of somatic embryos for encapsulation.
Encapsulation of Somatic Embryos and Germination
Somatic embryos at cotyledonary stage from nearly uniform suspensions, 20 days after subculture were used for encapsulation (Fig. 1A). The embryos were collected after filtering the suspension through a 500 μm nylon mesh and blot dried on sterile filter paper inside a laminar flow hood. The embryos were spread on sterile Petri dishes containing sterilized sodium alginate (Loba Chemie, India) gel of 2.0, 3.0, 4.0, 5.0 and 6.0% strength in SHS30 basal medium. These embryos were carefully picked up with sterile Pasteur pipette (4 mm inside diameter of tip), along with a drop of sodium alginate and gently dropped into a sterile solution of 100 mM calcium chloride (CaCl2 2H2O) in a 250 mL conical flask. Embryos from each sodium alginate concentration were put into different flasks labeled, respectively and kept on a gyratory shaker (80 rpm) at 25±1°C, under 36 μmol m‾2 s‾1 light, for 40 min.
After incubation, the calcium chloride solution was decanted and the beads were washed three times with sterile SH salt solution. Morphology of beads produced in each concentration of sodium alginate was recorded by photography using a Minolta SLR camera fitted with macro lens. Encapsulated somatic embryos or Synseeds were cultured in disposable Petri plates (90χ15 mm) containing a thin layer of agar gelled SHS30 basal medium, at 25±1°C, under diffused light (10 μmol m‾2 s‾1) for germination. There were 30 synseeds per petri plate. The cultures were observed at every alternate day for two weeks and the percentage of germination was recorded after two weeks.
There were three replicates per treatment and the experiment was repeated three times. Non-encapsulated cotyledonary somatic embryos formed the control. The germinated synseeds were transferred to 250 mL Erlenmeyer flasks containing 50 mL agar gelled SHS30 medium for further development of plantlets. As 4% sodium alginate concentration produced perfect beads with sufficient strength and reasonably good germination, this concentration was selected for subsequent experiments.
Storage of Encapsulated Somatic Embryos and Germination
To study the effect of the medium of storage on viability and germination of synseeds, the cotyledonary embryos were encapsulated in beads formed by a 4% solution of sodium alginate in SHS30 medium and kept on sterile petri dishes (90χ15 mm) containing different storage media such as moistened sterile filter paper, agar-gelled SH basal salt, agar-gelled SHS15 medium, sealed with parafilm and stored at 25±1°C as well as 4°C under 24 h darkness. Three sets of 30 capsules each were used in each treatment and the experiment was repeated three times. Non-encapsulated cotyledonary somatic embryos formed the control. Germination of encapsulated as well as non-encapsulated somatic embryos (control) after 15, 30 and 45 days of storage was tested on agar gelled SHS30 medium at 25±1°C, under diffused light (10 μmol m‾2 s‾1), and mean percentage of germination was recorded.
Germination of Encapsulated Somatic Embryos in Sterilized Soil
To test the germination of synseeds in soil, the freshly encapsulated somatic embryos in 4% sodium alginate following the method described previously were sown on sterilized garden soil in seed pans, moistened with double distilled water. The seed pans were covered with polythene sheets having few holes for aeration and maintained in the culture room conditions. Fifty seeds were sown in each pan and five seed pans were used as replicates. Percentage of germination was recorded after one month of sowing.
RESULTS
Encapsulation and Germination
Among the various concentrations of sodium alginate tested for encapsulating embryos, 4% produced perfect beads with sufficient strength (Fig. 1B). Capsules formed at 2% concentration were very soft and fused together in calcium chloride solution to form lumps. Sodium alginate concentration of 3% produced small oblong beads (Fig. 1C). Concentrations above 4% produced very hard capsules.
Encapsulated somatic embryos (synseeds) cultured on SHS30 basal medium started germination after one week of culture. Initially the root portion pierced though the matrix of the beads and gradually established contact with the medium (Fig. 1D). Subsequently, the shoot portion emerged. Well developed plantlets appeared after two weeks (Fig. 1E-F).
Encapsulation affected the germination percentage of somatic embryos considerably. The germination percentage of naked somatic embryos was 80.9 while that of encapsulated somatic embryos ranged from 13.3 to 73.3. The germination of synseeds depended upon the concentration of sodium alginate used. There was a decline in the percentage of germination with the increase in sodium alginate concentration. Germination of synseeds was reduced drastically above 4% concentration (Fig. 2). Germination at 4% sodium alginate concentration was 58%.
Effect of Storage Media and Temperature on Viability and Germination of Encapsulated Somatic Embryos
Of the three storage media and two storage temperatures tested for storing Encapsulated Somatic Embryos (ESE) for different durations, agar gelled SH basal salt solution at culture room temperature (25±1°C) found to be better for storage (Table 1). ESEs stored at culture room temperature (25±1°C) showed variation in germination percentage depending upon the medium of storage and duration of storage.
 | |
| Fig. 1: | Encapsulation of black pepper somatic embryos and germination of synthetic seeds. A. Cotyledonary somatic embryos used for encapsulation B. Perfect capsules produced with 4% sodium alginate C. Capsules of oblong shape resulted from 3% sodium alginate D. Initiation of germination after 1 week of culture E. Appearance of plantlets after 2 weeks F. Well developed plantlets showing vigourous growth, Bar = 2 mm in A and 10 mm in B-F |
The ESEs stored on moistened filter paper showed a germination of 36.67% after 15 days, 17.78% after 30 days and 0.0% after 45 days of storage while those stored on agar gelled SH basal salt solution showed germination percentages of 43.33, 24.44 and 12.22 after 15, 30 and 45 days of storage respectively. Germination percentages of ESEs stored in agar gelled SHS15 medium were 55.56, 53.33 and 48.89 after 15, 30 and 45 days, respectively. When SHS15 was used as storage medium, the ESEs started germination in the storage medium itself after 30 days.
| Table 1: | Effect of storage media, temperature of storage and duration of storage on germination of encapsulated somatic embryos of cv. Karimunda. Agar-gelled SH basal medium containing 3% sucrose was used as germination medium |
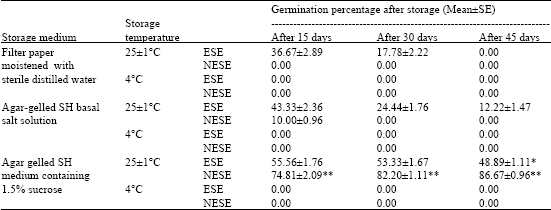 | |
| * Germination started in the storage medium itself ,** Germination in the storage medium at higher frequency | |
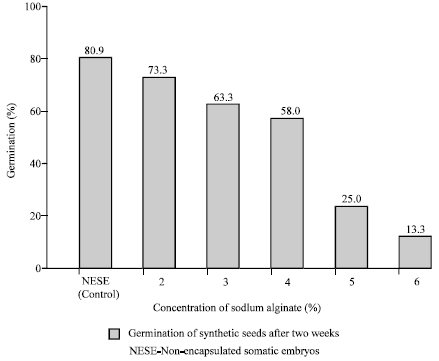 | |
| Fig. 2: | Effect of sodium alginate concentration used for encapsulation, on the germination percentage of synthetic seeds of cv. Karimunda on SH basal medium after two weeks of cultere. Non-encapsulated somatic embryos were used as control |
The ESEs stored at 4°C failed to give any germination irrespective of the storage medium or duration of storage. In most of these beads the embryos turned brown indicating lethality.
Non-encapsulated somatic embryos used as controls showed that they are not suitable for storage, irrespective of the storage media and temperatures tested in the present study. The NESEs stored on moistened filter paper at culture room temperature (25±1°C) as well as 4°C failed to show any germination after storage. Those NESEs stored on agar-gelled SH basal salt solution at culture room temperature (25±1°C) showed poor germination (10.0±0.96) after 15 days storage, but failed to germinate after 30 and 45 days of storage. On SHS15 medium used for storage at culture room temperature (25±1°C), NESEs started germinating at higher frequency even after 15 days of storage (74.81±2.09). The germination frequency increased by 30 days (82.20±1.11) and 45 days (86.67±0.96) and well developed plantlets started appearing. The NESEs stored at 4°C failed to survive and turned brown in all the storage media tested (Table 1).
Germination of Encapsulated Somatic Embryos in Sterilized Soil
Only 10% of the encapsulated somatic embryos germinated in the sterilized garden soil, after one month of sowing. The resulted plantlets were very weak with poorly developed cotyledons. They failed to grow further and gradually perished.
DISCUSSION
The principal objective of the present study was to examine the retention of viability of black pepper somatic embryos after encapsulation in the alginate matrix and survival of the encapsulated embryos on storage. Supplementary to the standardization of high frequency plant regeneration through somatic embryogenesis in black pepper (Nair and Dutta Gupta, 2003, 2005, 2006), synseeds prepared from somatic embryos provide an additional tool as an easy and novel delivery system. The major advantage of this technique is the combination of high volume production with low cost propagation (Fuji et al., 1987; Gray et al., 1995).
Following Redenbaugh et al. (1986 and 1991) and Bapat and Rao (1988) the hydrogel sodium alginate was used for encapsulation with 100 mM CaCl2 2H2O as complexing agent in the present study. Among the various concentrations tested, 4% sodium alginate found to be ideal for encapsulation of black pepper somatic embryos, to give synseeds of sufficient strength and perfect shape. Although the recommended concentration of sodium alginate is from 0.5-5% (w/v) (Redenbaugh et al., 1987b), most frequently used concentrations are at the range of 2-3%, to get the ideal results in many plant species (Jain, 1987; Rao and Singh, 1991; Mariani, 1992; Lulsdorf et al., 1993; Gill et al., 1994; Janeiro et al., 1997; Castillo et al., 1998; Ara et al., 1999; Vij et al., 2001; Malabadi and Nataraja, 2002; Choi and Jeong, 2002; Ipekci and Gozukirmizi, 2003; Lakshmi and Mythili, 2003; Jung et al., 2004; Wei-Chang and Sunitha, 2005). Higher concentrations such as 4% (Prewein and Wilhelm, 2003) and 5% (Ganapathi et al., 2001) were rarely used. Higher concentrations of sodium alginate may be required depending upon the medium in which it is dissolved and also the commercial source of the chemical. In the present study sodium alginate was dissolved in SH basal medium containing 3% sucrose (SHS30). It has been reported that sodium alginate from different sources used in same concentration may influence the germination of synseeds differently (Castillo et al., 1998; Gosh and Sen, 1994). This may be due to the difference in gel strength.
In general, alginate encapsulation affected the germination of black pepper somatic embryos compared to non encapsulated controls (Fig. 2). Reduced germination percentage of ESEs compared to NESEs was reported by earlier workers (Lulsdorf et al., 1993; Janeiro et al., 1997; Ipekci and Gozukirmici, 2003). It has been suggested that the encapsulation affect the embryo respiration (Redenbaugh, 1990) and this in turn may affect the viability and germination of somatic embryos. Conversely, Ara et al. (1999) reported that in mango ESEs given a higher percentage of germination compared to NESEs. They attributed the reasons for these results to the protection provided by the capsules and the presence of nutrients in the gel matrix. In the present context it appears that the reduced germination percentage is due to the impaired respiration and the mechanical barrier created by capsule matrix.
The percentage of germination of ESEs varied depending upon the concentration of sodium alginate used. There was a gradual decrease in germination percentage of ESEs as the concentration of sodium alginate increased and drastic reduction was observed at 5 and 6% (Fig. 2). Concentration of 4% was considered as optimum as it produced perfect beads with sufficient mechanical strength and given reasonably good germination of ESEs. Such variation in the quality of capsules and frequency of regeneration depending upon sodium alginate concentrations has already been indicated (Janeiro et al., 1997; Castillo et al., 1998; Ipekci and Gozukirmici, 2003). The increased sodium alginate concentrations may result in very hard capsules which hinder the penetration of radicle through the capsule wall to establish contact with the germination medium in most of the ESEs. Only very vigorous somatic embryos may be able to overcome such barrier, which results in low percentage of germination.
The somatic embryos encapsulated with 4% sodium alginate in SHS30 medium and stored on different storage media at different temperatures and duration showed varied responses.
Irrespective of the storage media and duration, storage at 4°C found to be lethal to the ESEs as well as NESEs (control) of black pepper. Storage of ESEs at 4°C and subsequent germination has been tested in many plant species of importance (Bapat and Rao, 1988; Datta and Potrykus, 1989; Liu et al., 1992; Lulsdorf et al., 1993; Gill et al., 1994; Onay et al., 1996; Janeiro et al., 1997; Madhav et al., 2002; Ipekci and Gozukirmizi, 2003; Winkelmann et al., 2004). In most of these cases (Bapat and Rao, 1988; Datta and Potrykus, 1989; Lulsdorf et al., 1993; Janeiro et al., 1997; Ipekci and Gozukirmizi, 2003; Winkelmann et al., 2004) storage of ESEs at 4°C reduced the germination, but complete suppression of germination was not observed. Germination percentage of NESEs stored at 4°C has been reported to be lower than ESEs in certain cases (Janeiro et al., 1997; Ipekci and Gozukirmizi, 2003) and rarely NESEs failed to survive the storage (Datta and Potrykus, 1989). Non germination of ESEs and NESEs of black pepper after storage at 4°C from 15-45 days indicates the high sensitivity of black pepper somatic embryos to low temperature which can not be overcome by encapsulation alone. Special treatments have to be standardized for solving this problem.
Germination percentage of the ESEs stored at culture room temperature of 25±1°C was influenced by the storage medium and duration of storage. In all the storage media tested, germination percentage of ESEs after storage at 25±1°C was lower (Table 1) than the freshly encapsulated somatic embryos in 4% alginate (Fig. 2) and declined depending upon the duration of storage. Reduced germination percentage of ESEs stored at culture room temperature for different time durations has been recorded by Janeiro et al. (1997) in Camellia japonica and Malabadi and Nataraja (2002) in Clitoria ternatea. Similar observations were also made by Vij et al. (2001) in encapsulated PLBs of Dendrobium densiflorum. These authors preferred 4°C as a better storage temperature than culture room temperature. But, in all these cases ESEs were stored without any storage media. According to Redenbaugh (1990), the hydrated capsules are more difficult to store because of the requirement of embryo respiration. The ideal storage medium for storing ESEs upto 45 days found to be agar-gelled SH basal salt solution (Table 1). This may be due the fact that the essential nutrients required except the sucrose is available from the storage medium once the nutrients in the gel matrix is depleted and absence of sucrose in the medium prevents the germination of ESEs during storage. In agar gelled SH medium supplemented with 1.5% sucrose (SHS15), germination frequency was higher after storage compared to agar-gelled SH basal salt solution, but ESEs started germination in the storage medium itself after 15 days of storage which impairs the purpose of storage. In filter paper moistened with sterile distilled water ESEs were viable only upto 30 days and frequency of germination was also very poor. All these observations indicate the role of nutrients on post storage germination of ESEs and essentiality of optimizing a storage medium for the somatic embryos encapsulated in sodium alginate. Studies on the effect of storage media on viability and storage potential were lacking in literature. Non-encapsulated somatic embryos stored as control failed to survive on moistened filter paper, but survived upto 15 d on agar gelled SH basal salt and a poor post-storage germination percentage was observed. This may be due to the total absence of nutrients in former and absence of energy source (sucrose) in the latter. Poor survival of NESEs on storage has been indicated in earlier reports (Janeiro et al., 1997; Malabadi and Nataraja, 2002). Malabadi and Natraja (2002) observed a very low percentage (2%) of germination of NESEs stored at 20°C and complete failure of germination in NESEs stored at 27°C. In agar gelled SHS15 medium the NESEs started germination at high frequency in the storage medium itself after 15 days. Germination of cotyledonary somatic embryos of black pepper in SHS15 medium at high frequency under normal culture conditions has already been reported (Nair and Dutta Gupta, 2003, 2006).
The results on direct sowing of ESEs of black pepper indicate that the technique has to be further perfected. Only 10% of ESEs sown on sterilized garden soil germinated and the resulted weak plantlets failed to grow further. This may be due to the reason that the nutrients available in the gel matrix are insufficient for the full development of plantlets. It is also possible that the ESEs are undergoing desiccation in the soil, which can not be tolerated by highly recalcitrant somatic embryos of black pepper. Further research on this aspect should improve the nutrient status of gel matrix and standardize an external coating of ESEs to avoid desiccation. The germination of synthetic seeds was tested in the soil by a few (Bapat and Rao, 1988; Patnaik et al., 1996). Bapat and Rao (1988) tested synthetic seeds of sandal wood in soil, but failed to get any germination. Artificial seeds of Cymbopogon martini planted in sand, sand + soil or sand + soil +compost have given germination percentages of 11.7-21.7% (Patnaik et al., 1996). Thus, the present result is encouraging, as a preliminary attempt.
In conclusion, the present study successfully describes the encapsulation of somatic embryos of a cultivar of black pepper and germination. Possibility of storage of ESEs at different media, temperature and time duration is examined and a preliminary attempt is made to germinate the ESEs on sterilized garden soil. However, further research is required to perfect these techniques for their effective application in germplasm conservation and commercial micropropagation
REFERENCES
- Ara, H., U. Jaiswal and V.S. Jaiswal, 1999. Germination and plantlet regeneration from encapsulated somatic embryos of mango (Mangifera indica L.). Plant Cell Rep., 19: 166-170.
CrossRef - Ara, H., U. Jaiswal and V.S. Jaiswal, 2000. Synthetic seed: Prospects and limitations. Curr. Sci., 78: 1438-1444.
Direct Link - Bapat, V.A. and P.S. Rao, 1987. Sandalwood plantlets from synthetic seeds. Plant Cell Rep., 7: 434-436.
CrossRef - Choi, Y.E. and J.H. Jeong, 2002. Dormancy induction of somatic embryos of Siberian ginseng by high sucrose concentrations enhances the conservation of hydrated artificial seeds and dehydration resistance. Plant Cell Rep., 20: 1112-1116.
CrossRef - Datta, S.K. and I. Potrykus, 1989. Artificial seeds in barley: Encapsulation of microspore derived embryos. Theor. Applied Genet., 77: 820-824.
CrossRef - Fuji, J., D. Slade, K. Redenbaugh and K. Walker, 1987. Artificial seeds for plant propagation. Trends Biotech., 5: 335-339.
CrossRef - Ganapathi, T.R., L. Srinivas, P. Suprasama and V.A. Bapat, 2001. Regeneration of plants from alginate-encapsulated somatic embryos of banana cv. Rasthali (Mussa sp. AAB Group). In vtro Cell. Dev. Biol. Plant, 37: 178-181.
Direct Link - Ghosh, B. and S. Sen, 1994. Plant regeneration from alginate encapsulated somatic embryos of Asparagus cooperi baker. Plant Cell Rep., 13: 381-385.
CrossRefDirect Link - Ipekci, Z. and N. Gozukimizi, 2003. Direct somatic embryogenesis and synthetic seed production from Paulownia elongate. Plant Cell Rep., 22: 16-24.
CrossRefDirect Link - Janeiro, L.V., A. Ballester and A.M. Vieitez, 1997. In vitro response of encapsulated somatic embryos of camellia. Plant Cell Tiss. Org. Cult., 51: 119-125.
CrossRef - Jung, S.J., E.S. Yoon, J.H. Jeong and Y.E. Choi, 2004. Enhanced post-germinative growth of encapsulated somatic embryos of Siberian ginseng by carbohydrate addition to the encapsulation matrix. Plant Cell Rep., 23: 365-370.
Direct Link - Nair, R.R. and S.D. Gupta, 2003. Somatic embryogenesis and plant regeneration in black pepper (Piper nigrum L.): I. Direct somatic embryogenesis from tissues of germinating seeds and ontogeny of somatic embryos. J. Hortic. Sci. Biotechnol., 78: 416-421.
CrossRefDirect Link - Nair, R.R. and S.D. Gupta, 2005. Effect of explants and genotypes on primary somatic embryogenesis in black pepper (Piper nigrum L.). Cytologia, 70: 192-202.
Direct Link - Castillo, B., M.A.L. Smith and U.L. Yadava, 1998. Plant regeneration from encapsulated somatic embryos of Carica papaya L. Plant Cell Rep., 17: 172-176.
CrossRefDirect Link