
A. Nagarajan
Department of Biotechnology, Laboratory of Applied and Industrial Microbiology, Bhupat and Jyoti Mehta School of Biosciences, Indian Institute of Technology Madras, Chennai, 600036, India
N. Thirunavukkarasu
Department of Botany, Ramakrishna Mission Vivekananda College, Chennai, 600004, India
T.S. Suryanarayanan
Vivekanandha Institute of Tropical Mycology, Ramakrishna Mission Vidyapith, Chennai, 600004, India
Sathyanarayana N. Gummadi
Department of Biotechnology, Laboratory of Applied and Industrial Microbiology, Bhupat and Jyoti Mehta School of Biosciences, Indian Institute of Technology Madras, Chennai, 600036, India
Research Journal of Microbiology
Year: 2014 | Volume: 9 | Issue: 4 | Page No.: 163-176







ABSTRACT
L-asparaginase (E.C.3.5.1.1) has been commonly used for the treatment of acute lymphoblastic leukemia in adults and children. It is also used in food industry to reduce acrylamide formation during the preparation of fried food items containing starch at high temperatures. Several microorganisms from the diverse group of bacteria and yeast were reported to be used for L-asparaginase production however, many of the strains also coproduce L-glutaminase which is highly undesirable as it results in cellular stress and neurotoxicity. Thus identification of new sources for the production of glutaminase free L-asparaginase needs to be explored. In this study, we screened endophytic fungi isolated from trees of moist deciduous and semi evergreen forests of the Western Ghats and plants growing in Rono Hills, Arunachal Pradesh, India for the production of glutaminase free L-asparaginase. Using a simple agar plate assay, we found that 33 strains were positive for the L-asparaginase activity among which 19 strains showed glutaminase free L-asparaginase activity. Our results show that: Alternaria sp. endophytic in the leaf of Withania somnifera and growing in the moist deciduous forest of the Western Ghats showed maximum enzyme activity. Optimization of process parameters reveal that maximum L-asparaginase production was observed at 96 h of fermentation and high concentration of glucose in the medium as the carbon source inhibited enzyme production in Alternaria sp. This is the first report on production of glutaminase free L-asparaginase by fungal endophyte Alternaria sp.
PDF Abstract XML References Citation
Received: January 28, 2014;
Accepted: April 06, 2014;
Published: June 02, 2014
How to cite this article
A. Nagarajan, N. Thirunavukkarasu, T.S. Suryanarayanan and Sathyanarayana N. Gummadi, 2014. Screening and Isolation of Novel Glutaminase Free L-asparaginase from Fungal
Endophytes. Research Journal of Microbiology, 9: 163-176.
URL: https://scialert.net/abstract/?doi=jm.2014.163.176
URL: https://scialert.net/abstract/?doi=jm.2014.163.176
INTRODUCTION
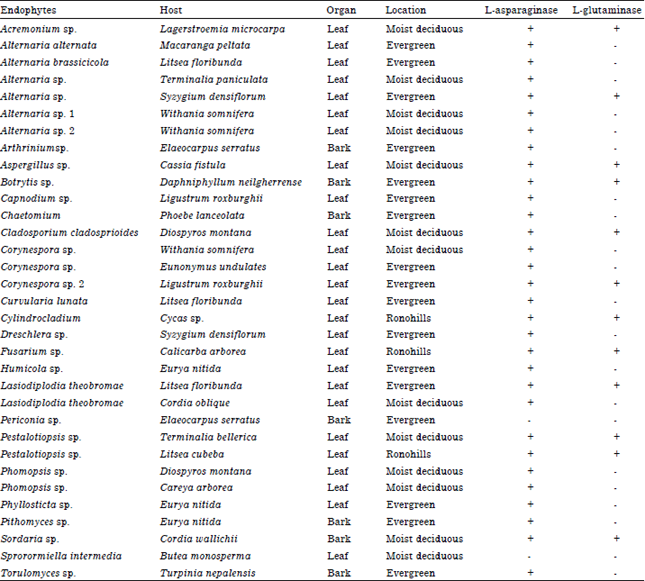
L-asparaginase (E.C.3.5.1.1) is an enzyme which is present in diverse types of microorganisms, plants and animals (Mohan Kumar et al., 2013). L-asparaginase has been commercially used to prevent acrylamide formation in fried food products (Pedreschi et al., 2008; Mohan Kumar et al., 2013). L-asparaginase from bacterial sources has been reported to be used especially as a therapeutic agent in the treatment of acute lymphoblastic leukemia in children (Muller and Boos, 1998; Pieters et al., 2011). This enzyme depletes malignant cells by preventing the formation of essential growth factors for tumor development. L-asparaginase production by various microorganisms including, Erwinia carotovora (Maladkar et al., 1993), Escherichia coli (Cedar and Schwartz, 1968; Wei and Liu, 1998), Aspergillus sp. (Sarquis et al., 2004) and also from marine derived fungal endophytes such as Fusarium sp., Phomopsis sp., Trichoderma sp. and Sargasam wightii (Thirunavukkarasu et al., 2011) has been studied extensively. A recent report revealed that clinical E. coli L-asparaginase is an important drug in treatment of patients with Acute Lymphoblastic Leukemia (ALL) (Rytting, 2012). Though it is a strong candidate for treatment of ALL, several secondary complications hinder the use of this enzyme in tumor treatment including side effects such as breathing problems, neural disorders, pancreatitis and also affects reproductive fertility (Duval et al., 2002). Pyrococcus furiosus and its mutants MTCC 5580-5582 produces L-asparaginase which was found to be highly stable (Kundu et al., 2013). Aspergillus tamari and Aspergillus terreus have been used for production of L-asparaginase but the yields were very low due to the presence of glutaminase and urease (Sarquis et al., 2004). Aspergillus niger and Aspergillus oryzae have been preferred because of their high yields (Laan et al., 2008; Eisele et al., 2011). L-asparaginase from Bacillus licheniformis (RAM 8) with low glutaminase levels was optimized for the removal of acrylamide that is formed in baked and fried food (Mahajan et al., 2012). Although the production of L-asparaginase has been reported from various organisms, the major constraint in commercialization is L-glutaminase coproduction with L-asparaginase wich leads to toxicity. Very few reports are available on glutaminase free L-asparaginase produced by microorganisms. An intercellular glutaminase free L-asparaginase from few microorganisms such as Pectobacterium carotovorurm MTCC 1428 (Kumar et al., 2011), Vibrio succinogenes (Distasio et al., 1982), Pseudomonous stutzeri (Manna et al., 1995), Pyrococcus furiosus and its mutants MTCC 5580-5582 (Kundu et al., 2013) has been reported. Endophytic fungi reside inside living tissues of all groups of plants without causing any disease in them. These fungi are known to produce novel bioactive compounds (Gunatilaka, 2006; Weber, 2009; Suryanarayanan et al., 2009; Debbab et al., 2011). These fungi have also been studied for their use in biological control of plant diseases (Vega, 2008; Rocha et al., 2011). Our recent studies and those by others indicate that endophytic fungi are also a source of novel industrial enzymes (Nagaraju et al., 2009; Rajulu et al., 2011; Suryanarayanan et al., 2012; Robl et al., 2013). In an earlier study, it was reported that many of the endophytic fungi associated with marine algae produce L-asparaginase (Thirunavukkarasu et al., 2011). In this study, we screened endophytic fungi isolated from trees of moist deciduous and semi evergreen forests of the Western Ghats and plants growing in Rono Hills, Arunachal Pradesh, India for the production of glutaminase free L-asparaginase.
MATERIALS AND METHODS
L-asparagine and L-glutamine was procured from Sigma-Aldrich. All chemicals used in the production medium and protein analysis were of analytical grade and of the highest purity from SRL, Hi-Media and Merck, India.
Isolation of endophytes: From each plant species, mature healthy leaves or outer bark were collected, washed thoroughly in running tap water and cut into segments (0.5 cm2) and surface sterilized by serial washing in 70% ethanol and sodium hypochlorite (Suryanarayanan et al., 1998). They were then plated on antibiotic-amended Potato Dextrose Agar (PDA) medium in petri dishes, sealed using ParafilmTM and incubated in a light chamber with 12 h light: 12 h dark cycle for 28 days at 26±1°C (Bills and Polishook, 1992). The tissue segments were observed periodically and the endophytic fungi growing out of them were cultured in PDA slants and were identified using standard taxonomic keys (Ellis, 1971, 1976; Subramanian, 1971; Barnett and Hunter, 1972; Von Arx, 1974; Sutton, 1980; Onions et al., 1981; Ellis and Ellis, 1987).
Plate assay for L-asparaginase: A modified Czapek Dox (CD) medium (glucose 2 g L-1, L-asparagine 10 g L-1 or L-glutamine 10 g L-1, KH2PO4 1.52 g L-1, KCl 0.52 g L-1, MgSO4.7H2O 0.52 g L-1, FeSO4.7H2O 0.01 g L-1 and agar 20 g L-1) was used for plate assay (Thirunavukkarasu et al., 2011). A 2.5% stock solution of phenol red was prepared in ethanol (pH 6.2) and 3 mL of this was added to 1000 mL of Czapek Dox medium. A mycelial plug (5 mm diameter) cut from the growing margin of the colony of an endophyte or phellophyte was placed in a petri dish containing 20 mL of this medium. After 72 h of incubation at 26±1°C, the appearance of a pink zone around the fungal colony in an otherwise yellow medium indicated L-asparaginase activity (Gulati et al., 1997). In order to study the effect of glucose on growth and enzyme production, fungal endophytes were inoculated on modified CD media with and without 2% (w/v) glucose. Fungal growth and the enzyme zone were measured as described earlier.
Shake flask experiments for enzyme production: Fungal endophyte strains producing glutaminase free L-asparaginase were maintained and sub-cultured every 7 days in PDA medium and incubated at room temperature. The production of L-asparaginase was optimized in modified CD medium (Kumar et al., 2010). Modified CD medium contained glucose 2.0 g L-1, L-asparagine 10.0 g L-1or L-glutamine 10.0 g L-1, KH2PO4 1.52 g L-1, KCl 0.52 g L-1, MgSO4.7H2O 0.52 g L-1, (NH4)2SO4 5.0 g L-1, FeSO4.7H2O 0.01 g L-1 and the initial pH was set to 6.2 (Thirunavukkarasu et al., 2011; Kumar et al., 2011). Seven days old fungal endophyte mycelium plug was cut (~5 mm diameter) using cork borer and inoculated in 50 mL modified CD medium in a 250 mL Erlenmeyer flask. The culture flask was incubated at 30°C and 180 rpm in an orbital shaker for 120 h. Samples were withdrawn at regular intervals of 24 h and used for the measurement of L-asparaginase production. In order to study the effect of carbon and nitrogen source, varying concentrations of glucose (1.0, 2.0 and 5.0 g L-1) and L-asaparagine (5.0, 10.0 and 20.0 g L-1) was incorporated in CD medium and growth was carried out for five days. The samples were collected for every 24 h and assayed as mentioned above.
Enzyme assays: The culture filtrates were centrifuged at 10,000xg for 10 min at 4°C and their protein contents were determined by Lowry method (Lowry et al., 1951). The enzyme assay reaction mixture contained 0.5 mL of 0.1 M potassium phosphate buffer pH 8.0, 0.1 mL of 40 mM L-asparagine and 100 μg of extracellular protein (culture filtrate of each endophyte) and final volume was made upto 2.0 mL with distilled water. Reaction mixture was incubated at 37°C in stirred water bath for 30 min and enzymatic reaction was stopped by adding 0.5 mL of 1.5 M Trichloro Acetic Acid (TCA). The reaction mixture was centrifuged at 10,000xg for 5 min at 4°C to remove precipitates. The ammonia released was determined using colorimetric method by adding 200 μL Nessler’s reagent into a tube containing 100 μL supernatant and 1700 μL distilled water. This mixture was vortexed and incubated at room temperature for 20 min and absorbance was measured at 436 nm (Kumar et al., 2010). Ammonium sulphate was used to plot the standard curve to determine ammonia released in the reaction mixture (Mahajan et al., 2012). One unit (IU) of L-asparaginase activity is determined as the amount of enzyme needed to liberate 1 μmol of ammonia per min at 37°C when L-asparagine is used as substrate (Imada et al., 1973; Kumar et al., 2010). Glutaminase activity was measured by direct nesslerization as described by Mashburn and Wriston (1964), using L-glutamine as the substrate. One Unit (IU) of L-glutaminase activity is calculated as the amount of enzyme needed to liberate 1 μmol of ammonia in presence of L-glutamine per min at 37°C. Specific activity of protein is defined as enzyme acitivity per milligram of protein used in the assay.
Statistical analysis: All the experiments are performed at least four times and one way analysis of variance (ANOVA) was performed in Minitab 16. A p<0.05 was considered significant.
RESULTS AND DISCUSSION
Screening of fungal endophytes for glutaminase free L-asparaginase: Thirty three different fungal endophyte strains were collected from plant hosts in Western Ghats and Rono Hills in India. The samples were critically scrutinized for the production of L-asparaginase by plate assay method. The plate assay method involves the use of Czapeck Dox medium with L-asparagine as a nitrogen source and phenol red as a pH indicator to monitor the changes in pH. If the organism produces L-asparaginase, it could grow on a normal plate containing L-asparagine and degrades it into aspartic acid and ammonia. Release of ammonia in the medium changes the pH towards alkaline which is indicated by a visible change in medium colour to pink. This pink zone was initially used to screen the fungal endophytes (Gulati et al., 1997; Thirunavukkarasu et al., 2011). For example C. lunata showed pink zone only when grown in plate containing L-asparaginase and not in plate containing L-glutamine (Fig. 1a), suggesting that the strain produces glutaminase free L-asparaginase. While another strain, L. theobromae showed pink zone when grown both on L-asparagine as well as in L-glutamine plate implying that this strain produces both the enzymes (Fig. 1b).
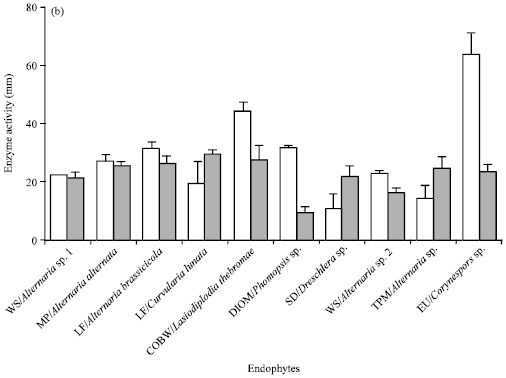
In order to screen the endophytes that produces glutaminase free L-asparaginase, all 33 fungal endophytes were allowed to grow on the modified CD medium containing L-glutamine as a nitrogen source and the colour change was monitored. Endophytes that produced pink colour zone on L-glutamine medium containing plates (Fig. 1b) were eliminated. Of the fungi studied, only a species of Periconia and S. intermedia lacked asparaginase activity (Table 1). Twelve fungi, viz., Acremonium sp., Alternaria sp. 2, Aspergillus sp., Botrytis sp., C. cladosporioides, Corynespora sp., Cylindrocladium sp., Fusarium sp., L. theobromae, Pestalotiopsis sp. 1 and 2 and Sordaria sp., were positive for both the enzymes (Table 1). Some strains producing only glutaminase free L-asparaginase (Fig. 1a) were selected and plate assay was performed. Nineteen fungal endophytes were shown to be producing glutaminase free L-asparaginase which were further examined for the highest activity of L-asparaginase production by measuring the colony diameter (mm) and diameter of the pink colour zone (mm) which indicates the enzyme production quantitatively (Fig. 2).
Effect of glucose on colony growth and enzyme activity by plate assay: Previous reports showed that concomitant addition of L-asparagine and glucose inhibits growth and activity (Geckil et al., 2006). Few reports showed that presence of glucose in media marginally favours activity and is a good choice compared to other substrates because of its low cost, making it an economically viable process (Kumar et al., 2010). The existing reports are contradictory regarding the effect of glucose and nitrogen. Hence, to check the effect of glucose on growth and L-asparaginase production, plate assay was performed in the presence and absence of glucose.
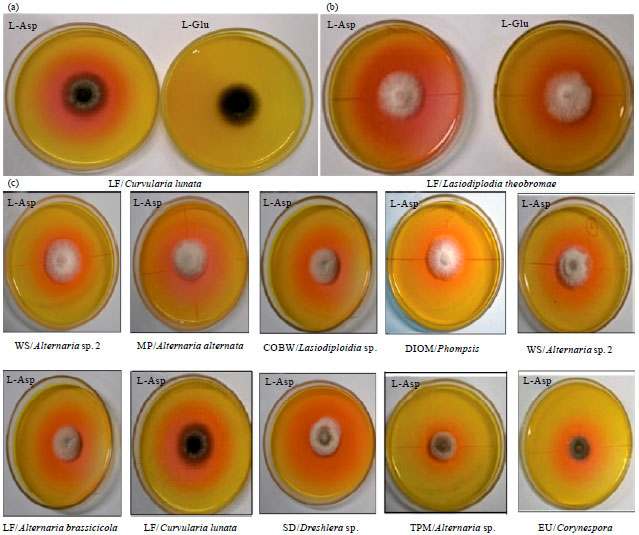 | |
| Fig. 1(a-c): | Plate assay for screening fungal endophytes producing L-asparaginase, (a) Curvularia lunata grown on plates containing L-asparagine and L-glutamine showing pink zone only when L-asparagine was used as the substrate, (b) Lasiodiplodia theobromae showing pink zone on both plates containing L-asparagine and L-glutamine as substrate and (c) Glutaminase free L-asparaginase showing pink colour zones around the fungal endophyte colony |
Among the 19 fungal endophytes screened and isolated that exhibited glutaminase free L-asparaginase activity, 10 strains which showed relatively higher enzyme activity zone were taken and grown in the presence and absence of media containing 2 g L-1 glucose. Results showed that the presence of glucose inhibited colony growth and enzyme activity in most of the endophytes (Fig. 3a and b). Curvularia lunata, Phomopsis sp. and Dreschlera sp. showed an increase in both colony growth and enzyme activity in the presence of glucose. Presence of glucose for Corynespora sp. did not have much effect on the colony growth but the enzyme activity zone was drastically reduced (Fig. 3b) (Narayana et al., 2008). As a widely used carbon source, effect of glucose on L- asparaginase activity has been well studied over the years.
| Table 1: | Isolated novel fungal endophytes screened for glutaminase free L-asparaginase production (plate assay) |
 | |
Glucose is reported to induce the production of L-asparaginase in several microbes including certain strains of Serratia marcescens (Sukumaran et al., 1979) and Bacillus circulans (Hymavathi et al., 2010). Glucose acts as a repressor and inhibits L-asparaginase production in a few organisms such as Streptomyces albidoflavus (Narayana et al., 2008) and Serratia marcescens (Heinemann and Howard, 1969). It has been reported that when 0.4% of glucose was used, L-asparaginase production was not inhibited but by further increase in the glucose concentration upto 1%, pH of the medium changed from alkaline (pH 7.5-8.0) to acidic (pH 5.2-6.9) and L-asparaginase production was inhibited (Heinemann and Howard, 1969).
L-asparaginase production by shake flask: L-asparaginase producing microorganisms either produce this enzyme constitutively or upon induction. The physico-chemical conditions for L-asparaginase production differ among various microorganisms (Savitri and Azmi, 2003). In this study, endophytes produced L-asparaginase when grown only in the presence of L-asparagine. L-asparaginase produced was determined by colorimetric estimation of released ammonia at 436 nm. Ten fungal endophytes, which exhibited highest activity in plate assay were taken for production studies in shake flask. Among the 10 fungal endophytes, WS/Alternaria sp. 2 exhibited the highest activity of 1.17±0.04 U mg-1 at 96 h, followed by TPM/Alternaria sp. 0.5±0.06 U mg-1 (Fig. 4).
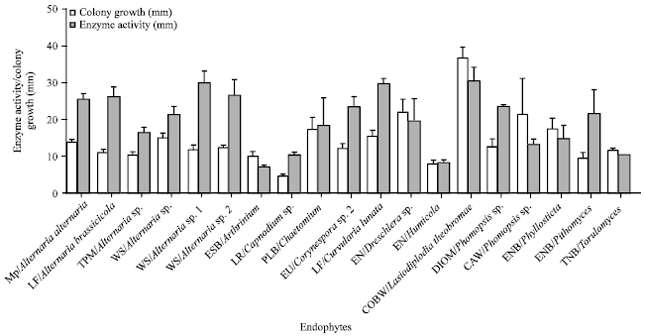 | |
| Fig. 2: | Colony growth and enzyme activity for fungal endophytes. Plate assay method was used to calculate the colony growth and enzyme activity by measuring the colony zone diameter (mm) and enzyme activity zone diameter (mm) in the plates. Experiment was repeated thrice and the error bars represent standard error of mean |
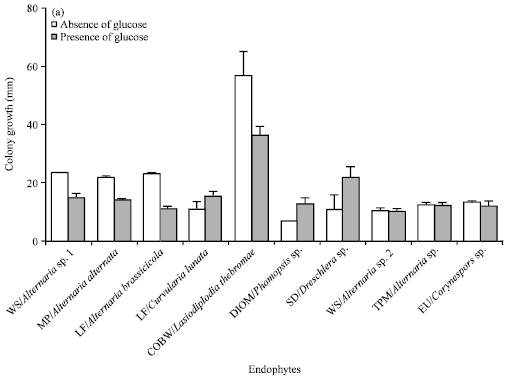  | |
| Fig. 3(a-b): | Effect of glucose on (a) Colony growth and (b) Enzyme activity by plate assay in L-asparaginase producing fungal endophytes. There is a significant decline in L-asparaginase activity of DIOM/Phomopsis sp., WS/Alternaria sp., 2 and EU/Corynespora sp. in the presence of glucose (p<0.05, by ANOVA). In WS/Alternaria sp. 1, Alternaria alternata and WS/Alternaria brassicicola, the mean colony growth decreases significantly (p<0.05, by ANOVA) in the presence of glucose. Experiment was repeated thrice and the error bars represent standard error of mean |
Though other class of microorganisms has been reported to produce L-asparaginase activity with higher and comparable values, this is the first report to show glutaminase free L-asparaginase activity by fungal endophytes.
Effect of carbon source on L-asparaginase production: Glucose is commonly used as the primary carbon source for most of the microorganisms producing primary and secondary metabolites. Since we have already confirmed that glucose play a key role in enzyme activity by plate assay method, it is essential to optimize the glucose concentration for maximizing the enzyme production in shake flasks. WS/Alternaria sp. 2, was grown in CD media containing 1, 2 and 5 g L-1 of glucose, respectively and samples were collected for every 24 till 120 h (Fig. 5a). Our results showed that culture grown in 1 g L-1 of glucose exhibited a maximum activity of 2.1 U mg-1 at 96h, whereas culture grown in 2 and 5 g L-1 for 96 h showed 0.34 and 0.31 U mg-1, respectively (Fig. 5b). Thus it is confirmed that 1 g L-1 was optimal for L-asparaginase production in WS/Alternaria sp. 2. The enzyme production beyond 96 h decreases drastically which could be due to several reasons.
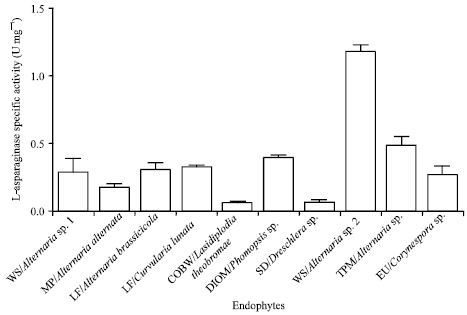 | |
| Fig. 4: | Specific activity of glutaminase free L-asparaginase in shake flask experiments. WS/Alternaria sp. 2 shows maximum specific activity. Experiment was repeated thrice and the error bars represent standard error of mean |
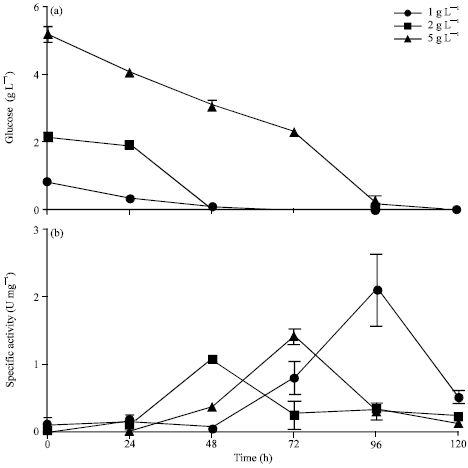 | |
| Fig. 5(a-b): | Effect of glucose on specific activity of glutaminase free L-asparaginase produced in shake flask method, (a) WS/Alternaria sp. was inoculated in the modified CD medium containing various concentration of glucose and (b) Specific activity at varying glucose concentrations |
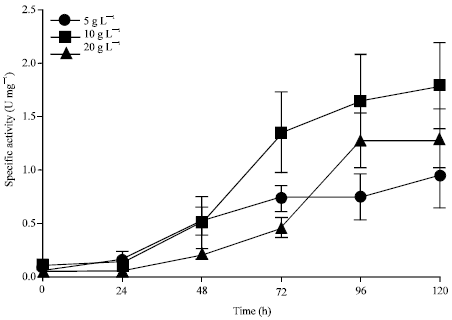 | |
| Fig. 6: | Effect of L-asparagine on specific activity of glutaminase free L- aspariginase in shake flask at varying concentrations of L-asparagine 5, 10 and 20 g L-1. Experiment was repeated thrice and the error bars represent standard error of mean |
It was observed that the production of the L-asparaginase was significantly reduced in the presence of glucose at higher concentrations, where it act as repressor for L-asparaginase in Enterobacter aerogenes (Mukherjee et al., 2000) and similar trend was observed in Fusarium sp. (Thirunavukkarasu et al., 2011), Serrtia marcescenss (Heinemann et al., 1970) and Erwinia aroideae (Liu and Zajic, 1972).
Effect of nitrogen concentration on L-asparaginase production: To study the effect of nitrogen source concentration on L-asparaginase production by WS/Alternaria sp. 2, different concentrations of L-asparagine (5, 10 and 20 g L-1) was supplemented in the media. Culture grown in 10 g L-1 of L-asparagine showed maximum activity of 1.8 U mg-1 at 120 h, followed by 1.3 and 0.9 U mg-1 for cultures grown on 5 and 20 g L-1, respectively (Fig. 6). The time dependent increase in the enzyme activity shows that L-asparaginase expression increases in the presence of L-asparagine, suggesting that it is nitrogen regulated and inducible as observed in other microbes (Sarquis et al., 2004). Effect of varying concentration (0.5-2.0%) of L-asparagine as a sole nitrogen source on L-asparaginase production was studied in Streptomyces ABR2 and the optimum concentration was determined to be 1.0% (Mostafa and Salama, 1979; Sudhir et al., 2012). Another report showed that 1% (w/v) of L-asparagine in the media exhibited maximum production of L-asparaginase by Aspergillus terreus MTCC 1782. The activity of L-asparaginase decreases with increase in concentration of L-asparagine above 1% which might be due to substrate inhibition (Shaffer et al., 1988; Baskar and Renganathan, 2011).
CONCLUSION
In this study, several endophytes have been identified that are capable of producing glutaminase free L-asparaginase. Among all the screened endophytes, WS/Alternaria sp. 2, produced maximum glutaminase free L-asparaginase. The optimal concentrations of glucose and L-asparagine were found to be 10 and 1 g L-1, respectively. Under these conditions, the strain produced a maximum specific activity of 1.65 U mg-1. This is the first report on the production of glutaminase free L-asparaginase by endophytic fungi Alternaria sp. Although, preliminary, the study gains importance as it identifies a novel eukaryotic (and hence possibly more human-compatible than a bacterial source of the enzyme) source of such a desirable enzyme for therapeutic uses. Further media components screening and optimization in shake flasks and bioreactors are needed to enhance the productivity.
REFERENCES
- Baskar, G. and S. Renganathan, 2011. Optimization of media components and operating conditions for exogenous production of fungal L-asparaginase. Chiang Mai J. Sci., 38: 270-279.
Direct Link - Bills, G.F. and J.D. Polishook, 1992. Recovery of endophytic fungi from Chamaecyparis thyoides. Sydowia, 44: 1-12.
Direct Link - Cedar, H. and J.H. Schwartz, 1968. Production of L-asparaginase II by Escherichia coli. J. Bacteriol., 96: 2043-2048.
PubMed - Debbab, A., A.H. Aly and P. Proksch, 2011. Bioactive secondary metabolites from endophytes and associated marine derived fungi. Fungal Diversity, 49: 1-12.
CrossRef - Robl, D., P. Da Delabona, C.M. Mergel, J.D. Rojas and P. dos Costa et al., 2013. The capability of endophytic fungi for production of hemicellulases and related enzymes. BMC Biotechnol., Vol. 13.
CrossRef - Duval, M., S. Suciu, A. Ferster, X. Rialland and B. Nelken et al., 2002. Comparison of Escherichia coli-asparaginase with Erwinia-asparaginase in the treatment of childhood lymphoid malignancies: Results of a randomized European organisation for research and treatment of cancer-children's leukemia group phase 3 trial. Blood, 99: 2734-2739.
CrossRef - Eisele, N., D. Linke, K. Bitzer, S. Na'amnieh, M. Nimtz and R.G. Berger, 2011. The first characterized asparaginase from a basidiomycete, Flammulina velutipes. Bioresour. Technol., 102: 3316-3321.
CrossRef - Geckil, H., S. Gencer, B. Ates, U. Ozer, M. Uckun and I. Yilmaz, 2006. Effect of Vitreoscilla hemoglobin on production of a chemotherapeutic enzyme, L‐asparaginase, by Pseudomonas aeruginosa. Biotechnol. J., 1: 203-208.
CrossRef - Rajulu, M.B.G., N. Thirunavukkarasu, T.S. Suryanarayanan, J.P. Ravishankar, N.E. El Gueddari and B.M. Moerschbacher, 2011. Chitinolytic enzymes from endophytic fungi. Fungal Diversity, 47: 43-53.
CrossRef - Gulati, R., R.K. Saxena and R. Gupta, 1997. A rapid plate assay for screening L-asparaginase producing micro-organisms. Lett. Applied Microbiol., 24: 23-26.
CrossRefPubMedDirect Link - Gunatilaka, A.A.L., 2006. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity and implications of their occurrence. J. Nat. Prod., 69: 509-526.
CrossRefDirect Link - Heinemann, B. and A.J. Howard, 1969. Production of tumor-inhibitory L-asparaginase by submerged growth of Serratia marcescens. Applied Microbiol., 18: 550-554.
PubMed - Heinemann, B., A.J. Howard and H.J. Palocz, 1970. Influence of dissolved oxygen levels on production of L-asparaginase and prodigiosin by Serratia marcescens. Applied Environ. Microbiol., 19: 800-804.
PubMedDirect Link - Kumar, S., V.V. Dasu and K. Pakshirajan, 2010. Localization and production of novel L-asparaginase from Pectobacterium carotovorum MTCC 1428. Process Biochem., 45: 223-229.
CrossRef - Kumar, S., V.V. Dasu and K. Pakshirajan, 2011. Purification and characterization of glutaminase-free L-asparaginase from Pectobacterium carotovorum MTCC 1428. Bioresour. Technol., 102: 2077-2082.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mahajan, R.V., S. Saran, K Kameswaran, V. Kumar and R.K. Saxena, 2012. Efficient production of L-asparaginase from Bacillus licheniformis with low-glutaminase activity: Optimization, scale up and acrylamide degradation studies. Bioresour. Technol., 125: 11-16.
CrossRef - Maladkar, N.K., V.K. Singh and S.R. Naik, 1993. Fermentative production and isolation of L-asparaginase from Erwinia carotovora, EC-113. Hindustan Antibiot. Bull., 35: 77-86.
PubMedDirect Link - Manna, S., A. Sinha, R. Sadhukhan and S.L. Chakrabarty, 1995. Purification, characterization and antitumor activity of L-asparaginase isolated from Pseudomonas stutzeri MB-405. Curr. Microbiol., 30: 291-298.
CrossRefDirect Link - Mashburn, L.T. and Jr. J.C. Wriston, 1964. Tumor inhibitory effect of L-asparaginase from Escherichia coli. Arch. Biochem. Biophys., 105: 450-453.
PubMed - Mohan Kumar, N.S., R. Ramasamy and H.K. Manonmani, 2013. Production and optimization of L-asparaginase from Cladosporium sp. using agricultural residues in solid state fermentation. Ind. Crops Prod., 43: 150-158.
CrossRef - Mostafa, S.A. and M.S. Salama, 1979. L-Asparaginase-producing Streptomyces from the soil of Kuwait. Zentralblatt Bakteriologie Parasitenkunde Infektionskrankheiten Hygiene, 134: 325-334.
CrossRef - Muller, H.J. and J. Boos, 1998. Use of L-asparaginase in childhood ALL. Crit. Rev. Oncol. Hematol., 282: 97-113.
PubMedDirect Link - Narayana, K.J.P., K.G. Kumar and M. Vijayalakshmi, 2008. L-asparaginase production by Streptomyces albidoflavus. Indian J. Microbiol., 48: 331-336.
CrossRefDirect Link - Pedreschi, F., K. Kaack and K. Granby, 2008. The effect of asparaginase on acrylamide formation in French fries. Food Chem., 109: 386-392.
CrossRefDirect Link - Pieters, R., S.P. Hunger, J. Boos, C. Rizzari and L. Silverman et al., 2011. L-asparaginase treatment in acute lymphoblastic leukemia: A focus on Erwinia asparaginase. Cancer, 117: 238-249.
CrossRefDirect Link - Rocha, A.C., D. Garcia, A.P. Uetanabaro, R.T. Carneiro, I.S. Araujo, C.R. Mattos and A. Goes-Neto, 2011. Foliar endophytic fungi from Hevea brasiliensis and their antagonism on Microcyclus ulei. Fungal Diversity, 47: 75-84.
CrossRef - Rytting, M.E., 2012. Role of L-asparaginase in acute lymphoblastic leukemia: Focus on adult patients. Blood Lymphatic Cancer: Targets Therapy, 2: 117-124.
Direct Link - Sarquis, M.I.M., E.M.M. Oliveira, A.S. Santos and G.L. Da Costa, 2004. Production of L-asparaginase by filamentous fungi. Mem. Inst. Oswaldo Cruz., 99: 489-492.
CrossRefDirect Link - Savitri, N.A. and W. Azmi, 2003. Microbial L-asparaginase: A potent antitumour enzyme. Indian J. Biotechnol., 2: 184-194.
Direct Link - Shaffer, P.M., H.N. Arst, L. Estberg, L. Fernando, T. Ly and M. Sitter, 1988. An asparaginase of Aspergillus nidulans is subject to oxygen repression in addition to nitrogen metabolite repression. Mol. Gen. Genet. MGG, 212: 337-341.
CrossRef - Sudhir, A.P., B.R. Dave, K.A. Trivedi and R.B. Subramanian, 2012. Production and amplification of an L-asparaginase gene from actinomycete isolate Streptomyces ABR2. Ann. Microbiol., 62: 1609-1614.
CrossRef - Sukumaran, C.P., D.V. Singh and P.R. Mahadevan, 1979. Synthesis of L-asparaginase by Serratia marcescens (Nima). J. Biosci., 1: 263-269.
CrossRef - Suryanarayanan, T.S., N. Thirunavukkarasu, M.B. Govindarajulu and V. Gopalan, 2012. Fungal endophytes: An untapped source of biocatalysts. Fungal Diversity, 54: 19-30.
CrossRef - Suryanarayanan, T.S., N. Thirunavukkarasu, M.B. Govindarajulu, F. Sasse, R. Jansen and T.S. Murali, 2009. Fungal endophytes and bioprospecting. Fungal Biol. Rev., 23: 9-19.
CrossRefDirect Link - Suryanarayanan, T.S., V. Kumaresan and J.A. Johnson, 1998. Foliar fungal endophytes from two species of the mangrove Rhizophora. Can. J. Microbiol., 44: 1003-1006.
CrossRef - Thirunavukkarasu, N., T.S. Suryanarayanan, T.S. Murali, J.P. Ravishankar and S.N. Gummadi, 2011. L-asparaginase from marine derived fungal endophytes of seaweeds. Mycosphere, 2: 147-155.
Direct Link - Vega, F.E., 2008. Insect pathology and fungal endophytes. J. Invertebr. Pathol., 98: 277-279.
CrossRefDirect Link - Wei, D.Z. and H. Liu, 1998. Promotion of L-asparaginase production by using n-dodecane. Biotechnol. Techniques, 12: 129-131.
CrossRef