
Parvaze Ahmad Wani
Department of Biological Sciences, College of Natural and Applied Sciences, Crescent University, Abeokuta, Ogun State, Nigeria
Mohammad Saghir Khan
Department of Agricultural Microbiology, Faculty of Agricultural Sciences, Aligarh Muslim University, Aligarh, UP, India
Research Journal of Microbiology
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 25-35







ABSTRACT
Heavy metal contamination has accelerated due to the rapid industrialization world wide. Accumulation of metals in excess can modify the structure of essential protein or can replace an essential element. Aim of this study is to check the Mesorhizobium species for their tolerance towards multiple metals as well as antibiotics and further check whether these metal resistant microbes are producing any plant growth promoting substances. Mesorhizobium strains were isolated from the nodules of chickpea, were tested for their tolerance to both heavy metals and antibiotics by agar plate dilution method and by the disc diffusion method respectively. Plant growth promoting activity was checked by the standard methods. All of the isolates showed tolerance to cadmium, chromium, nickel, lead, zinc and copper. All the isolates showed maximum tolerance towards lead and zinc which was followed by nickel and chromium. Mesorhizobium species also showed tolerance towards antibiotics, 70% of the isolates were tolerant to nalidixic acid, 60% were tolerant to each tetracycline and nitrofurantoin whereas no isolate was found to be resistant towards cloxacillin, ciprofloxacin and chloramphenicol. Mesorhizobium strains were also tested for their Plant Growth Promoting (PGP) substances, all the isolates produced good amount of indole acetic acid and were positive for ammonia and Hydrogen Cyanide (HCN) but only three isolates (RC1, RC3 and RC4) produced siderophore on the plates as well as in the liquid culture. In conclusion Mesorhizobium strains can be used for the growth promotion, as well for the detoxification of the heavy metals in metal polluted soils.
PDF Abstract XML References Citation
Received: January 19, 2013;
Accepted: March 11, 2013;
Published: May 29, 2013
How to cite this article
Parvaze Ahmad Wani and Mohammad Saghir Khan, 2013. Isolation of Multiple Metal and Antibiotic Resistant Mesorhizobium
species and Their Plant Growth Promoting Activity. Research Journal of Microbiology, 8: 25-35.
URL: https://scialert.net/abstract/?doi=jm.2013.25.35
URL: https://scialert.net/abstract/?doi=jm.2013.25.35
INTRODUCTION
Contamination of the environment by heavy metals is the wide spread problem due to their use in industries and agricultural purposes (Fernandes and Henriques, 1991). It adversely affect about 12% of the world’s agricultural land (Moffat, 1999). Heavy metal pollution has increased from the start of the industrial revolution. The primary source of this pollution includes the industrial operations such as, smelting, mining, metal forging, manufacturing of alkaline storage batteries, combustion of fossil fuel and sewage sludge of industrial/domestic origin (Ibekwe et al., 1995). The application of sewage sludge in agronomic practices is often the most economical means of disposal. It is beneficial because it increases the organic matter content and water holding capacity of soil (Pagliai et al., 1981) and also provides plant with sufficient nutrients. However, sewage sludges from industrial sources, often contain variable amounts of potentially toxic heavy metals, such as, lead, cadmium, nickel, chromium, copper and zinc (McGrath, 1987). When these sludges are repeatedly applied to agronomic lands, heavy metals accumulate and persist in the top cultivated layer (0-20 cm) (McGrath, 1987). The persistence of these metals in soil adversely affect the agro-ecosystem (Mcllveen and Negusanti, 1994; Broos et al., 2004, 2005). Despite the reports of availability of larger quantities of heavy metals in sewage water, it is widely applied in agronomic practices for irrigation purposes. Though, a large number of reports on the effects of sewage sludge having multiple metals on microbial communities (McGrath et al., 1988; Giller et al., 1998) and plants (Ibekwe et al., 1995) are available, yet there is discrepancy in the reported results (Ramirez et al., 2008). And hence, a firm conclusion on the toxicity of heavy metals on plants including legumes and their symbiotic partners or plant growth promoting rhizobacteria can not be drawn. Moreover, the majority of the adverse effects observed in sludge treated soils are possibly due to the factors other than metals (e.g., contaminants, excess N supply) which increases the toxicity.
The elevated concentration of such metals adversely affects the quantitative and qualitative composition of microbial communities in soil including those bacterial populations that aggressively colonize plant roots and termed as Plant Growth Promoting Rhizobacteria (PGPR) (Kloepper and Schroth, 1978), leading to an altered microbial equilibrium in rhizosphere (Gray and Smith, 2005). Also, the enhanced concentrations of metals affect growth, metabolisms and consequently the total biomass of naturally occurring beneficial microbes.
To survive under metal stress conditions, plant growth promoting bacteria have evolved several mechanisms to tolerate the uptake of heavy metal ions (Nies, 1999). These mechanisms include precipitation of metal as insoluble salts by chemical transformation, accumulation and sequestration of the metal ions inside the cell, biotransformation-transformation of toxic metal to less toxic forms (Thacker and Madamwar, 2005). Almost all known bacterial resistance mechanisms are encoded on plasmids and transposons (Silver and Walderhaug, 1992) and the bacteria acquire resistance to heavy metals either by the gene transfer or spontaneous mutation. Metal ions are known to cause oxidative stress, stress response genes are induced as metal ion concentrations increase from starvation to toxic level. It is studied that genes are expressed under specific metal stress (Singh et al., 2001). For example, in Gram-negative bacteria (e.g., Ralstonia eutropha), the czc system is responsible for the resistance to Cd, Zn and Co. The czc-genes encode for a cation-proton antiporter (CzcABC) which exports Cd, Zn and Co. Similarly in Alcaligenes xylosoxidans, ncc system was found which is resistant to Ni, Cd and Co. On the contrary, Cd-efflux ATPase was responsible for Cd resistance in Gram-positive bacteria (e.g., Staphylococcus, Bacillus or Listeria). Cu resistance systems observed in Pseudomonas syringae pv. tomato is cop whereas, in Escherichia coli it is pco which encode for different Cu-binding proteins which allow Cu to bind in the periplasm or in the outer membrane. In contrast, the pco system is an ion-dependent Cu antiporter (Kunito et al., 1997). Additionally, a few naturally occurring microorganisms with high metal resistance in plant rhizosphere significantly reduced metal toxicity (Kamaludeen and Ramasamy, 2008). Generally, plant growth promoting rhizobacteria release plant growth promoting substances (Sheng and Xia, 2006; Ahmad et al., 2008), vitamins, enzymes, siderophores and antibiotics (Noordman et al., 2006; Burd et al., 2000) which result in the over all improvement in the growth of the plants. They also promote the growth of plants by alleviating the stress induced by ethylene-mediated impact on plants by synthesizing 1-aminocyclopropane-1-carboxylate (ACC) deaminase (Madhaiyan et al., 2007; Glick et al., 2007). The present study was therefore, undertaken to check the resistance pattern of plant growth promoting rhizobacteria to heavy metals and antibiotics and their phyhormone production.
MATERIALS AND METHODS
Collection of samples: Plant samples for the isolation of bacteria were collected from the soils of the experimental sites of Faculty of Agricultural Sciences, Aligarh Muslim University Aligarh, U.P., India.
Evaluation of bacterial strains for metal tolerance: Mesorhizobiuu strains were isolated from the nodules of chickpea plants grown in the soils of the experimental sites of Faculty of Agricultural Sciences, Aligarh Muslim University Aligarh, U.P, India. Mesorhizobium strains were tested for their resistance to six metals like cadmium, chromium, nickel, lead, zinc and copper not by agar plate dilution method (Holt et al., 1994) using YEM agar medium. The freshly prepared agar plates amended with increasing concentration of cadmium (0-400 μg mL-1), chromium (0-500 μg mL-1), nickel (0-500 μg mL-1), lead (0-1500 μg mL-1), zinc (0-1500 μg mL-1) and copper (0-400 μg mL-1) were spot inoculated (100 μL) with 108 cells mL-1. Plates were incubated at 28±2° for 3-5 days. Lowest concentration of metals inhibiting bacterial growth on nutrient agar plate was defined as a minimum inhibitory concentration. Each experiment was replicated three times.
Determination of antibiotic resistance: To determine resistance to antibiotics, the plant growth promoting Mesorhizobium were tested for their sensitivity to ten antibiotics. The reactions to antibiotics were determined by the disc diffusion method (Bauer et al., 1966). Mesorhizobium species were grown in Yeast Extract Mannitol (YEM) broth respectively, at 28±2°C for 24 h. A 0.1 mL of the over night grown culture was spread on the surface of yeast extract mannitol agar. The antibiotic discs of known potency were then placed on the agar surface and the plates were incubated at 28±2°C for 24 h and the zones of inhibition around the antibiotic discs were measured to the following antibiotics (all from Hi-media Mumbai): ampicillin (25 μg), chloramphenicol (25 μg), ciprofloxacin (30 μg), cloxacillin (30 μg), gentamycin (30 μg), methicillin (30 μg), nalidixic acid (30 μg), nitrofurantoin (100 μg), streptomycin (25 μg) and tetracycline (30 μg).
Plant growth promoting activities: Indole-3-acetic acid was quantitatively assayed by the method of Gordon and Weber (1951) later modified by Bric et al. (1991). For the activity of indole acetic acid bacteria were grown in Luria Bertani (LB) broth (g L-1: tryptone 10; yeast extract 5; Nacl 10 and pH 7.5). Luria Bertani broth (100 mL) supplemented with 0, 20, 40, 60, 80 and 100 μg mL-1 of tryptophan was inoculated with 108 cells mL-1 of one mL culture and was incubated for 24 h at 28±2°C with shaking at 125 rpm. After 24 h, 5 mL of each culture was centrifuged (10,000 rpm) for 15 min and an aliquot of 2 mL supernatant was mixed with 100 μL of orthophosphoric acid and 4 mL of Salkowsky reagent (2% 0.5 M FeCl3 in 35% per-chloric acid) and incubated at 28±2°C in darkness for 1 h. The absorbance of pink color developed was read at 530 nm. The Indole Acetic Acid (IAA) concentration in the supernatant was determined using a calibration curve of pure IAA as a standard (Bric et al., 1991). The experiments were repeated three times on different time intervals.
The bacterial strains were further assayed for siderophore production on the Chrome Azurol S (CAS) agar medium by the method of Alexander and Zuberer (1991). Chrome Azurol S agar plates were prepared separately and divided into equal sectors and spot inoculated with 100 μL of 108 cells mL-1 and incubated at 28±2°C for 72-96 h. Development of yellow orange halo around the growth was considered as positive for siderophore production. Each individual experiment was replicated three times. The siderophore produced by the test strains were further quantitatively assayed using Modi medium (K2 HPO4 0.05%; MgSO4 0.04%; NaCl 0.01%; mannitol 1%; glutamine 0.1%; NH4NO3 0.1%). Modi medium was inoculated with 108 cells mL-1 of bacterial cultures and incubated at 28±2°C for 5 days. Catechol type phenolates were measured on ethyl acetate extracts of the culture supernatant using a modification of the ferric chloride-ferricyanide reagent of Hathway. Ethyl acetate extracts were prepared by extracting 20 mL of supernatant twice with an equal volume of solvent at pH 2. Hathway,s reagent was prepared by adding 1 mL of 0.1 M ferric chloride in 0.1 N HCl to 100 mL of distilled water and to this was then added 1 mL 0.1 M potassium ferricyanide (Reeves et al., 1983). For the assay, one volume of the reagent was added to one volume of sample and absorbance was determined at 560 nm for salicylates with sodium salicylate as standard and at 700 nm for dihydroxy phenols with 2, 3-dihydroxy benzoic acid as standard. Hydrogen cyanide production by bacterial isolates was detected by the method of Bakker and Schippers (1987). For HCN production, the bacterial strains were grown on an HCN induction medium (30 g tryptic soy broth, 4.4 g glycine, 15 g agar L-1) at 28±2°C for four days. For each bacterial isolate, 100 μL of 108 cells mL-1 was placed in the centre of the petri plates. A disk of Whatman filter paper No. 1 dipped in 0.5% picric acid and 2% Na2CO3 was placed at the lid of the petri plates. Plates were sealed with parafilm. After four days incubation at 28±2°C, an orange brown colour of the paper indicating HCN production was observed. For ammonia production, the rhizobial strains were grown in peptone water (g L-1: peptone 10 g, NaCl 5 g, pH 7) and incubated at 30±2°C for four days. One milliliter of Nessler reagent was added to each tube and the development of yellow color indicating ammonia production was recorded (Dye, 1962).
Statistical analysis: Data of three replicates were subjected to statistical analysis using pair samples T test with significance level of p<0.05. The values indicate the Mean±SD of three replicates.
RESULTS
Tolerance of plant growth promoting rhizobacteria to metals: The selected plant growth promoting rhizobacterial strains were tested for their ability to tolerate various concentrations of heavy metals like cadmium, chromium, nickel, lead, zinc and copper using agar plate dilution method. Generally, the Plant Growth Promoting Rhizobacterial (PGPR) strains showed a varied level of tolerance to heavy metals. Among the Mesorhizobium strains, strain RC3 showed highest tolerance to most of the metals (Fig. 1). Strain RC3 tolerated a concentration of 400, 500, 500, 1500, 1500 and 400 μg mL-1 of cadmium, chromium, nickel, lead, zinc and copper, respectively, amended in agar plates whereas strain RC4 showed a tolerance level of 400, 400, 400, 1400, 1400 and 300 μg mL-1 to cadmium, chromium, nickel, lead, zinc and copper, respectively, added to solid plates.
Antibiotic Resistance of plant growth promoting rhizobacteria: Resistance to antibiotics among metal tolerant rhizobacterial strains differed considerably (Table 1). Among Mesorhizobium spp., 70% of strains were resistant to nalidixic acid, 60% to tetracycline and nitrofurantoin while 30% were resistant to each streptomycin and methicillin. No strain was found to be resistant to cloxacillin, ciprofloxacin and chloramphenicol.
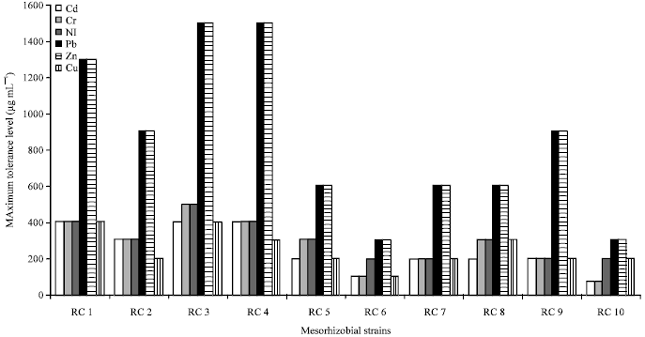 | |
| Fig. 1: | Maximum tolerance level of Mesorhizobium strains to different heavy metals |
| Table 1: | Resistant pattern of Mesorhizobium species to various antibiotics |
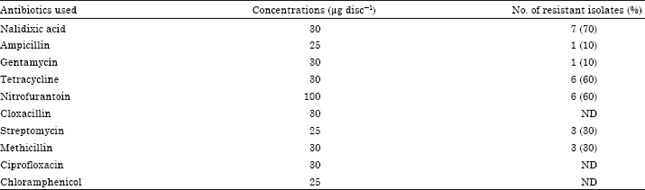 | |
| ND: Not detected, Total number of isolates = 10 | |
Bioassay of plant growth promoting activities: The Plant Growth Promoting (PGP) substances like IAA, siderophore, hydrogen cyanide and ammonia synthesized by the metal tolerant PGPR strains were assayed both qualitatively and quantitatively under in vitro experiments and are explained as follows:
Indole acetic acid: The production of IAA by the metal tolerant bacterial strains was assayed in LB broth supplemented with different concentrations of tryptophan (Table 2). The Mesorhizobium spp. exhibited a substantial production of IAA after 24 h of incubation. Moreover, the data revealed a concentration dependent increase in IAA, the maximum being 34.5, 30.6, 27.9, 26.5, 23.5 and 10.9 μg of IAA/mL in LB broth supplemented with 100, 80, 60, 40, 20 and 0 (without tryptophan) μg tryptophan/mL, respectively, by strain RC3. This was followed by strain RC4 which produced a maximum amount of 34.2, 29, 27.3, 26.1, 23.1 and 10.4 μg IAA/mL in LB broth supplemented with 100, 80, 60, 40, 20 and 0 μg tryptophan/mL, respectively.
| Table 2: | Plant growth promoting activities of Mesorhizobium species (N = 10) isolated from chickpea nodules |
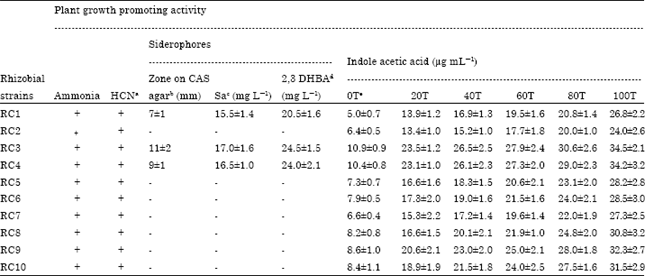 | |
| a: Hydrogen cyanide, b: Chrome azurol S agar, c: Salicylic acid, d: 2,3 Dihydroxy benzoic acid, e: Tryptophan concentration (mg mL-), +: Positive reaction, - : No reaction: The values in this table indicate the Mean±SD of three replicates | |
While comparing the effect of various concentrations of tryptophan on IAA production by the Mesorhizobial strains, 100 μg mL-1 tryptophan showed a significant (p≤0.05) increase of 229 and 18% in IAA over 20 and 80 μg mL-1 tryptophan, respectively, by the strain RC4.
Bioassay of siderophore: Another important trait of plant growth promoting rhizobacteria is the production of siderophores that may indirectly affect the growth of plants. In the present investigation, the metal tolerant PGPR strains were tested for both qualitative and quantitative production of siderophores using CAS agar and ethyl acetate extraction method (Table 2). On CAS agar plates, 30% of the Mesorhizobium strains produced siderophore. Of these strains, RC1, RC3 and RC4 displayed 7, 11 and 9 mm colored zone on CAS plates after four days of incubation. Further, the ethyl acetate extraction from culture supernatant of Mesorhizobium strain RC1 yielded 15.5 and 20.5 mg mL-1 salicylate (SA) and 2,3-dihydroxy benzoic acid (DHBA), strain RC3 produced 17 and 24.5 mg mL-1 of SA and DHBA and strain RC4 yielded 16.5 and 24 mg mL-1 SA and DHBA, respectively (Table 2).
In vitro assay of ammonia and HCN: The metal tolerant plant growth promoting rhizobacterial strains were tested further for the synthesis of ammonia and hydrogen cyanide using peptone water and HCN induction medium, respectively. Generally, all PGPR strains were found positive for ammonia and HCN (Table 2).
DISCUSSION
Deposition of metal into soil over a long period of time results in high concentration of metal in the soil which adversely affects the microflora of the soil (Matsuda et al., 2002). Heavy metals in general show adverse effect on the soil microbial flora by blocking their functional groups or these metals modify the biological molecules in particular their active sites. But these metals when present in low concentrations are important for the microbes as they supply the microorganisms with the essential co-factors for metallo proteins and enzymes (Nies, 1999). The metal- microbe interaction in natural environment is complex and is influenced by pH or organic matter content (Saeki et al., 2002). The ability to grow at concentration of metals is however, found in many plant growth promoting rhizobacteria including symbiotic nitrogen fixing bacteria (Lakzian et al., 2002) and may be the result of intrinsic or induced mechanism (Giller et al., 1998). There are reports that have shown a high level tolerance to heavy metals by rhizobia (Wani and Khan, 2010). Conflicting reports are, however, available in the literature on the tolerance level of rhizobia which could possibly be due to the variation in the tolerance level of rhizobia and growth conditions employed (Rajkumar et al., 2005). For instance, Rhizobium leguminosarum isolated from metal contaminated soil tolerated 92.9 μM of zinc (Delorme et al., 2003) while Rhizobium species isolated from nodules of Trifolium repense tolerated 300 mg kg-1 nickel and showed an effective symbiosis with its legume host, when grown in nickel amended soils (Smith and Giller, 1992). Similarly Luo et al. (2011), isolated Serratia sp. LRE07 from cadmium hyperaccumulator Solanum nigrum L. was resistant to to the toxic effects of heavy metals.
In the present study, Mesorhizobium RC3 isolated from chickpea nodules, displayed a high resistance towards cadmium (400 μg mL-1), Chromium (500 μg mL-1), nickel (500 μg mL-1), Lead (1500 μg mL-1) , zinc (1500 μg mL-1) and copper (400 μg mL-1) whereas strain RC4 showed a tolerance level of 400, 400, 400, 1400, 1400 and 300 μg mL-1 to cadmium, chromium, nickel, lead, zinc and copper, respectively. Mesorhizobium strains showed a high tolerance to lead and zinc which was followed for chromium, whereas cadmium was found to be more toxic than the other heavy metals. The metal tolerant strains were characterized by physiological and biochemical methods. In other studies, the Gram-negative bacteria have also shown resistance to cadmium and zinc salts. For instance, cadmium and zinc tolerance by Protobacteria, Actinobacteria and Bacteroidetes were resistant to Zn and Cadmium (Kuffner et al., 2010). Nickel and zinc tolerance by Rhizobium leguminosarum biovar trifolii isolated from sewage sludge treated soil was also reported by Purchase and Miles (2001), who observed a metal tolerance of 0.24-0.26 mM Ni2+ and 6.0-8.0 mM Zn2+. Similarly, metal tolerance by Rhizobium, Bradyrhizobium and Azotobacter (Pajuelo et al., 2008) and varying level of resistance among other PGPR (Bacillus and Pseudomonas) have also been reported (Yilmaz, 2003; Thacker et al., 2007; Wasi et al., 2008).
Bacterial resistance to antibiotics is an emerging problem these days. Resistance to antibiotics is acquired by a change in the genetic make up of microbes which can occur by either a genetic mutation or by transfer of antibiotic resistant genes between organisms in the environment (Spain and Alm, 2003). Furthermore, the increased use of antibiotics in health care as well as in agriculture, is in turn contributing to the growing problems of antibiotic resistant bacteria. Products such as heavy metals used in industry along with antibiotics create a selective pressure in the environment that consequently leads to the mutation in organism that will allow them better to survive and multiply. Clustering of genes on a plasmid, are beneficial to the survival of that organism and its species because those genes are more likely to be transferred together in the event of conjugation. Thus, in an environment with multiple stresses, for example antibiotics and heavy metals, it would be more ecologically favorable in terms of survival for a bacterium to acquire resistance to both stresses. If the resistance is plasmid mediated, bacteria harbouring clustered genes are more likely to pass on those genes to other neighbouring bacteria which would then have a better chance of survival. With these considerations, the antibiotic resistance among PGPR was studied which differed from antibiotic to antibiotic for all the PGPR strains. Multiple antibiotic resistances shown by PGPR strains (e.g., Mesorhizobium RC3, RC4) might be associated with a high degree of tolerance to metals. In many studies, metal tolerance and antibiotic resistance have been reported (Wani et al., 2009; Yilmaz, 2003; Verma et al., 2001). It has been suggested that under environmental conditions of metal stress, metal and antibiotic resistant microorganisms will adapt faster by the spread of R-factors than by mutation and natural selection (Silver and Misra, 1988). Similar observations on antibiotics resistance by PGPR strains have been reported (Thacker et al., 2007). The variation in the resistance to many tested antibacterial drugs (antibiotics) may possibly be due to the differences in growth conditions and exposure of PGPR to stress conditions or toxic substance as well as presence or absence of resistance mechanisms that could be encoded either by chromosome and/or R-plasmid (Spain and Alm, 2003).
In the present study Mesorhizobium strains were positive for plant growth promoting activities and produced substantial amount of IAA, siderophore, HCN and ammonia. Bacterial strains produce ammonia; this ammonia plays a signalling role when the plant growth promoting bacteria and plants interact with each other (Becker et al., 2002). Moreover, the ammonia released by the bacterial strains are known to increase the glutamine synthetase activity (Chitra et al., 2002). In addition, ammonium transporters found in several plant growth promoting rhizobacteria are thought to be involved in the reabsorption of NH4+ released as a consequence of NH3 diffusion through the bacterial membrane (Van Dommelen et al., 1997). Similarly phytohormone production (Wani and Khan, 2010; Rajkumar et al., 2006; Ahmad et al., 2008) and IAA by rhizobia is reported (Antoun et al., 1998). Siderophores is also known to bind to the available form of iron Fe3+ in the rhizosphere thus making it unavailable to the phytopathogens and consequently protects the plant health. In other study, the heavy metal resistant Bacillus species are also known to produce considerable amount of plant growth promoting substances (Wani et al., 2007).
CONCLUSION
This study concluded that the Mesorhizobium strains not only tolerated heavy metals but also antibiotics. Mesorhizobium also produced substantial amount of plant growth promoting substances. Due to multifarious properties expressed by the Mesorhizobium strains, these strains could therefore, be used as bioinoculant to increase the performance of crops in soils contaminated with heavy metals.
REFERENCES
- Ahmad, F., I. Ahmad and M.S. Khan, 2008. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res., 168: 173-181.
CrossRefDirect Link - Alexander, D.B. and D.A. Zuberer, 1991. Use of chrome azurol S reagents to evaluate siderophore production by Rhizosphere bacteria. Biol. Fertil. Soils, 12: 39-45.
CrossRefDirect Link - Antoun, H., C.J. Beachamp, N. Goussard, R. Chabot and R. Lalande, 1998. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effects on radishes (Rhaphanus sativus L.). Plant Soil, 204: 56-67.
Direct Link - Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Bakker, A.W. and B. Schippers, 1987. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas SPP-mediated plant growth-stimulation. Soil Biol. Biochem., 19: 451-457.
CrossRefDirect Link - Becker, D., R. Stanke, I. Fendrik, W.B. Frommer, J. Vanderleyden, W.M. Kaiser and R. Hedrich, 2002. Expression of the NH4+-transporter gene LEAMT1;2 is induced in tomato roots upon association with N2-fixing bacteria. Planta, 215: 424-429.
CrossRefPubMedDirect Link - Bric, J.M., R.M. Bostock and S.E. Silverstone, 1991. Rapid in situ assay for indoleacetic acid production by bacteria immobilized on a nitrocellulose membrane. Applied Environ. Microbiol., 57: 535-538.
PubMedDirect Link - Broos, K., M. Uyttebroek, J. Mertens and E. Smolders, 2004. A survey of symbiotic nitrogen fixation by white clover grown on metal contaminated soils. Soil Biol. Biochem., 36: 633-640.
CrossRef - Broos, K., H. Beyens and E. Smolders, 2005. Survival of rhizobia in soil is sensitive to elevated zinc in the absence of the host plant. Soil Biol. Biochem., 37: 573-579.
CrossRef - Burd, G.I., D.G. Dixon and B.R. Glick, 2000. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol., 46: 237-245.
CrossRefPubMedDirect Link - Sood, C.R., S.V. Chanda and Y.D. Singh, 2002. Effect of different nitrogen sources and plant growth regulators on glutamine synthetase and glutamate synthase activities of radish cotyledons. Bulg. J. Plant Physiol., 28: 46-56.
Direct Link - Delorme, T.A., J.V. Gagliardi, J.S. Angle, P.B. van Berkum and R.L. Chaney, 2003. Phenotypic and genetic diversity of rhizobia isolated from nodules of clover grown in a zinc and cadmium contaminated soil. Soil Sci. Soc. Am. J., 67: 1746-1754.
CrossRefDirect Link - Dye, D.W., 1962. The inadequacy of the usual determinative tests for the identification of Xanthomonas spp. N. Z. J. Sci., 5: 393-416.
Direct Link - Fernandes, J.C. and F.S. Henriques, 1991. Biochemical, physiological and structural effects of excess copper in plants. Bot. Rev., 57: 246-273.
CrossRefDirect Link - Giller, K.E., E. Witter and S.P. Mcgrath, 1998. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem., 30: 1389-1414.
CrossRefDirect Link - Glick, B.R., Z. Cheng, J. Czarny and J. Duan, 2007. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol., 119: 329-339.
CrossRef - Gordon, S.A. and R.P. Weber, 1951. Colorimetric estimation of indoleacetic acid. Plant Physiol., 26: 192-195.
PubMedDirect Link - Gray, E.J. and D.L. Smith, 2005. Intracellular and extracellular PGPR: Commonalities and distinctions in the plant-bacterium signaling processes. Soil Biol. Biochem., 37: 395-412.
CrossRefDirect Link - Ibekwe, A.M., J.S. Angle, R.L. Chaney and P. van Berkum, 1995. Sewage sludge and heavy metal effects on nodulation and nitrogen fixation of legumes. J. Environ. Qual., 24: 1199-1204.
Direct Link - Kamaludeen, S.P.B. and K. Ramasamy, 2008. Rhizoremediation of metals: Harnessing microbial communities. Indian J. Microbiol., 48: 80-88.
CrossRef - Kunito, T., K. Nagaoka, N. Tada, K. Saeki, K. Senoo, H. Oyaizu and S. Matsumoto, 1997. Characterization of Cu-resistant bacterial communities in Cu-contaminated soils. Soil Biol. Plant Nutr., 43: 709-717.
CrossRef - Lakzian, A., P. Murphy, A. Turner, J.L. Beynon and K.E. Giller, 2002. Rhizobium leguminosarum bv. viciae populations in soils with increasing heavy metal contamination: Abundance, plasmid profiles, diversity and metal tolerance. Soil Biol. Biochem., 34: 519-529.
Direct Link - Madhaiyan, M., S. Poonguzhali and T. Sa, 2007. Metal tolerating methylotrophic bacteria reduces nickel and cadmium toxicity and promotes plant growth of tomato (Lycopersicon esculentum L.). Chemosphere, 69: 220-228.
CrossRef - McGrath, S.P., P.C. Brookes and K.E. Giller, 1988. Effects of potentially toxic metals in soil derived from past applications of sewage sludge on nitrogen fixation by Trifolium repens L. Soil Biol. Biochem., 20: 415-424.
CrossRefDirect Link - McIlveen, W.D. and J.J. Negusanti, 1994. Nickel in the terrestrial environment. Sci. Total Environ., 148: 109-138.
CrossRef - Nies, D.H., 1999. Microbial heavy-metal resistance. Applied Microbiol. Biotechnol., 51: 730-750.
PubMedDirect Link - Noordman, W.H., R. Reissbrodt, R.S. Bongers, I.L.W. Rademaker, W. Bockelmann and G. Smit, 2006. Growth stimulation of Brevibacterium sp. by siderophores. J. Applied Microbiol., 101: 637-646.
CrossRef - Pagliai, M., G. Guidi, M. La Marca, M. Giachetti and G. Lucamante, 1981. Effects of sewage sludges and composts on soil porosity and aggregation. J. Environ. Qual., 10: 556-561.
CrossRefDirect Link - Pajuelo, E., I.D. Rodriguez-Llorente, M. Dary and A.J. Palomares, 2008. Toxic effects of arsenic on Sinorhizobium-Medicago sativa symbiotic interaction. Environ. Pollut., 154: 203-211.
CrossRefDirect Link - Rajkumar, M., R. Nagendran, K.J. Lee and W.H. Lee, 2005. Characterization of a novel Cr6+ reducing Pseudomonas sp. with plant growth-promoting potential. Curr. Microbiol., 50: 266-271.
CrossRefDirect Link - Rajkumar, M., R. Nagendran, K.J. Lee, W.H. Lee and S.Z. Kim, 2006. Influence of plant growth promoting bacteria and Cr6+ on the growth of Indian mustard. Chemosphere, 62: 741-748.
CrossRefDirect Link - Ramirez, W.A., X. Domene, O. Ortiz and J.M. Alcaniz, 2008. Toxic effects of digested, composted and thermally-dried sewage sludge on three plants. Bioresour. Technol., 99: 7168-7175.
CrossRef - Reeves, M.W., L. Pine, J.B. Neilands and A. Balows, 1983. Absence of siderophore activity in Legionella species grown in iron-deficient media. J. Bacteriol., 154: 324-329.
PubMedDirect Link - Saeki, K., T. Kunito, H. Oyaizu and S. Matsumoto, 2002. Relationships between bacterial tolerance levels and forms of copper and zinc in soils. J. Environ. Qual., 31: 1570-1575.
PubMed - Silver, S. and T.K. Misra, 1988. Plasmid-mediated heavy metal resistance. Annu. Rev. Microbiol., 42: 717-743.
Direct Link - Silver, S. and M. Walderhaug, 1992. Gene regulation of plasmid-and chromosome determined inorganic ion transport in bacteria. Microbiol. Rev., 56: 195-228.
Direct Link - Singh, S., A.M. Kayastha, R.K. Asthana, P.K. Srivastava and S.P. Singh, 2001. Response of Rhizobium leguminosarum to nickel stress. World J. Microbiol. Biotechnol., 17: 667-672.
CrossRef - Smith, S.R. and K.E. Giller, 1992. Effective Rhizobium leguminosarum biovar Trifolii present in five soils contaminated with heavy metals from long-term applications of sewage sludge or metal mine spoil. Soil Biol. Biochem., 24: 781-788.
CrossRefDirect Link - Spain, A. and E. Alm, 2003. Implications of microbial heavy metal tolerance in the environment. Rev. Undergraduate Res., 2: 1-6.
Direct Link - Thacker, U. and D. Madamwar, 2005. Reduction of toxic chromium and partial localization of chromium reductase activity in bacterial isolate DM1. World J. Microbiol. Biotechnol., 21: 891-899.
CrossRef - Thacker, U., R. Parikh, Y. Shouche and D. Madamwar, 2007. Reduction of chromate by cell-free extract of Brucella sp. isolated from Cr(VI) contaminated sites. Bioresour. Technol., 98: 1541-1547.
CrossRefPubMedDirect Link - Van Dommelen, A., E. van Bastelaere, V. Keijers and J. Vanderleyden, 1997. Genetics of Azospirillum brasilense with respect to ammonium transport, sugar uptake and chemotaxis. Plant Soil, 194: 155-160.
CrossRefDirect Link - Wani, P.A., M.S. Khan and A. Zaidi, 2007. Chromium reduction, plant growth promoting potentials and metal solubilization by Bacillus sp. isolated from alluvial soil. Curr. Microbiol., 54: 237-243.
CrossRef - Wani, P.A., A. Zaidi and M.S. Khan, 2009. Chromium reducing and plant growth promoting potential of Mesorhizobium species under chromium stress. Bioremediation J., 13: 121-129.
CrossRefDirect Link - Wani, P.A. and M.S. Khan, 2010. Bacillus species enhance growth parameters of chickpea (Cicer arietinum L.) in chromium stressed soils. Food Chem. Toxicol., 48: 3262-3267.
CrossRefDirect Link - Wasi, S., G. Jeelani and M. Ahmad, 2008. Biochemical characterization of a multiple heavy metal, pesticides and phenol resistant Pseudomonas fluorescens strain. Chemosphere, 71: 1348-1355.
CrossRefDirect Link - Yilmaz, E.I., 2003. Metal tolerance and biosorption capacity of Bacillus circulans strain EB1. Res. Microbiol., 154: 409-415.
CrossRefDirect Link
Carlos J. Becquer Granados Reply
I find very interesting this paper and I encourage authors to continue with this research. I have used rhizobia for PGPR purposes in cereals under stressing pedoclimatic factors and have obtained very promissing results.
Congratulations!