
S. Tiewcharoen
Department of Parasitology, Faculty of Medicine Siriraj Hospital, Mahidol University, 2 Prannok Road, Bangkok-noi, Bangkok, 10700, Thailand
J. Rabablert
Department of Biology, Faculty of Science, Silpakorn University, Sanamchan Palace, Ampour Mung, Nakhon Pathom, 73000, Thailand
V. Junnu
Department of Parasitology, Faculty of Medicine Siriraj Hospital, Mahidol University, 2 Prannok Road, Bangkok-noi, Bangkok, 10700, Thailand
Research Journal of Microbiology
Year: 2009 | Volume: 4 | Issue: 9 | Page No.: 320-333







ABSTRACT
The objective of this study was to study the susceptibility of Naegleria fowleri trophozoites to Amphotericin B-combined chlorpromazine investigated the activities of single drugs used in combination with amphotericin B compared to those of each drug alone in vitro. The 50% inhibitory concentrations (IC50) and 100% minimal concentrations (MIC100) were calculated for single drugs and the drugs combination with fixed combination ratios of IC50 of amphotericin B. Single drugs, amphotericin B had the best IC50 and MIC100 scores against N. fowleri trophozoites. chlorpromaxine, Artesunate and azitromycin had following IC50 and MIC100 scores against trophozoites. However, we found that chlorpromazine in combination with amphotericin B was the best synergistic drug against N. fowleri trophozoites. According to single drugs, chlorpromaxine, artesunate and azitromycin plus amphotericin B had also been synergistic drugs against N. fowleri trophozoites. It was suggested that the combined use of these agents may be beneficial in treating Primary amoebic meningoencephalitis.
PDF Abstract XML References Citation
How to cite this article
S. Tiewcharoen, J. Rabablert and V. Junnu, 2009. In vitro Susceptibility of Naegleria fowleri Trophozoites to Amphotericin B-combined Chlorpromazine. Research Journal of Microbiology, 4: 320-333.
URL: https://scialert.net/abstract/?doi=jm.2009.320.333
URL: https://scialert.net/abstract/?doi=jm.2009.320.333
INTRODUCTION
Free living amoebae of the genus Naegleria are found in a variety of natural environments including soil, ponds and fresh water and feed on bacteria present in these environments (Ma et al., 1990). Several species have so far been identified, but only Naegleria fowleri has known to be an opportunistic pathogen, which causes primary amoebic meningoencephalitis, a severe disease of the central nervous system in human and mammals (John, 1982; Marcino-Cabral, 1988). Infection occurs usually by inhalation or aspirating of water contaminated with the amoeba trophozoites. The invasive amoebae enter the nervous system through the olfactory nerve and cause extensive inflammation, hemorrhage and necrosis in brain by disrupting neuronal tissues and other mammalian cells. Usually, the infection is fatal, resulting in death on 3 to 7 days after infection (Ma et al., 1990; Shin and Ki, 2004).
The reports of successful treatment of PAM involved a 9 year old girl who was diagnosed early in the disease course and treated with intravenous and intrathecal amphotericin B, intravenous and intrathecal miconazole and oral rifampin. The patient survived with minimal neurologic sequelae (Schuster and Rechthand, 1975). Additionally, the clinical use of amphotericin B in other patients is limited by a high frequency of renal toxicity and several other adverse effects (McCurdy et al., 1968; Goodman and Koenig, 1970; Proffitt et al., 1991). Moreover, not all patients treated with amphotericin B survived from PAM (Cursons et al., 2003) due to several problems, one of the most important of which is finding therapeutic agents for clinician management. In addition to amphotericin B, the sensitivities of other drugs such as artemisinin, azithromycin, chlorpromazine, clotrimazole, fluconazole, itraconazole, ketoconazole, metragainst to N. fowleri had also been evaluated (Tiewcharoen et al., 2002; Goswick and Brenner, 2003; Schuster and Visvesvara, 2004; Vargas-Zepeda et al., 2005; Ondarza et al., 2006, 2007; Soltow and Brenner, 2007).
It had been reported that interferon-α (IFN-α) and the nucleoside analog, acyclovir inhibited herpes simplex virus type 1 (HSV-1) replication. IFN-α and acyclovir can cross the blood brain barrier (Liu, 2005; Anand and Mitra, 2009). The IFN-α inhibited both DNA polymerase and alkaline DNase whereas acyclovir inhibited only DNA polymerase in post-transcription mechanism (Taylor et al., 1998). No reports of IFN-α and acyclovir against N. fowleri were previously studied. Our group was interested to study the susceptibility of N. fowleri trophozoites to sensitivities of both drugs compared with antimicrobial and antipsychotic drugs either in drug alone or drug combination with Amphotericin B.
MATERIALS AND METHODS
Naegleria fowleri Cultivation
This research project was conducted from October 2008 to April 2009. Trophozoites of N. fowleri (strain Siriraj) were isolated from a clinical specimen of a patient from Siriraj Hospital in 1986. The N. fowleri trophozoites were grown axenically in Nelson medium supplemented with 5% FCS. Cultures were incubated at 37°C in T-75 cm2 tissue culture flasks (Corning, USA). Trophozoites were harvested after 3-5 days during the logarithmic phase of growth by chilling the culture tubes in an ice-water bath for 15 min. After centrifugation at 2000 rpm for 5 min, the pellets were resuspended in culture medium and culture as previously described (Tiewcharoen et al., 2008).
Pharmaceutical Agents
Drugs used in the study could be categorized to be five groups as followed. Group I, Antifungal drugs; Amphotericin B (Abbott, Saint-Remy-sur-Avre, France, S.A.; MW 924.1: C47H73NO17), Fluconazole (Siam Bheasach, Bangkok, Thailand; MW 306.3: C13H12F2N6O) and Voricinazole (Vfend (R); Roerig/Pfizer Inc., NewYork, NY; MW 349.3105: C16H14F3N5O), Group II, Antibacterial drugs; azithromycin (Ben Venue Laboratories, Ohio, USA; MW 748.984, C38H72N2O12), Gentamicin sulfate (T.P. Drug Laboratories, Bangkok, Thailand; MW 477.59: C21H43N5O7) and Ciprofloxacin (Ranbaxy laboratories limited, Secunderabad, India; MW 331.4: C17H18FN3O3). Group III, Antiprotozoal drugs; Artesunate (Gulin Pharmaceutical Organization, Guangxi, China; MW 384.42: C19H28O8), Quinine dihydrochlorinate (Guangzhou Person Pharmaceutical Organization, China; MW 396.90: C20H24N2O2•HCl•2H2O) and Metronidazole (Biolab, Bangkok, Thailand; MW 171.16: C6H9 N3O3). Group IV, Antiviral drug; acyclovir (Mayne Pharma Pty Ltd., Mulgrave, Australia; MW 225: C8H11N5O3). Group V, Antiphychotic drug; Chlorpromazine hydrochloride (The Government. Pharmaceutical Organization, Bangkok, Thailand; MW 355.3: C17H19ClN2S). Group VI, Interferon-alph (IFN-α2b, MW 19.5 kDa, Reliance Biopharmaceuticals, Argentina). Pharmaceutical agents were dissolved in and diluted with sterile distilled water according to the manufacturer’s specifications to provide the concentrations and doses used in this study.
Morphology and Growth of N. fowleri Trophozoites
One mililiters of the trophozoites resuspended in Nelson medium containing 5% FCS (1x104 cells) were cultured in each well of 12 well tissue culture plates (Corning). Then, the plates were sealed and incubated at 37°C. Tests were performed in duplicate and were repeated at least three times. Trophozoites were observed for morphological changes for 3 days post-inoculation. The number of dead and viable cells was counted for growth rate of the trophozoites with a haemocytometer using binocular microscope (Olympus, Japan) at 12, 24, 36, 48, 60 and 72 h of post-inoculation, respectively (Tiewcharoen et al., 2008).
Susceptibility of N. fowleri to Single Drugs
Experiments were performed in sterile 12 well plates (Corning), each well contains 1.12 cm2. Serial twofold dilutions of the single drugs, except Amphotericin B at conc. 0.976-1000 μg mL-1 were prepared in Nelson medium containing 5% FCS. Amphotericin B had ranging of concentrations at conc. 0.01-0.1 μg mL-1. Control wells received 500 μL of medium alone. Five hundred microliters of calibrated trophozoite suspension (1x104 cells mL-1) were added to each well and then the plates were sealed and incubated at 37°C. Tests were performed in duplicate and were repeated at least three times. Trophozoites were observed for cell viability at 24, 48 and 72 h of incubation under inverted microscope. At 72 h of incubation, the plates were kept at 4°C for 10 min. The cells were harvested and centrifuged at 2000 g for 2 min and then the culture medium was discarded. The cells were dissolved with 0.1 mL of PBS 7.2 containing 5% FBS. To assess the inhibitory effect of the drugs, 0.4% trypan blue in PBS was used. Dead trophozoites were stained after 5-15 min, while viable trophozoites remained unstained. The numbers of dead and viable cells were counted as described above.
Susceptibility of N. fowleri to Combined Drugs
Serial twofold dilutions of combined drugs were prepared from the various concentrations of the single drugs mixed with Amphotericin B at IC50 scores in 500 μL of Nelson medium containing 5% FCS. Control well was medium alone. The 500 μL of trophozoite suspension ((1x104 cells mL-1) was added to each well and the plates were incubated at 37°C. Total cell viability were counted as described above.
Statistics
Data were analyzed or plotted and statistics were calculated with Microsoft Excel. The results were expressed as the means±standard errors of the means (SEM). Student’s t test was used to determine the significance, which was taken as a value of p≤0.05.
RESULTS
Morphology and Growth Kinetic of N. fowleri Trophozoites
Morphology, N. fowleri trophozoites growing in axenix medium exhibited eruptive movement by producing hemispherical bulges of hyaline cytoplasm (Fig. 1a). A single nucleus with a large, centrally placed nucleolus was present in the trophozoites. The trophozoite measured from 8 to 20 μm with a mean of 10 μm based on the measurements of at least 20 trophozoites. Growth kinetic, N. fowleri trophozoites (5x103 cells at started point) were detected at about 6 to 72 h interval with doubling times. The yields of trophozoites were detectable approximately 1x107 cells after 72 h post-inoculation (Fig. 1b).
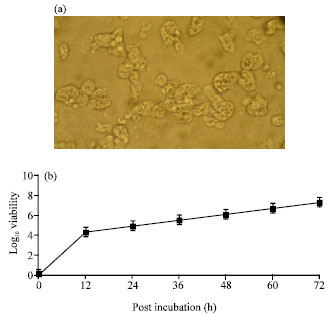 | |
| Fig. 1: | (a) Micorscopic characteristic of N. fowleri and (b) growth kinetic of N. fowleri. N. fowleri trophozoites was cultured in Nelson’s medium with 5% FCS at 37°C for 72 h. Morphological shapes was observed under inverted microscope, 400X and the viability of the cells were counted at indicated times using Trypan blue exclusion method. Results were the average of three experiments and errors were standard deviation |
Susceptibility of N. fowleri Trophozoite to Drugs Alone and Drugs Combination
Naegleria fowleri trophozoites were cultured in 12 well-plates in the presence of the various concentrations of either drugs alone or drug combination at 37°C for 72 h. At 72 h-post incubation, the IC50 and MIC100 scores for tested drugs against N. fowleri trophozoites were expressed in μg mL-1 appear in Table 1.
Antifungal Drugs
Single drugs, amphotericin B had the best IC50 and MIC100 scores at 0.0246±0.004 μg mL-1 and 0.098±0.0037 μg mL-1 (Fig. 2a). At MIC100 scores of drug combination, treatment with fluconazole plus Amphotericin B (MIC100 scores at 126.114±0.63 μg mL-1) to N. fowleri trophozoites showed 3.99 fold-lower than treated the trophozoites with fluconazol alone (MIC100 scores at 503.75±0.35 μg mL-1). The sensitivity of voriconazole plus Amphotericin B was effective to the trophozoites with MIC 100 scores at 126.072±0.62 μg mL-1, while voriconazole alone (1004.19±0.45 μg mL-1) had no detectable effect on the trophozoites (Fig. 2b).
Antibacterial Drugs
Single drugs, azithromycin, gentamicin sulphate and ciprofloxacin were effective to the N. fowleri trophozoites with the IC 50 scores at 6.767±0.04, 15.662±0.03 and 63.521±0.71 μg mL-1, respectively. Azithromycin was effective to the trophozoites MIC100 at 510.308±0.62 μg mL-1, while gentamicin sulphate or ciprofloxacin was ineffective to the trophozoites at concentrations of MIC100 scores (Table 1 and Fig. 2c). Combined treatment with azithromycin plus amphotericin B (131.206±0.64 μg mL-1) to the trophozoites was 3.89-fold less than treatment with azithromycin alone (510.308±0.62 μg mL-1), suggesting that both amphotericin B and azithromycin had the synergistic effect to N. fowleri trophzoites. In contrast, we observed the sensitivity of combined gentamicin sulphate was not different from gentamicin sulphate alone at MIC100 scores to the trophozoites. Neither amphotericin B-combined ciprofloxacin nor ciprofloxacin had detectable effect to the trophozoites. It was suggested that either ciprofloxacin or gentamicin sulphate had not synergistic effect to N. fowleri trophozoites.
| Table 1: | Mean (±SD) μg mL-1 of IC50 and MIC100 scores of either single drugs alone or single drugs plus with Amphotericin B against Naegleria fowleri Siriraj strain |
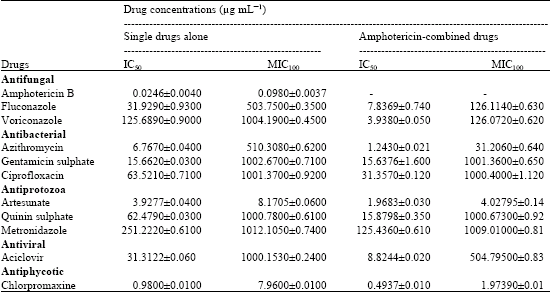 | |
Antiprotozoal Drugs
As shown in Table 1 and Fig. 2d, artesunate was effective to the N. fowleri trophozoites than quinine sulphate and metronidazol at IC50 and MIC100 scores, respectively. The sensitivities of combined artesunate with amphotericin B to the trophozoites were 2-fold less than artesunate alone at IC50 and MIC100 scores. In contrast, no susceptible activity of trophozoites to quinine sulphate alone or combined quinine sulphate was observed. Metronidazone alone or drug combination with amphotericin B had no also detectable effect on to the trophozoites. It was suggested that artesunate and amphotericin B had the synergistic effects on N. fowleri trophozoites.
Antiviral Drugs
As shown in Table 1 and Fig. 2e, the IC50 of combined acyclovir (8.8244±0.02 μg mL-1) to the N. fowleri trophozoites was 3.548 fold than acyclovir alone (31.3122±0.06 μg mL-1). The MIC100 of combined acyclovir to the N. fowleri trophozoites was concentrations of 504.795±0.83 μg mL-1, while acyclovir alone was ineffective to the trophozoites. It was suggested that combination treatment with acyclovir and amphotericin B had the synergistic effect on N. fowleri trophozoites. This is the first data showed that acyclovir in combination with amphotericin B could be used for N. fowleri infection.
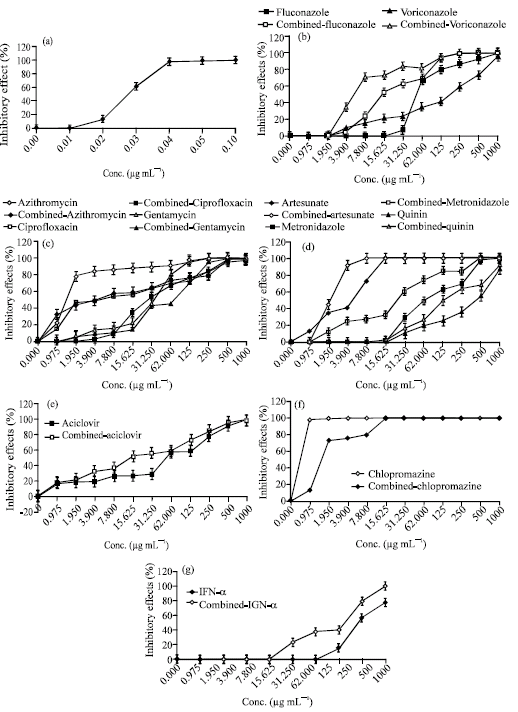 | |
| Fig. 2: | (a) Susceptibility of N. fowleri to Amphotericin B alone, (b) antifungal drugs alone or plus with Amphotericin B, (c) antibacterial drugs alone or plus with Amphotericin B, (d) antiprotozoal drugs alone or plus with Amphotericin B, (e) antiviral drugs alone or plus with Amphotericin B, (f) antiphychotic drugs alone or plus with Amphotericin B and (g) IFN-α alone or plus with Amphotericin B. N. fowleri trophozoites were studied in the presence or various concentrations of either single drug alone or single drug plus Amphotericin B at 37°C for 72 h. The number of viable trophozoites was counted by Trypan blue exclusion method. Results were the average of three experiments and errors were standard deviation |
Antiphycotic Drugs
As shown in Table 1 and Fig. 2f, chlorpromazine had an IC50 score at 0.980±0.01 μg mL-1 with a MIC100 of 7.96±0.01 μg mL-1, thereby combined Chlorpromazine had an IC50 score at 0.4937±0.01 μg mL-1 with a MIC100 of 1.9739±0.01 μg mL-1. The MIC100 scores of combined chlorpromaxine to the trophozoites were 4 fold lower than chlorpromazine alone.
Interferon-Alpha
IFN-α had an IC50 score at 5.8 log 10 IU with a MIC100 of >7 log 10 IU to N. fowleri trophozoites (data not shown). The trophozoites were susceptible to IFN-α in combination with amphotericin B (MIC100 of 6.18 log 10 IU mL-1), while they was not susceptible to IFN-α alone (Fig. 2g). It was suggested that IFN-α in combination with amphotericin B had the synergistic effect on the trophozoites.
DISCUSSION
In the present study, we examined the morphology and growth kinetic of trophozoites of N. fowleri Siriraj-strain grew in Nelson’s medium at 37°C. Morphologically, the trophozoites were similar to those of N. fowleri and other Naegleria sp., with respect to the nucleus and cytoplasmic organelles (Visvesvara et al., 2005). Growth kinetic, total double numbers of N. fowleri Siriraj strain trophozoites were also similar to those of other N. fowleri trophozoites at approximate 6 h-post inoculation, as previous described (Ondarza and Tamayo, 2004). Inhibition of growth kinetic of N. fowleri amoeba using single drugs or drug combination was the one of the choice for PAM therapy in human.
Many investigators screened a lot of drugs for in vitro and in vivo therapeutic activity against N. fowleri. Amphotericin B was used as the first therapeutic drug for N. fowleri infection (Goodman and Koenig, 1970). Other drugs such as chlorpromazine, clotrimazole, fluconazole, itraconazole, ketoconazole, metronidazole, micronazole, rifampicin and trifluoperazine had also been tested in vitro (Tiewcharoen et al., 2002; Schuster and Mandel, 1984; Vargas-Zepeda et al., 2005; Ondarza et al., 2006, 2007). Artemisinin and azithromycin has been described as effective treatment of experimental PAM in mice (Goswick and Brenner, 2003). Recently, miltefosine which was developed and used as an anticancer drug and voriconazole, which was used in systemic fungal infections were found to be effective in in vitro studies (Schuster and Visvesvara, 2004).
In this study, we evaluated the sensitivities of antifungal (amphotericin B, fluconazole and voriconazole), antibacterial (azitromycin, gentamicin sulphate and ciprofloxacin) antiprotozoa (artesunate) and antiviral (anticyclovir) and antipsychotic (chlopromazine) drugs as well as interferon-alpha against N. fowleri trophozoites. Additionally, we also evaluated the effects of synergistic drugs occur when the effect of two agents used in combination compared with the sum of the individual effects of each agent given alone. The greater synergy between two agents, the less of each agent is required to produce a specific effect. Synergy may enable the use of lower doses to produce a defined level of efficacy, thereby increasing safety and tolerability. Synergy may also enable two agents, both of which are <50% efficacious, to be combined to produce 100% efficacy. Furthermore, we were also interested the sensitivity of single drugs or drugs combination to N. fowleri during 1-3 day interval in order to follow drug-resistant N. fowleri trophozoites.
Antifungal agents, the MIC scores of amphotericin B (0.098±0.0037) had still been the highest potential against N. fowleri. Schuster and Rechthand (1975) reported that the action of amphotericin B binded to ergosterol in the fungal cell membrane immediately, resulting in depolarization of the membrane and formation of pores that leads to K+ leakage and fungal cell death. The mechanism of amphotericin B against N. fowleri was established. This drug was bind to membrane sterols, leading to membrane permeability, leakage and strongly inhibited three NADPH-dependent flavoprotein disulfide reductases CysR (NADH-dependent reductase), GR and TR, either by blocking the electron transfer from NADPH to FAD, or by reacting directly with the reducing catalytic site of N. fowleri (Ondarza et al., 2007). The MIC100 scores of fluconazole alone and combined fluconazole were 503.75±0.35 and 126.114±0.63 μg mL-1, respectively. It was suggested that the sensitivity of combined fluconazole was 3.99 times less than fluconazole against N. fowleri trophozoites. No mechanism of fluconazole against N. fowleri. trophozoites has been reported before. Earlier studies showed that fluconazole has been used for treatment of cryptococcal meningitis, dissemininated candidiasis and invasive candidial infection (Clark et al., 1996). It has been proven to penetrate the blood-brain barrier remarkably well (Yang et al., 1996). However, this drug has also shown an unexpected effectiveness for reducing organ dysfunction and mortality in septic shock, probably by increasing bactericidal activity and the recruitment of neutrophils (Jacobs et al., 2003). In this study, the amphotericin B combined fluconazole were more effective than fluconazole alone against N. fowleri. These findings are identical to previous in vitro studies reported that the amphotericin B and fluconazole had amoebocidal effect against N. fowleri. trophozoites (Tiewcharoen et al., 2002). This results were supported by the successful treatment of N. fowleri infection by using intravenous amphotericin B, fluconazole and oral rifampin in a 10 year-old boy patient (Vargas-Zepeda et al., 2005). In the present, the mechanism of fluconazole against N. fowleri has not been revealed. Clinical studies in the treatment of fungal infection sustained by Candida sp., the effect of fluconazole acts on ergosterol biosynthesis by inhibition of cytochrome P450 14a-demethylase (P450 14DM) (Bachmann et al., 2002). The MIC100 scores of voriconazole in combination with amphotericin B to the trophozoites was 126.072±0.62 μg mL-1. Our groups did not observe detectable effect of voricnazole on the trophozoites. In contrast, Schuster et al. (2006) revealed that voriconazole had the greater potency to N. fowleri trophozoites than fluconazole. As with all azole antifungal agents, voriconazole worked principally by inhibition of cytochrome P450 14a-demethylase (P45014DM). This enzyme is in the sterol biosynthesis pathway that leads from lanosterol to ergosterol compared to fluconazole, voriconazole inhibited P45014DM to a greater extend (Sanati et al., 1997). Antibacterial agents, the MIC100 scores of azithromycin (510.308±0.62 μg mL-1) were 3.89 times greater than those of combined azithromycin (131.206±0.64 μg mL-1) against N. fowleri trophozoites. It has been reported that azithromycin, a derivative of erythromycin with improved activity against Gram-negative bacteria,inhibited bacterial protein synthesis by binding to the 50S ribosomal subunit and blocking peptide bond formation and translocation including exhibits a marginal inhibition effect in a model system derived from Escherichia coli, in which a peptide bond is formed between puromycin and AcPhe-tRNA bound at the P-site of poly(U) (Dinos et al., 2001).
The susceptibility of N. fowleri trophozoites of gentamicin sulphate alone and combined gentamicin sulphate at MIC 100 were not observed. In general, gentamicin inhibited bacterial protein synthesis mainly through binding with the 30S ribosomal subunit and acted through two different mechanisms. In one mechanism at high concentrations, it could interfere with the correct amino acid polymerization and elongation. Another mechanism predominated at low concentrations in which amino acid codons were misread by tRNA and proofreading is impaired. This leaded to incorrect amino acid sequencing and nonsense proteins (Auslander et al., 1973). Like genamicin sulphate, ciprofloxacin was ineffective to N. fowleri trophozoites at the concentrations of MIC100. Ciprofloxacin was a fluoroquinolone derivative of nalidixic acid which targets DNA gyrase of the bacterial cell (Lebel, 1988). During DNA synthesis, DNA gyrase facilitated the movement of replication by releasing the tension of the coiled DNA. However, when ciprofloxacin was used, binding occurs to form a fluoroquinolone-gyrase-DNA complex. This complex stopped the replication. DNA gyrase was also involved in the supercoiling of the bacterial DNA into cell, thus, ciprofloxacin will inhibit DNA gyrase from packing the DNA into bacterial cell. As supercoiling were essential for cell growth, the inhibition eventually cause the bacterial cell to lyse (Griggs et al., 2005).
Antiprotozoal drugs; artesunate alone showed high potency at IC50 (3.9277±0.04 μg mL-1) and MIC100 (8.1705±0.06 μg mL-1). Artesunate is one of semi-synthetic derivatives of artesinin isolated from decoctions of traditional Chinese medicine Artemisia annua L. (quingho, sweet wormwood) (Meshnick and Kamchonwongpaisan, 1996). During recent years artemisinin and its derivatives have emerged as the most effective and safe drugs for the treatment of severe and chloroquine-resistant malaria (Haynes, 2001) Artemisinin and its derivatives have been demonstrated to affect a number of other cellular biochemical processes, such as anti-proliferation, anti-angiogenic effects, induction of cell apoptosis and oxidative stress (Wartenberg et al., 2003; Huan-huan et al., 2004). More recently, there is increasing evidence to support the notion that these compounds may have potential role on anti-inflammation and immune regulation. For instance, artemisinin suppresses NF-κB-mediated NO synthesis in human astrocytoma T67 cells (Aldieri et al., 2003) and the inhibitory effects of dihydroarteannuin, one of artemisinin derivatives, on elevated TNF-α production may result from the blockade of NF-κB signaling pathway in peritoneal macrophages and renal tissue from lupus BXSB mice (Li et al., 2006). 3-(12-beta-artemisininoxy) phenoxyl succinic acid (SM735), a novel artesinin derivative, was also demonstrated to play a potential immunosuppressive role (Zhou et al., 2005). Furthermore, artemisinin potently suppressed release of TNF-α and IL-6 induced by CpG-containing oligodeoxy-nucleotides, lipopolysaccharide or heat-killed Escherichia coli (Wang et al., 2006). Artemisinin inhibits TNF-α induced nuclear NF-κB translocation, DNA-binding activity and gene transcriptional activity, as well as, degradation and phosphorylation of IκBα in rheumatoid synoviocytes (Xu et al., 2007). However, the inhibitory effects of artesunate against N. fowleri are unknown. Quinine alone was effective to N. fowleri at the concentrations of IC50 = 62.479±0.03 μg mL-1 and MIC100 = 1000.78±0.61 μg mL-1, respectively. Combined-quinine showed high potency at IC50 (15.8798±0.35 μg mL-1) and MIC100 (1000.673±0.92 μg mL-1) to N. fowleri trophozoites. This data revealed the sensitivity of amphotericin B-combined quinin at IC50 scores were 3.934 times to N. fowleri than Quinin alone, suggesting that both drugs were synergistic drugs to the amoebae. Research on the mechanism of quinine and how the Naegleria has acquired quinine resistance is still ongoing. Sanchez et al. (2008) investigated passive distribution, in accordance with the intracellular pH gradients and intracellular binding could account for only a small fraction of the high amount of quinine accumulated by human malaria parasite Plasmodium falciparum. The results of trans-stimulation kinetics suggested that high accumulation of quinine had been brought about by a carrier-mediated import system. This import system might be weakened in parasites with reduced quinine susceptibility. Other data show that polymorphisms within PfCRT are causatively linked with an increased verapamil-sensitive quinine efflux that, depending on the genetic background, resulted in reduced quinine accumulation. As an antiviral agent, quinin inhibits the glycosylation of the HIV envelope glycoprotein gp120 which occurs within the golgi apparatus (Lelievre et al., 2007). Metronidazole had IC50 and MIC100 effects against Naegleria trophozoites at 251.222±0.61 and 1012.105±0.74 μg mL-1. Combined metronodazole against the trophozoites expressed as IC50 and MIC100 of 125.436±0.61 and 1009.01±0.81, respectively. The inhibitory effect of combined metronidazole was less 2 folds than metronidazole alone. Metronidazole is a prodrug. It is converted in anaerobic organism by the redox enzyme pyruvate ferredoxin oxidoreductase. The nitro group of metronidazole is chemically reduced by ferredoxin (or a ferredoxin-linked metabolic process) and the products are responsible for disrupting the DNA helical structure, thus inhibiting nucleic acid synthesis. Metronidazole is selectively taken up by anaerobic bacteria (Church et al., 2006) and sensitive protozoal organisms because of the ability of these organisms to reduce metronidazole to its active form intracellularly (Wassman et al., 1999).
Antiviral agents, the sensitivity of acyclovir alone was infective to N. fowleri trophozoites at day 3 post-inoculation. Drug combination between acyclovir and amphotericin B to N. fowleri trophozoites had the MIC100 scores of 504.795±0.83 μg mL-1. The mechanism of acyclovir had been studied in HSV-infected human cell. The acyclovir molecules entered the cell and were converted to acyclovir monophosphate by the HSV enzyme Thymidine Kinase (TK). Enzymes in the human cell added two more phosphates to eventually form the active drug acyclovir triphosphate. The acyclovir triphosphate competed with 2-deoxyguanosine triphosphate (dGTP) as a substrate for viral DNA polymerase, as well as acting as a chain terminator. In actual infection, the HSV released its naked capsid that delivers DNA to the human nucleus; the active drug acyclovir triphosphate exerted its action on the viral DNA located in the nucleus (Balfour, 1999). It was proposed that both drugs had synergistic drugs against N. fowleri trophozoites, thereby acyclovir and amphotericin B inhibited chain elongation and protein synthesis of N. fowleri trophozoites, respectively.
Antipsychotic agents, the phenothiazine compounds chlorpromazine hydrochloride either alone or in combination with amphotericin B had in vitro activity against N. fowleri trophozoites. The MIC100 scores of chlorpromazine and combined chlorpromazine to N. fowleri trophozoites were 7. 96±0.01 and 1.9739±0.01 μg mL-1, respectively. The MIC100 scores of chlorpromazine were 4.03 folds rather than combined chlorpromazine. Chlorpromazine inhibited 80% cysteine reductase and glutathione reductase of the N. fowleri trophozoites and decreased 80% trophozoites during the 24 h incubation (Kim et al., 2008) where as the amphotericin B and miconazole inhibited were the most potent inhibitors of trypanothione reductase and inhibited 100% trophozoites (Ondarza et al., 2007). Moreover, chlorpromazine reflected sensitivity of amoeba calcium regulatory protein to the phenothiazine compounds or was due to the lipophilic action of the drugs on the amoeba plasma membrane (Schuster and Mandel, 1984).
Interferon, the sensitivity of IFN-α to N. fowleri trophozoites was 5.6 log10 IU mL-1 of IC50 scores and >6.18 log10 IU mL-1 of MIC100 scores compared with one of amphotericin B-combined IFN-α was 5.1 log10 IU mL-1 of IC50 and 6.18 log10 IU mL-1 of MIC100, respectively. Thyrell et al. (2002) found that apoptosis induced by IFN-α was associated with activation of caspases-1, -2, -3, -8 and -9 and that this activation was a critical event. Caspase-3 activation was dependent on activity of caspases-8 and -9, moreover, activation of caspase-8 seems to be the upstream event in IFN-α-induced caspase cascade. Thyrell et al. (2002) also found loss of mitochondrial membrane potential as well as release of cytochrome c post IFN-treatment, clearly implicating the involvement of mitochondria in IFN-mediated apoptosis. Furthermore, IFNα-induced apoptosis was found to be independent on interactions between the Fas-receptor and its ligand.
CONCLUSION
In the present study, we evaluated additional synergy studies of amphotericin Bwith other agents in order to improve drug selection and treatment of N. fowleri infection. Present results indicated that amphotericin B plus chlorpromazine were new synergistic drugs against N. fowleri trophozoites. Moreover, we first reported that acyclovir or IFN-α in combination with amphotericin B had effect on N. fowleri trophozoites. It was suggested that the combined use of these agents might provide a useful regimen for treating human infections with this organism. In Further interesting topic, the drug mechanisms and the cytopathic effect in mouse model against N. fowleri should be studied.
ACKNOWLEDGMENTS
This study was partial supported by the Faculty of Medicine, Siriraj Hospital, Mahidol University Foundation in 2007-2009 with Grant Number 017(II)50 and partial supported by with Grant Number RGP 2551/01 from Department of Biology, Faculty of Science, Silpakorn University at Sanamchan Palace, Nakhon Pathom, Thailand.
REFERENCES
- Aldieri, E., D. Atragene, L. Bergandi, C. Riganti, C. Costamagna, A. Bosia and D. Ghigo, 2003. Artemisinin inhibits inducible nitric oxide synthase and nuclear factor NF-B activation. FEBS. Lett., 552: 141-144.
PubMedDirect Link - Anand, B.S. and A.K. Mitra, 2009. Mechanism of corneal permeation of L-Valyl Ester of acyclovir: Targeting the oligopeptide transporter on the rabbit cornea. Pharm Res., 19: 1194-1202.
CrossRefDirect Link - Auslander, D.E., A. Felmeister, B.J. Sciarrone and A.B. Segelman, 1973. Proposed mechanism for gentamicin transport. J. Pharm. Sci., 62: 167-168.
PubMedDirect Link - Bachmann, S.P., T.F. Patterson and T.F. Lopez-Ribot, 2002. In vitro activity of caspofungin MK-0991) against Candida albicans clinical isolates displaying different mechanisms of azole resistance. J. Clin. Microbiol., 40: 2228-2230.
CrossRefDirect Link - Cursons, R., J. Sleigh, D. Hood and D. Pullon, 2003. A case of primary amoebic meningoencephalitis: North Island, New Zealand. N. Z. Med. J., 116: 712-712.
PubMedDirect Link - Dinos, G.P., M. Michelinaki and D.L. Kalpaxis, 2001. Insights into the mechanism of azithromycin interaction with an Escherichia coli functional ribosomal complex. Mol. Pharm., 59: 1441-1445.
Direct Link - Goodman, J.S. and M.G. Koenig, 1970. Amphotericin B-specifics of administration. Mod. Treat., 7: 581-595.
PubMedDirect Link - Goswick, S.M. and G.M. Brenner, 2003. Activities of azithromycin and amphotericin B against Naegleria fowleri in vitro and in a mouse model of primary amebic meningoencephalitis. Antimicrob. Agents Chemother., 17: 524-528.
CrossRefDirect Link - Griggs, D.J., M.M. Johnson, J.A. Frost, T. Humphrey, F. Jorgensen and L.J.V. Piddock, 2005. Incidence and mechanism of ciprofloxacin resistance in Campylobacter spp. Isolated from commercial poultry flocks in the United Kingdom before, during and after fluoroquinolone treatment. Antimicrob. Agents Chemother., 49: 699-707.
CrossRefDirect Link - Li, W.D., Y.J. Dong, Y.Y. Tu and Z.B. Lin, 2006. Dihydroarteannuin ameliorates lupus symptom of BXSB mice by inhibiting production of TNF- and blocking the signaling pathway NF-kappa B translocation. Int. Immunopharmacol., 6: 1243-1250.
Direct Link - Haynes, R.K., 2001. Artemisinin and derivatives: The future for malaria treatment? Curr. Opin. Infect. Dis., 14: 719-726.
PubMedDirect Link - Huan-Huan, C., Y. Li-Li and L. Shang-Bin, 2004. Artesunate reduces chicken chorioallantoic membrane neovascularisation and exhibits antiangiogenic and apoptotic activity on human microvascular dermal endothelial cell. Cancer Lett., 211: 163-173.
Direct Link - Jacobs, S., E.D.A. Price, M. Tariq and N.F. Al-Omar, 2003. Fluconazole improves survival in septic shock: A randomized double-blind prospective study. Crit. Care Med., 31: 1938-1946.
PubMedDirect Link - John, D.T., 1982. Primary amebic meningoencephalitis and the biology of Naegleria fowleri. Ann. Rev. Microbiol., 36: 101-123.
CrossRefDirect Link - Kim, J.H., S.Y. Jung, Y.J. Lee, K.J. Song and D. Kwon(et al)., 2008. Effect of therapeutic chemical agents in vitro and on experimental meningo-encephalitis due to Naegleria fowleri. Antimicrob. Agents Chemother., 52: 4010-4016.
Direct Link - Lebel, M., 1988. Ciprofloxacin: Chemistry, mechanism of action, resistance, antimicrobial spectrum, pharmacokinetics, clinical trials and adverse reactions. Pharmacotherapy, 8: 3-33.
PubMedDirect Link - Lelievre, J., A. Berry and F. Benoit-vical, 2007. Artemisinin and chloroquine: Do mode of action and mechanism of resistance involve the same protagonists? Curr. Opin. Investig. Drugs, 8: 117-124.
PubMedDirect Link - Liu, Y.J., 2005. IPC: Professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors. Annu. Rev. Immunol., 23: 275-306.
CrossRefPubMedDirect Link - Ma, P., G.S. Visvesvara, A.J. Martinez, F.H. Theordore, P.M. Daggett and T.K. Sawyer, 1990. Naegleria and Acanthamoeba infections. Rev. Infect. Dis., 12: 490-513.
Direct Link - McCurdy, D.K., M. Frederic and J.R. Elkinton, 1968. Renal tubular acidosis due to amphotericin B. N. Engl. J. Med., 278: 124-130.
PubMedDirect Link - Kamchonwongpaisan, S. and S.R. Meshnick, 1996. The mode of action of the antimalarial artemisinin and its derivatives. General Pharmacol., 27: 587-592.
CrossRef - Tamayo, E.M. and R.N. Ondarza, 2004. Characterization of two long vesicle-associated membrane proteins or longins genes from Entamoeba histolytica. Arch. Med. Res., 35: 378-384.
CrossRefDirect Link - Ondarza, R.N., A. Iturbe and E. Hernandez, 2006. In vitro antiproliferative effects of neuroleptics, antimycotics and antibiotics on the human pathogens Acanthamoeba polyphaga and Naegleria fowleri. Arch. Med. Res., 37: 723-729.
Direct Link - Ondarza, R.N., A. Iturbe and E. Hernandez, 2007. The effects by neuroleptics, antimycotics and antibiotics on disulfide reducing enzymes from the human pathogens Acanthamoeba polyphaga and Naegleria fowleri. Exp. Parasitol., 115: 41-47.
CrossRef - Proffitt, R.T., A. Satorious, S.M. Chiang, L. Sullivan and J.P. Adler-Moore, 1991. Pharmacology and toxicology of a liposomal formulation of amphotericin B (AmBisome) in rodents. J. Antimicrob. Chemother., 28: 49-61.
Direct Link - Sanchez, C.P., W.D. Stein and M. Lanzer, 2008. Dissecting the components of quinine accumulation in Plasmodium falciparum. Mol. Microbiol., 67: 1081-1093.
PubMed - Schuster, F.L. and E. Rechthand, 1975. In vitro effects of amphotericin B on growth and ultrastructure of the amoeboflagellates Naegleria gruberi and Naegleria fowleri. Antimicrob. Agents Chemother., 8: 591-605.
PubMedDirect Link - Schuster, F.L. and N. Mandel, 1984. Phenothiazine compounds inhibit in vitro growth of pathogenic free-living amoebae. Antimicrob. Agents Chemother., 25: 109-112.
Direct Link - Schuster, F.L. and G.S. Visvesvara, 2004. Opportunistic amoebae:Challenges in prophylaxis and treatment. Drug Resist. Update, 7: 41-51.
PubMedDirect Link - Schuster, F., J.B. Gudlielmo and G.S. Visvesvara, 2006. In vitro Activity of Miltefosine and Voriconazole on Clinical Isolates of Free-Living Amebas: Balamuthia mandrillaris, Acanthamoeba spp. and Naegleria fowleri. J. Eukar. Microbiol., 53: 121-126.
Direct Link - Shin, N.J. and K.I. Im, 2004. Pathogenic free-living amoebae in Korea. Korean J. Parasitol., 42: 93-119.
Direct Link - Soltow, S.M. and G.M. Brenner, 2007. Synergistic activities of azithromycin and amphotericin B against Naegleria fowleri, in vitro and in a mouse model of primary amebic meningoencephalitis. Antimicrob. Agents Chemother., 51: 23-27.
CrossRefDirect Link - Taylor, J.L., P. Tom and O.W.J. Brien, 1998. Combined effects of interferon-α and acyclovir on herpes simplex virus type 1 DNA polymerase and alkaline DNase. Antiviral Res., 38: 95-106.
Direct Link - Tiewcharoen, S., V. Junnu and P. Chinabut, 2002. In vitro effect of antifungal drugs on pathogenic Naegleria spp. Southeast Asian J. Trop. Med. Public Health, 33: 38-41.
PubMedDirect Link - Tiewcharoen, S., N. Malainual, V. Junnu, P. Chetanachan and J. Rabablert, 2008. Cytopathogenesis of Naegleria fowleri Thai strains for cultured human neuroblastoma cells. Parasitol. Res., 102: 997-1000.
CrossRefDirect Link - Thyrell, L., S. Erickson, B. Zhivotovsky, K. Pokrovskaja and O. Sangfelt et al., 2002. Mechanisms of Interferon-alpha induced apoptosis in malignant cells. Oncogene., 21: 1251-1262.
PubMedDirect Link - Vargas-Zepeda, J., A.V. Gomez-Alcala, J.A. Vazquez-Morales, L. Licea-Amaya, J.F. de Jonckheere and F. Lares-Villa, 2005. Successful treatment of Naegleria fowleri meningoencephalitis by using intravenous amphotericin B, fluconazole, and rifampicin. Arch. Med. Res., 36: 83-86.
CrossRefDirect Link - Visvesvara, G.S., J.F. de Jonckheere, R. Sriram and B. Sriram, 2005. Isolation and molecular typing of Naegleria fowleri from the brain of a cow that died of primary amebic meningoencephalitis. J. Clin. Microbiol., 43: 4203-4204.
Direct Link - Wang, J., H. Zhou, J. Zheng, J. Cheng and W. Liu(et al)., 2006. The antimalarial artemisinin synergizes with antibiotics to protect against Lethal live Escherichia challenge by decreasing proinflammatory cytokine release. Antimicrob. Agents Chemother., 50: 2420-2427.
CrossRefPubMedDirect Link - Wartenberg, M., S. Wolf, P. Budde, F. Grunheck and H. Acker(et al)., 2003. The antimalaria agent artemisinin exerts antiangiogenic effects in mouse embryonic stem cell-derived embryoid bodies. Lab. Invest., 83: 1647-1655.
Direct Link - Wassmann, C., A. Hellberg, E. Tannich and I. Bruchhaus, 1999. Metronidazole resistance in the protozoan parasite Entamoeba histolytica is associated with increased expression of iron-containing superoxide dismutase and peroxiredoxin and decreased expression of ferredoxin 1 and flavin reductase. J. Biol. Chem., 274: 26051-26056.
CrossRefDirect Link - Yang, H., Q. Wang and W.F. Elmquist, 1996. Fluconazole distribution to the brain: A crossover study in freely-moving rats using in vivo microdialysis. Pharm. Res., 13: 1570-1575.
Direct Link - Zhou, W.L., J.M. Wu, Q.L. Wu, J.X. Wang and Y. Zhou(et al)., 2005. A novel artemisinin derivative, 3-(12-beta-artemisininoxy) phenoxyl succinic acid (SM735), mediates immunosuppressive effects in vitro and in vivo. Acta Pharmacol. Sin., 26: 1352-1358.
PubMedDirect Link