
D. Kumar
Department of Biotechnology, Abhilashi Institute of Life-Sciences, Tanda, Ner Chowk, Mandi-175008, H.P., India
Savitri
Department of Biotechnology, Abhilashi Institute of Life-Sciences, Tanda, Ner Chowk, Mandi-175008, H.P., India
N. Thakur
Department of Biotechnology, Abhilashi Institute of Life-Sciences, Tanda, Ner Chowk, Mandi-175008, H.P., India
R. Verma
Department of Biotechnology, Abhilashi Institute of Life-Sciences, Tanda, Ner Chowk, Mandi-175008, H.P., India
T. C. Bhalla
Department of Biotechnology, Himachal Pradesh University, Shimla-171005, H.P., India
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 12 | Page No.: 661-672







ABSTRACT
Proteases represents one of the major groups of industrial enzymes and a number of detergent stable proteases have been isolated and characterized because of its widespread use in detergents. It is worthwhile to screen microbes from new habitats for proteases with novel properties to meet the needs of rapidly growing detergent industry. High-alkaline serine proteases have been successfully applied as protein degrading components of detergent formulations and are subject to extensive protein engineering efforts to improve their stability and performance. Protein engineering has been extremely used to study the structure-function relationship in proteases and led to deeper understanding of the factors influencing the cleaning performance of detergent proteases. This study, discusses the types and sources of proteases with an overview on applications of proteases as laundry detergent additives and some advances in improving the stability and performance of detergent enzymes.
PDF Abstract XML References Citation
How to cite this article
D. Kumar, Savitri, N. Thakur, R. Verma and T. C. Bhalla, 2008. Microbial Proteases and Application as Laundry Detergent Additive. Research Journal of Microbiology, 3: 661-672.
URL: https://scialert.net/abstract/?doi=jm.2008.661.672
URL: https://scialert.net/abstract/?doi=jm.2008.661.672
INTRODUCTION
The estimated value of world wide use of industrial enzyme has increased from $1 billion in 1995 to $1.5 billion in 2000 (Kirk et al., 2002). As per the forecast, the global demand for enzymes will rise 7% per annum through 2006 to $6 billion in 2011 (McCoy, 2000). Proteases represent one of the major groups of industrial enzymes, because of their widespread use in detergents and dairy industry and industrial sales of protease are estimated at more than $350 million annually (Kirk et al., 2002). Proteases account for the 60-65% of the global industrial enzyme market and out of this 25% is constituted by alkaline proteases, 3% by trypsin, 10% by renin and 21% by the other proteases (Bhosale et al., 1995; Rao et al., 1998). Proteases show a vast diversity in their physio-chemical and catalytic properties and lot of literature is available on their biochemical and biotechnological aspects (Rao et al., 1998; Saeki et al., 2007). The proteases of industrial importance are obtained from animals, plants and microorganisms. The proteolytic enzymes hydrolyse the peptide links of proteins and peptides to form smaller subunits of amino acids and are produced both extracellularly as well as intracellularly (Gajju et al., 1996; Kumar et al., 2002). The proteases play an important role in a wide range of industrial processes viz., baking, brewing, detergents, leather processing, pharmaceuticals, meat tenderization, cosmetics, peptide synthesis and medical diagnosis (Bhalla et al., 1999; Kumar and Bhalla, 2005; Kumar et al., 2003; Gupta et al., 2002; Najafi et al., 2005). The application of proteases in different industries is show in Table 1. After the food industry, proteases find its widespread application in laundry detergents (Maurer, 2004; Prakash et al., 2005; Jaswal and Kocher, 2007; Takimura et al., 2007).
| Table 1: | Application of proteases in industry |
 | |
The thermostability and activity at high pH are the characteristics that have made proteolytic enzymes an ideal candidate for laundry applications (Gupta et al., 2002). Proteases are present in different brands of detergents for use in home and commercial establishments. It is important to understand the type and characteristics of protease before designing a new detergent enzyme for laundry formulation. This study gives an overview on properties and applications of proteases for laundry detergent formulations and some advances in improving the stability and performance of detergent enzymes.
TYPES OF MICROBIAL PROTEASE SYSTEMS
Microbial proteases are classified in to various groups, depending on whether they are active under acidic, neutral, or alkaline conditions and on the characteristics of the active site group of the enzyme (Garcia-Carreno, 1991). There are four classes of proteases with six families recognized by the International Union of Biochemistry viz., serine I and II, cysteine (plants), cysteine (animals), aspartic and metalloproteases. The microbial proteases which cleave internal peptide bonds are subdivided into four groups according to their side chain specificity and on the functional group present at the active site (Rao et al., 1998):
| • | Serine proteases (EC.3.4.21) |
| • | Cysteine proteases (EC.3.4.22) |
| • | Aspartic proteases (EC.3.4.23) |
| • | Metalloproteases (EC.3.4.24) |
The schematic division of proteases is given in Fig. 1. Only serine protease finds their application in detergent formulations. On the other hand, thiol proteases (e.g., papain) easily get oxidized by the bleaching agents and metalloproteases (e.g., thermolysin) lose their metal cofactors due to complexing with the water softening agents or hydroxyl ions.
Alkaline serine proteases are the most important group of enzymes exploited commercially. They have a nucleophilic serine residue in their active site and are also distinguished by having aspartate and histidine residues which, along with serine forms the catalytic tried (Maurer, 2004). They are generally active from neutral to alkaline pH, with optimum at pH 7-11. However, high optima with some protease have been reported and show broad substrate specificity and are generally of low molecular mass (18-35 kDA). Various types of serine proteases have been described in great detail earlier by Kalisz (1988), Garcia-Carreno (1991) and Morihara (1974). A number of thermophilic alkaline serine proteases have been reported from Thermus sp. Strain TOK3 (Saravani et al., 1989), Bacillus stearothermophilus AP-4 (Dhandapani and Vijayaragavan, 1994), Bacillus sp. KSM-K16 (Kobayashi et al., 1995), Thermoactinomycetes sp. E79 (Lee et al., 1996), Bacillus cereus (Prakash et al., 2005) and Bacillus circulans (Jaswal and Kocher, 2007) and used in detergents.
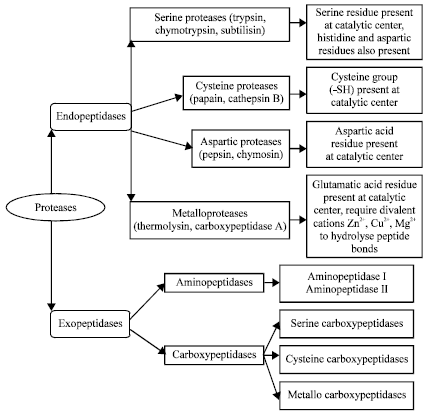 | |
| Fig. 1: | Classification of proteases |
Â
Subtilisins
Subtilisins are a prototypical group of bacterial serine proteases used extensively in detergents. Subtilisins are defined by their catalytic mechanism as serine proteases. Their amino acid sequence and three dimensional structures can be clearly differentiated from other serine proteases, such as chymotrypsin, carboxypeptidase and peptidase A from E. coli. All the subtilisins used in detergents have a size of approximately 27 kDA. Alkaline proteases from Bacillus amyloliquefaciens and Bacillus licheniformis represents the lead molecules for subtilisin. Subtilisins are used in all types of laundering detergents and used to degrade proteinaceous stains (Ward, 1986). Three subtilisins of detergent importance are (i) Subtilisin Carlsberg produced by B. licheniformis (Barfoed, 1981; Geesecke et al., 1991) and was discovered in 1947 by Linderstrom, Lang and Otteson at Carlsberg Laboratory (Rao et al., 1998); (ii) Subtilisin bacterial protease nagase (BPN) from Bacillus subtilis, B. stearothermophilus and B. amyloliquefaciens and (iii) Subtilisin Novo produced from B. subtilis.
Other Alkaline Proteases
In addition to the serine proteases a large number of uncharacterized proteases have been produced by alkaliphic bacteria with a potential for its application in detergent formulations. Some of these includes viz, Bacillus spharicus (Singh et al., 2004), Bacillus sp. (Patel et al., 2005), alkaliphilic bacteria (Dodia et al., 2006), Bacillus brevis (Aftab et al., 2006), Bacillus halodurans (Ibrahahim et al., 2007), Bacillus licheniformis N-2 (Nadeem et al., 2007) and Bacillus sp. (Chu, 2007).
PROTEASES AS LAUNDRY DETERGENT ADDITIVE
Enzymes have been added to laundry detergents for over 50 years to facilitate the release of proteinaceous materials in stains such as those of milk and blood. The proteinaceous dirt coagulates on the fabric in the absence of proteinases as a result of washing conditions (Maurer, 2004). The enzyme remove not only the obvious stains, such as blood, but also other obvious materials including proteins from body secretion and food such as milk, egg, meat and fish. The suitability of an enzyme preparation for use in detergents depends on its compatibility with the detergents at a high temperature. An ideal detergent enzyme should be stable and active in the detergent solution and should have adequate temperature stability to be effective in a wide range of washing temperatures (Bhosale et al., 1995).
The proteases are one of the standard ingredients of all kinds of detergents ranging from those used for household laundering to reagents used for cleaning contact lenses or dentures. The preparation of the first enzymatic detergent Brunus dates back to 1913 and consisted of sodium carbonate and a crude pancreatic extract. The first detergent containing the bacterial enzyme was introduced in 1956 under the trade name Bio-40. In 1960, Danish company Novo Industry A/S introduced Alcalase, produced by Bacillus licheniformis under commercial name Biotex. This was followed by Maxatase, detergent by Gist-Brocades (Rao et al., 1998). Detergents such as Era Plus ® (Procter and Gamble), Tide ® (Colgate Palmolive) and Dynamo ® (Procter and Gamble) contain proteolytic enzymes, the majority of which are produced by the members of the genus Bacillus (Samal et al., 1989). During 1995, high cost of manufacturing and increasing pressure from manufacturers to reduce the raw material cost led to rationalization of detergent enzyme industries. Following this, Genencor International purchased the detergent enzyme business of Gist-Brocades and Solvay and Novo Nordisk acquired Showa-Denko detergent enzyme business. Today, Novozymes and Genencor International are the major suppliers of detergent enzymes, supplying up to 95% of the global market of proteases. All major subtilisins for detergents are produced by Bacillus, because these species are able to secrete large amounts of extracellular enzymes (Gupta et al., 2002).
ENZYME DETERGENT GRANULATION AND FORMULATIONS
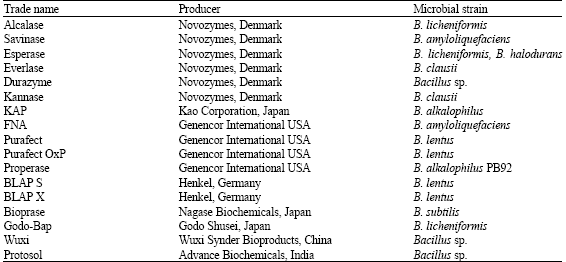
The use of enzymes in detergent formulations is now common in developed countries, with over half of all detergents presently available containing enzymes. However, in today’s dynamic market, the most successful detergent brands combine proteases with other enzymes to deliver outstanding cleaning performance. In spite of the fact, that the detergent industry is one of the largest market for enzymes, details of the enzymes used and the ways in which they are used, have rarely been published by Graham et al. (2001). Other enzymes used in detergents include α-amylases, cellulases and lipases (Gerhartz, 1990). A number of subtilisin variants are used in detergent worldwide with different trade names (Table 2).
The rapid growth of detergents was temporarily set back in the early 1970s, when workers in detergent factories developed allergies to the enzyme preparations. Enzyme manufacturers solved this problem by developing dust free protease formulations. Today, the enzyme preparations are marketed either as a stabilized enzyme solution or as encapsulated and coated granulates. The liquid preparations normally have a reduced water content and contain significant amounts of 1,2-propanediol. Granulation processes makes the use of extrusion, high shear mixing and fluidized beds (Becker et al., 1997). In developing dust-free granulates (about 0.5 mm in diameter), the enzyme is incorporated into an inner core, containing inorganic salts (e.g., NaCl) and sugars as preservative, bound with reinforcing, fibers of carboxymethylcellulose or similar protective colloid. This core is coated with inert waxy materials made from paraffin oil or polyethylene glycol plus various hydrophilic binders, which later disperse in the wash.
| Table 2: | Some commercial protease detergents |
 | |
| Table 3: | Composition of a powder enzyme detergent and liquid detergent |
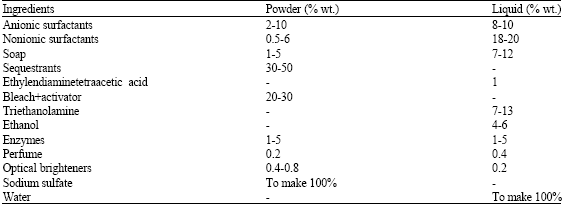 | |
This combination of materials both prevents dust formation and protects the enzymes against damage by other detergent components during the storage (Chaplin and Bucke, 1990). After release from granulated form enzyme must withstand anionic and non-ionic detergents, soaps, oxidants such as sodium perborate which generate hydrogen peroxide, optical brighteners and various less-reactive materials, all at pH values between 8.0 and 10.5 (Table 3).
DETERGENT GELS
Laundering is a worldwide common need, which has to be satisfied form view point of health, hygiene-care and life style. Cleaning habits are changing day by day with the development of new technologies, new soil substrate systems and diversity of water. The rapid growth and diversity of detergent market requires continuous consideration of new materials to meet the demand of new, improved and modified products. Detergent gels are exciting new type of detergent formulations used for fabric cleaning with advantages viz, 100% water solubility, preventing fibers weakening and depolarization, eco-friendly, reduced environmental impacts, easy and small dosing with high cleaning performance (Rachna and Tyagi, 2006). A typical detergent gel consists of all or some of the components shown in Table 4 in addition to the special ingredients designed for specific application.
| Table 4: | Common constituents of detergent gels |
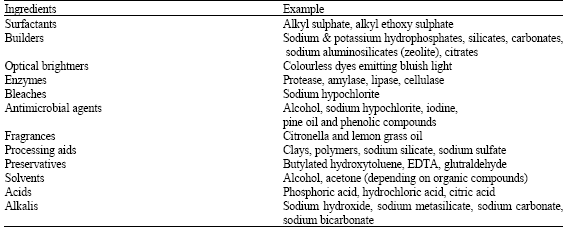 | |
| Reference (Rachna and Tyagi, 2006) | |
SEARCH FOR NOVEL DETERGENT PROTEASES
Until a few years ago new enzymes for the detergent industry were isolated in classical way by screening microorganisms in nature. Industrial strain improvement programs using classical microbiological methods have been carried out over many years and have resulted in the development of several highly productive strains (Maurer, 2004). This practice is still used today for screening but new technologies viz, genetic engineering and protein engineering are changing the way new enzymes are developed. Genetic engineering can help to increase production yield and thereby open up new possibilities. Enzymes from genetically engineered microorganisms are already in the market and the number will increase significantly in the future and this development will give both more stable and better performing enzymes in the future. Protein-engineered enzymes entered the market at the beginning of the 1990s and established themselves as benchmarks in several applications (Maurer, 2004). Kobayash et al. (1995) have isolated an alkalophilic Bacillus sp. KSM-K16 that produces an alkaline protease with properties that fulfill the essential requirements for use in both powder and liquid detergents. The structural gene for a thermostable protease from B. stearothermophilus was cloned in a plasmid pTB90 and was expressed in both B. stearothermophilus and B. subtilis. Recombinat plasmid produced 15 fold increase in protease production compared to the wild type strain (Fujii et al., 1983).
All detergent proteases currently used in the market are serine proteases produced by Bacillus strains. B. licheniformis is one of the predominant sources of alkaline proteases (Gupta et al., 2002). The fungal alkaline proteases are advantageous due to the ease of downstream processing to prepare a microbe free enzyme. An alkaline protease from Conidiobolus coronatus was found to be compatible with the commercial detergents used in India (Phadatare et al., 1993) and retained 43% of its activity at 50°C for 56 min in the presence of Ca2+ (25 mM) and glycine (1M) (Bhosale et al., 1995). Other detergent proteases have been isolated from Conidiolobus coronatus (Bhosale et al., 1995; Phadatare et al., 1993), B. licheniformis (Ward, 1983), B. stearothermophilus (Dhandapani and Vijayaragavan, 1994; Cowan et al., 1985; Coolbear et al., 1991), B. thermoruber (Tomaschova et al., 1998), Thermoactinomyces sp. HS682 (Tsuchiya et al., 1992) and Bacillus sp. APR-4 (Kumar and Bhalla, 2004a, b).
New interest in properties such as low-temperature performance and the complexity of the patent situation has led to renewed interest in screening for novel enzymes in nature. The development of new detergent compositions will also have an important impact on the development of new proteases and other types of detergent enzymes. The search for new proteases is, of course, not limited to subtilisins, but is also directed at finding completely new protease backbones. Some interesting molecules have been identified, but none of them has as yet made it into a detergent product (Cherry and Fidantsef, 2003). Every year approximately ten new wild-type subtilisins are now being described in the scientific or patent literature. Interesting enzymes are still isolated by classical microbiological screening methods e.g., the oxidation-stable subtilisin (Saeki et al., 2000, 2002). Graham et al. (2001) designed a laundry sheet containing a predetermined amount of a laundry detergent which permits the consumer to simply add the laundry sheet to the washing machine and can optionally include a fabric softener or other chemicals such as brighteners, oxidizing agents and provide an environmentally friendly detergent system to avoid the use of powdered detergents. The stabilization of proteases in liquid preparations is still a field for research (Russell and Britton, 2002). The major problem in aqueous environments is auto-proteolysis. Some general principles in formulating liquid detergents include the reduction of the free water concentration and the use of reversible inhibitors like borate or phenyl-boronic acid derivatives. Also, the composition and nature of the surfactants in the liquid detergent greatly influence the storage stability of the enzyme.
In addition to microbiological screening methods based on the cultivation of protease-producing microorganisms, the exploitation of genome programs and metagenomic screening methods have been established and enlarging the screening pool (Gupta et al., 2002; Lorenz et al., 2001).
ENGINEERING OF DETERGENT ENZYMES FOR IMPROVED STABILITY AND CATALYTIC BEHAVIOUR
Protein engineering allows the introduction of pre-designed changes into the gene for synthesis of protein with an altered function that is desired for its application. Protein engineering techniques, where one or more amino acids in the protein are changed, enable manufacturer to improve enzyme for the detergent industry. Protein engineering creates possibilities for new as well as old enzymes. Most of the industrially important enzymes are now produced from genetically engineered microorganisms. This technique has made possible to selectively delete domains from multifunctional proteins and to examine the loss of the presumed function (Gupta et al., 2002). Site directed mutation of active site amino acids has made it feasible to test their functional importance. This method has already proved its value in the study of pancreatic trypsin and carboxypeptidase and can be extended to many other proteases of industrials importance.
Protein engineering with subtilisins began in 1984 and all the amino acid positions have been modified either by site-directed mutagenesis based on rational design or, by different methods of random mutagenesis. Replacement of Met222 (adjacent to the active Ser221) with amino acid residues that are stable towards hydrogen peroxide has become a example of a rational approach to site-directed mutagenesis (Estell et al., 1985). Dynamics play an important role in determining enzyme activity and specificity and it is therefore of interest to establish how local changes in internal mobility affect protein stability, specificity and performance. The dynamic properties of the 269 residue serine proteases subtilisin PB92, secreted by Bacillus lentus and an engineered quadruple variant, DSAI, has improved washing performance. The loop regions involved in substrate binding become more structured in the engineered variant as compared with the two native proteases, exhibiting a relation between altered dynamics and performance (Mulder et al., 1999).
Substitutions at nearly every position in the mature 275 amino acid BPN0 subtilisin (Bacillus Protease Novo type, subtilisin from B. amyloliqefaciens) had been claimed in patents. The BPN0 subtilisin is generally considered to be the lead molecule for subtilisin modifications and mutations in other subtilisins often refer to the homologous position in this lead molecule. There are some excellent general reviews on the protein engineering of subtilisin, as well as articles on more specific detergent applications (Bryan, 2000; Bott, 1997). The three-dimensional structures of engineered variants of Bacillus lentus subtilisin have been engineered to exhibit altered flexibility and increased activity of RSYSA and DSAI were determined by X-ray crystallography. In addition to identifying changes in atomic position a method for identifying protein segments with altered flexibility is reported. This method reveals changes in main-chain mobility in both variants. Residues 125-127 have increased mobility in the RSYSA variant while residues 100-104 have decreased mobility in the DSAI variant. These segments are located at the substrate-binding site and changes in their mobility relate to the observed changes in proteolytic activity and the effects of altered crystal lattice flexibility becomes apparent when these variants were compared with the native enzyme (Graycar et al., 1999).
Gene shuffling is another engineering approach performed with subtilisins. Interesting results from the shuffling of 26 protease genes have been described for properties such as activity in organic solvents, temperature stability and activity at high or  low pH (Ness et al., 1999, 2002; Wintrode et al., 2000). A highly flexible region for a psychrophilic enzyme, TA39 subtilisin (S39), was transferred in silico to the mesophilic subtilisin, savinase (EC 3.4.21.62), from Bacillus lentus (clausii). The engineered hybrid and savinase were initially investigated by molecular dynamic simulations at 300K to show binding region and global flexibility. By site-directed modifications, the region was transferred to the binding region of savinase, thus a savinase-S39 hybrid, named H5, was constructed. The designed hybrid showed the same temperature optimum and pH profile as savinase. The H5 hybrid showed increased activity at low temperature, increased binding region and global flexibility, as investigated by molecular dynamic simulations and global destabilization from differential scanning calorimetric measurements (Tindbaek et al., 2004).
Legendre et al. (2000) used subtilisin 309 to illustrate the potential of phage display method for the selection of enzymes with desired properties, modifying its substrate specificity with respect to the amino acid at the P4 site of the substrate. As all mutations affecting the specificity of subtilisins may also influence the autoproteolytic processing of the proenzyme to the mature form and the engineering of the proregion or its uncoupling from this biosynthesis step becomes relevant (Takagi and Takahashi, 2003; Almog et al., 1998). Choi et al. (1999) have reported cloning of a gene encoding a serine type protease from an extreme thermophilic Aquifex pyrophilus, using a sequence tag containing the consensus sequence of protease as a probe. Sequence analysis showed an open reading frame of 619 amino acid residues with three residues forming the catalytic site of this serine protease. Protein protease inhibitors could potentially be used to stabilize proteases in commercial liquid laundry detergents and many inhibitors are susceptible to hydrolysis inflicted by the protease. Streptomyces subtilisin inhibitor (SSI) has been engineered to resist proteolysis by adding an inter-chain disulfide bond and removing a subtilisin cleavage site at leucine 63. These stabilizing changes were combined with changes to optimize the affinity for subtilisin, the resulting inhibitor provided complete protease stability for at least 5 months at 31°C in a subtilisin-containing liquid laundry detergent (Gunz et al., 2004). The success of all above approaches has to be evaluated in market for its effective use.
CONCLUSION
The use of enzymes in detergents can be extended both geographically and numerically. They have not found widespread use in developing countries which are often hot and dusty thus making frequent washing of clothes necessary. The development of subtilisins as typical detergent proteases has employed all the tools of enzyme technology, resulting in a constant flow of new and improved enzymes. A number of protease molecules have been identified and characterized and are in clear opposition to the number of molecules that are entering the market. Liquid detergents have to be formulated around the needs of the enzymes they contain, optimizing ways to stabilize and inhibit them reversibly. Finally, the search for next-generation novel detergent proteases shall be based on new backbones different from subtilisins.
REFERENCES
- Almog, O., T. Gallagher, M. Tordova, J. Hoskins, P. Bryan and G.L. Gilliland, 1998. Crystal structure of calcium-independent subtilisin BPN0 with restored thermal stability folded without the prodomain. Proteins, 31: 21-32.
PubMed - Becker, T., G. Park and A.L. Gaertner, 1997. Formulating of detergent enzymes. Surfactant Sci. Ser., 69: 299-325.
Direct Link - Bhosale, S.H., M.B. Rao, V.V. Deshpande and M.C. Srinivasan, 1995. Thermostability of high activity alkaline protease from Conidiobolus coronatus (NCL 86.8.20). Enzyme Microb. Technol., 17: 136-139.
CrossRef - Bott, R., 1997. Development of new proteases for detergents. Surfactant Sci. Ser., 69: 75-91.
Direct Link - Bryan, P.N., 2000. Protein engineering of subtilisin. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol., 1543: 203-222.
CrossRefDirect Link - Cherry, J.R. and A.L. Fidantsef, 2003. Directed evolution of industrial enzymes: An update. Curr. Opin. Biotechnol., 14: 438-443.
PubMed - Choi, I.G., W.G. Bang, S.H. Kim and Y.G. Yu, 1999. Extremely thermostable serine type protease from Aquifex pyrophilus, molecular cloning, expression and characterization. J. Biol. Chem., 274: 881-888.
Direct Link - Coolbear, T., C.W. Eames, Y. Casey, R.M. Daniel and H.W. Morgan, 1991. Screening of strains identified as extreamely thermophilic bacilli for extracellular proteolytic activity and general properties of the proteinases from two of the strains. J. Applied Bacteriol., 71: 252-264.
CrossRef - Cowan, D., R.M. Daniel and H. Morgan, 1985. Thermophilic proteases: Properties and potential applications. Trends Biotechnol., 3: 68-72.
CrossRef - Dhandapani, R. and R. Vijayaragavan, 1994. Production of a thermophilic, extracellularalkaline protease by Bacillus sterothermophilus AP-4. World J. Microbiol. Biotechnol., 1: 33-35.
CrossRef - Estell, D.A., T.P. Graycar and J.A. Wells, 1985. Engineering an enzyme by site-directed mutagenesis to be resistant to chemical oxidation. J. Biol. Chem., 260: 6518-6521.
PubMed - Fujii, M., M. Takagi, T. Imanaka and S. Aiba, 1983. Molecular cloning of a thermostable neutral protease gene from Bacillus stearothermophilus in a vector plasmid and its expression in Bacillus stearothermophilus and Bacillus subtilis. J. Bacteriol., 154: 831-837.
PubMed - Garcia-Carreno, F.L., 1992. Protease inhibition in theory and practice. Biotechnol. Educ., 3: 145-150.
Direct Link - Geesecke, U.E., G. Bierbaum, H. Rudde, U. Spohn and C. Wandrey, 1991. Production of alkaline protease with Bacillus licheniformis in a controlled fed-batch process. Applied Microbiol. Biotechnol., 35: 720-724.
CrossRef - Graycar, T., M. Knapp, G. Ganahaw, J. Dauberman and R. Bott, 1999. Engineered Bacillus lentus subtilisins having altered flexibility. J. Mol. Biol., 10: 97-109.
PubMed - Gunz, P.J., M.D. Bauer, Y. Sun, A.M. Fieno and R.A. Grant et al., 2004. Stabilized variant of streptomyces subtilisin inhibitor and its use in stabilizing subtilisin BPN'. Protein Eng. Des. Selection, 17: 333-339.
Direct Link - Gupta, R., Q. Beg and P. Lorenz, 2002. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol., 59: 15-32.
CrossRefPubMedDirect Link - Jaswal, R.K. and G.S. Kocher, 2007. Partial characterization of a crude alkaline protease from Bacillus circulans and its detergent compatibility. Internet J. Microbiol., 4: 1-5.
Direct Link - Kobayashi, T., Y. Hakamada, S. Adachi, J. Hitomi and T. Yoshimatsue et al., 1995. Purification and properties of an alkaline protease from alkalophilic Bacillus sp. KSM-K16. Applied Microbial. Biotechnol., 43: 473-481.
CrossRef - Kumar, D. and T.C. Bhalla, 2004. Bacillus sp. APR-4 protease as a laundry additive. Indian J. Biotechnol., 3: 563-567.
Direct Link - Kumar, D. and T.C. Bhalla, 2004. Purification and characterization of a small size protease from Bacillus sp. APR-4. Indian J. Exp. Biol., 42: 515-521.
PubMedDirect Link - Kumar, D. and T.C. Bhalla, 2005. Microbial proteases in peptide synthesis: Approaches and applications. Applied Microbiol. Biotechnol., 68: 726-736.
CrossRefPubMedDirect Link - Kumar, D., D. Chand, S. Uma Dutt and T.C. Bhalla, 2003. Utilization of a Bacillus sp. APR4: Protease in peptide synthesis. Indian J. Microbiol., 43: 131-133.
Direct Link - Legendre, D., N. Laraki, T. Graslund, M.E. Bjornvad and M. Bouchet et al., 2000. Display of active subtilisin 309 on phage: Analysis of parameters influencing the selection of subtilisin variants with changed substrate specificity from libraries using phosphonylating inhibitors. J. Mol. Biol., 296: 87-102.
CrossRef - Morihara, K., 1974. Comparative specificity of microbial proteinases. Adv. Enzymol., 41: 179-243.
CrossRefDirect Link - Mulder, F.A., D. Schipper, R. Bott and R. Boelens, 1999. Altered flexibility in the substrate-binding site of related native and engineered high-alkaline Bacillus subtilisins. J. Mol. Biol., 10: 111-123.
PubMed - Najafi, M.F., D. Deobagkar and D. Deobagkar, 2005. Potential application of protease isolated from Pseudomonas aeruginosa PD100. Electronic J. Biotechnol., 8: 197-203.
Direct Link - Ness, J.E., M. Welch, L. Giver, M. Bueno and J.R. Cherry et al., 1999. DNA shuffling of subgenomic sequences of subtilisin. Nature Biotechnol., 17: 893-896.
PubMed - Ness, J.E., S. Kim, A. Gottman, R. Pak and A. Krebber et al., 2002. Synthetic shuffling expands functional protein diversity by allowing amino acids to recombine independently. Nature Biotechnol., 20: 1251-1255.
CrossRefDirect Link - Phadatare, S.U., V.V. Deshpande and M.C. Srinivasan, 1993. High activity alkaline protease from Conidiobolus coronatus (NCL 86.8.20): Enzyme production and compatibility with commercial detergents. Enzyme Microb. Technol., 15: 72-76.
CrossRefDirect Link - Prakash, M., R.M. Banik and C. Koch-Brandt, 2005. Purification and characterization of Bacillus cereus protease suitable for detergent industry. Applied Biochem. Biotechnol., 127: 143-155.
CrossRef - Rachna and V.K. Tyagi, 2006. Gels: Novel detergents for laundry applications. J. Oleo Sci., 55: 555-562.
Direct Link - Rao, M.B., A.M. Tanksale, M.S. Ghatge and V.V. Deshpande, 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev., 62: 597-635.
Direct Link - Russell, G.L. and L.N. Britton, 2002. Use of certain alcohol ethoxylates to maintain protease stability in the presence of anionic surfactants. J. Surfact. Detergents, 5: 5-10.
CrossRefDirect Link - Saeki, K., J. Hitomi, M. Okuda, Y. Hatada and, T. Kobayashi et al., 2000. Novel oxidatively stable subtilisin-like serine proteases from alkaliphilic Bacillus spp.: Enzymatic properties, sequences and evolutionary relationships. Biochem. Biophys. Res. Commun., 279: 313-319.
CrossRef - Saeki, K., K. Ozaki, T. Kobayashi and S. Ito, 2007. Detergent alkaline proteases: enzymatic properties, genes and crystal structures. J. Biosci. Bioeng., 103: 501-508.
PubMed - Saeki, K., J. Hitomi, M. Okuda, Y. Hatada and Y. Kageyama et al., 2002. A novel species of alkaliphilic Bacillus that produces an oxidatively stable alkaline serine protease. Extremophiles, 6: 65-72.
CrossRefPubMedDirect Link - Samal, B., B. Karan, T.C. Boone, K.K. Chen, M.F. Rahde and Y. Stabinsky, 1989. Cloning and expression of the gene encoding a novel proteinase from Tritirachium album. Limber Genet., 85: 329-333.
PubMed - Saravani, G.A., D.A. Cowan, R.M. Daniel and H.W. Morgan, 1989. Caldolase, a chelator insensitive extracellular serine proteinase from Thermus sp. Biochem. J., 262: 409-416.
PubMed - Takagi, H. and M. Takahashi, 2003. A new approach for alteration of protease functions: Pro-sequence engineering. Applied Microbiol. Biotechnol., 63: 1-9.
CrossRef - Takimura, Y., K. Saito, M. Okuda, Y. Kageyama and K. Saeki et al., 2007. Alkaliphilic Bacillus sp. Strain KSM-LD1 contains a record number of subtilisin-like serine proteases genes. Applied Microbiol. Biotechnol., 76: 395-405.
PubMed - Tindbaek, N., A. Svendsen, P.R. Oestergaard and H. Draborg, 2004. Engineering a substrate-specific cold-adapted subtilisin. Protein Eng. Des. Selection, 17: 149-156.
PubMed - Tomaschova, J., W. Buchinger, W. Hampel and J. Zemanovic, 1998. Purification and characterization of extracellular proteinase produced by Brevibacterium linens ATCC9172. Food Chem., 4: 499-503.
CrossRef - Tsuchiya, K., Y. Nakamura, H. Sakashita and T. Kimura, 1992. Purification and characterization of a thermostable alkaline protease from alkalophilic Thermoactinomyces sp. HS682. Biosci. Biotech. Biochem., 56: 246-250.
PubMed - Wintrode, P.L., K. Miyazaki and F.H. Arnold, 2000. Cold adaptation of a mesophilic subtilisin-like protease by laboratory evolution. J. Biol. Chem., 275: 31635-31640.
CrossRefDirect Link - Aftab, S., S. Ahmed, S. Saeed and S.A. Rasool, 2006. Screening, isolation and characterization of alkaline protease producing bacteria from soil. Pak. J. Biol. Sci., 9: 2122-2126.
CrossRefDirect Link - Chu, W.H., 2007. Optimization of extracellular alkaline protease production from species of Bacillus. J. Ind. Microbiol. Biotechnol., 34: 241-245.
CrossRef - Dodia, M.S., R.H. Joshi, R.K. Patel and S.P. Singh, 2006. Characterization and stability of extracellular alkaline protease from halophilic and alkaliphilic bacteria isolated from saline habitat of coastal Gujarat, India. Braz. J. Microbiol., 37: 276-282.
CrossRef - Ibrahim, A.S.S., N.M.A. EI-Shayeb and S.S. Mabrouk, 2007. Isolation and identification of alkaline protease producing bacteria from an Egyptian soda lake. J. Applied Sci. Res., 3: 1363-1368.
Direct Link - Patel, R., D. Mittal and S.P. Singh, 2005. Extracellular alkaline protease from a newly isolated haloalkaliphilic Bacillus sp.: Production and optimization. Process Biochem., 40: 3569-3575.
CrossRef - Kirk, O., T.V. Borchert and C.C. Fuglsang, 2002. Industrial enzyme applications. Curr. Opin. Biotechnol., 13: 345-351.
CrossRefPubMedDirect Link