
Intira Thampayak
Department of Biotechnology, Graduated School, Chiang Mai University, Chiang Mai 50200, Thailand
Naowarat Cheeptham
Department of Biological Sciences, Faculty of Science, Thompson Rivers University, 900 McGill Road, Box 3010, Kamloops, BC, V2C 5N3, Canada
Wasu Pathom-Aree
Department of Biology, Faculty of Science, Chiang Mai University, Chiang Mai 50200, Thailand
Pimporn Leelapornpisid
Faculty of Pharmacy, Chiang Mai University, Chiang Mai 50200, Thailand
Saisamorn Lumyong
Department of Biotechnology, Graduated School, Chiang Mai University, Chiang Mai 50200, Thailand
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 7 | Page No.: 499-507







ABSTRACT
Two hundred and twenty-nine soil actinomycete strains were initially screened for extracellular biosurfactant activity by a drop-collapse method in Kim`s medium containing sesame oil as a sole source of carbon. Three isolates, namely S71, S72 and S177, were capable of biosurfactant production. Phenotypic and genotypic analysis strongly suggested that they were members of the genus Streptomyces. The isolates S71 and S177 were closely related to S. griseoflavus sharing 99% 16S rRNA gene similarities, whereas S72 was closely related to S. fradiae sharing only 98% 16S rRNA gene similarities suggesting that this may represent a novel species. The cell-free culture broth of the three isolates had emulsification activity and decreased surface tension. According to emulsification activity (E24) and surface tension values observed in the three isolates, Streptomyces sp. S72 was selected for biosurfactant production in larger scale. The cell-free culture broth of the isolate S72 was further extracted with chloroform:methanol (2:1) and two fractions were found positive in producing biosurfactants. To determine structure and molecular weight of the two positive fractions, the Nuclear Magnetic Resonance (NMR) spectroscopy and Mass Spectrometry (MS) will be carried out.
PDF Abstract XML References Citation
How to cite this article
Intira Thampayak, Naowarat Cheeptham, Wasu Pathom-Aree, Pimporn Leelapornpisid and Saisamorn Lumyong, 2008. Isolation and Identification of Biosurfactant Producing Actinomycetes From Soil. Research Journal of Microbiology, 3: 499-507.
URL: https://scialert.net/abstract/?doi=jm.2008.499.507
URL: https://scialert.net/abstract/?doi=jm.2008.499.507
INTRODUCTION
Biosurfactants are biological surface-active compounds produced by microorganisms such as bacteria, yeasts and fungi from various sources of substrates including sugars, oils, alkanes and wastes (Lin, 1996). These compounds are amphipathic molecules containing hydrophilic and hydrophobic domains. Biosurfactants reduce surface and interfacial tensions in both aqueous solutions and hydrocarbon mixtures, which make them potential candidates in enhancing oil recovery and in the emulsification process (Lin, 1996; Maier, 2003). Compared to chemical surfactants, biosurfactants hold significant advantages including lower toxicity, higher structural diversity and concomitant diversity in properties. In addition, biosurfactants may be produced from renewable resources with a higher biodegradability and better environmental compatibility.
In terms of production, it has been observed that biosurfactants are commonly produced in the stationary phase of microbial growth, for instance, in Bacillus subtilis MTCC 2423 (Makkar and Cameotra, 1997) and Arthrobacter paraffineus ATCC 19558 (Duvnlak et al., 1982). The environmental factors and growth conditions of the culture such as pH, temperature, agitation and oxygen availability have also been shown to affect biosurfactant production through their effects on cellular growth or activity (Desai and Banat, 1977). In addition, biosurfactant production using water-soluble carbon sources was inferior to that obtained with water-immiscible substrates such as n-alkanes and olive oil (Robert et al., 1989). However, biosurfactant production has been proven to be quite a challenge in industry. Problems such as low yield and high production cost of biosurfactants (Lang and Wullbrandt, 1999) are important and cannot be ignored. It is hoped that these problems can be solved by searching for more effective biosurfactants and/or higher yield of biosurfactant production from microorganisms. Therefore, when it comes to the production of biosurfactants for industrial application, actinomycetes, gram-positive filamentous bacteria, are of good choice as biosurfactant producers because of their abundance in soil and their major roles in recycling of material in nature. Moreover, they have been found to produce many kinds of metabolites, including antibiotics, pigments, enzymes and biosurfactants (Oskay et al., 2004; Augustine et al., 2005; Imasda, 2005; Richter et al., 1998). Until recently, there have been only a limited number of reports published on biosurfactant-producing actinomycetes. For instance, Streptomyces tendae Tu 901/8c produces an extracellular hydrophobic peptide compound, named streptofactin (Richter et al., 1998). This example highlights the potential of actinomycetes as biosurfactant producers.
The objective of this study was to isolate soil actinomycetes in Chiang Mai and to screen for extracellular biosurfactants using sesame oil that is easily available in the area as a sole carbon source.
MATERIALS AND METHODS
Selective Isolation of Actinomycetes
Soil samples were collected from various locations in Chiang Mai province, Northern Thailand in 2005. The samples were air dried at room temperature for 1 week and pretreated at 55 °C in a hot-air oven for 3 h then stored at room temperature in plastic bags. One gram of air-dried soil was serially diluted and 100 μL aliquots of the appropriate dilution was applied to humic acid-salts-vitamin agar (HV) plates (Hayakawa and Nonomura, 1987) which the pH was adjusted to 7.0. These plates were supplemented with 50 μg mL-1 of nalidixic and nystatin and incubated at 30 ± 2 °C for up to 3 weeks. Actinomycete colonies were preliminarily selected based on colony morphology and transferred on to Hickey-Tresner agar (HT) plates (Hickey and Tresner, 1952). Pure isolates were maintained on HT agar medium at 4 °C. Alternatively, suspension of mycelia and spores were stored in 15% glycerol at -20 °C for long term preservation.
Screening for Biosurfactant-Producing Isolates
Seven day-old actinomycetes were inoculated into 6x100 mm test tubes containing 3 mL Kim`s medium (Kim et al., 2000) with 3% sesame oil as carbon source. The broth cultures were incubated at 30 ± 2oC on a reciprocal shaker at 120 rpm for 5 days. The culture broth was then tested for the production of extracellular biosurfactants with the drop-collapse method (Bodour and Miller-Maier, 1998). The drop-collapse method was performed in a polystyrene lid of a 96-microwell plate coated with 1.8 μL of 10W-40 Pennzoil® oil, which was spread as a thin coating over the bottom of the well. The coated wells were allowed to equilibrate for 24 h at room temperature. A 5 μL of sample was added into the center of the well and after 1 min, the results were recorded. If the droplet remained intact, the result was scored as negative. If the droplet collapsed, the result was scored as positive meaning biosurfactant production occurred. Distilled water and 10% Sodium Dodecyl Sulfate (SDS) were used as negative and positive controls respectively and all the tests were carried out in triplicate.
Analytical Techniques
Biosurfactant Activity
The oil-displacement method was used to detect the activity of biosurfactant. Forty milliliters of distilled water was added to a Petri dish followed by the addition of 10 μL of crude oil to the surface of the water. Ten microliters of sample was added onto the center of the oil film. The diameters of the clear zone on the oil surface were measured and compared with control using uninoculated medium (Youssef et al., 2004).
Emulsification Activity
The emulsification capacity was determined by adding 2 mL of kerosene to the same amount of cell-free culture broth, mixed for 2 min on a vortex mixer and allowed to stand for 24 h. E24 index is defined as percentage of the height of emulsified layer divided by the total height of the liquid column (Cooper and Goldenberg, 1987).
Surface Tension Measurement
Surface tension measurement was done by using Kruss tensiometer, Model K10 (Kruss, Germany) equipped with plate. Samples were cell-free culture broths. Measurements of surface tension from distilled water were used as negative controls.
Identification of Isolates
Phenotypic Characterization
Phenotypic characterization of the selected isolates was based on cell morphology (Pridham et al., 1958) and classical biochemical test (Williams et al., 1983). After the preliminary screening, positive strains were characterized using traditional microbiological methods such as growth on microbiological media, aerial spore mass color, substrate mycelium pigmentation and color of any soluble pigments. They were recorded using the method described by Kelly (1958); these properties were noted from mature, heavily sporing cultures. The spore chain morphology was examined with light and scanning electron microscopy (JEAL, JSM-5910, Japan) of 14-day-old cultures grown on HT plates at 28 °C. Diaminopimelic acid isomers and sugar from whole-cell extract were analyzed for chemotaxonomic studies by thin layer chromatography of whole-cell hydrolysates (Becker et al., 1964; Boone and Pine, 1968).
Biochemical Tests
The biochemical tests used in this study were degradation of casein, tyrosine, gelatin, starch and aesculin, carbohydrate utilization and lipase activity (Williams et al., 1983).
16S rRNA Gene Amplification, Sequencing and Phylogenetic Analysis
Cell pellets were obtained from three day-old culture of S71, S72 and S177. Total genomic DNA of each isolate was extracted with the salting out procedure (Kieser et al., 2000). PCR amplification of 16S rRNA gene sequence was done in GeneAmp® (PCR System 9700, AB Applied Biosystem) using primer 27f (5`-AGA,GTT,TGA,TCM,TGG,CTC,AG-3`) and primer 1525r (5`-AAG,GAG,GTG,WTC,CAR,CC-3`). The PCR amplifications were done using an initial denaturation step at 95 °C for 5 min, followed by 35 cycles of 1 min at 95 °C, 1 min at 58 °C and 2 min at 72 °C and a final extension at 72 °C for 10 min and cooled to 4 °C. Bidirectional sequencing of the PCR product was done with four primers. These were primer 27f, primer 1525r, primer MG3f (5`-CTA CGG GRS GCA GCA G -3`) and primer MG5f (AAA CTC AAA GGA ATT GAC GG -3`). The cycle sequencing was performed by TECH DRAGON LIMITED, Hong Kong.
The probable identity or nearest match of each new sequence was determined by performing a Basic Local Alignment Search Tool (BLAST) search at the NCBI (National Center for Biotechnology Information, Building 38A, 8600 Rockville Pike, Bethesda, MD 20894, USA) website (Wheeler et al., 2001). The almost complete 16S rDNA sequences of each of the test strains were aligned manually using the PHYDIT program against corresponding sequences of representatives of appropriate actinomycete reference strains retrieved from the GenBank (Benson et al., 2004) and the RDPII databases (Ribosomal Database Project; Maidak et al., 2001). The aligned sequences were used to generate phylogenetic trees and similarity matrices using the PHYDIT software. Unrooted phylogenetic trees were inferred by using the least-squares (Fitch and Margoliash, 1967), maximum-parsimony (Kluge and Farris, 1969) and neighbour-joining (Saitou and Nei, 1987) tree-making algorithms. An evolutionary distance matrix was generated after Jukes and Cantor (1969). The topologies of the resultant trees were evaluated by bootstrap analyses of the neighbor-joining method based on 1000 resamplings. The phylogenetic analyses, which were carried out using the TREECON program (Van de Peer and deWachter, 1994), were presented as rooted dendrograms using the TREECON and TREEVIEW programs.
Nucleotide Sequence Accession Number
The nucleotide sequences of S71, S72 and S177 have been deposited with GenBank under accession numbers EF392566, EF208617 and EF197893, respectively.
Extraction of Biosurfactant
The biosurfactant was extracted from culture after cell removal by filtration. A mixture of chloroform:methanol (2:1 v/v) was added to the culture medium, after being vigorously shaken, this was allowed to stand until phase separation. The extracts were combined and concentrated by rotary evaporator (Buchi Rotavapor R-200, Switzerland) and then sodium sulfate anhydrous was added to remove water. The crude extract was obtained after removal of the solvent and moisture by evaporation and vacuum drying (Dura-DryTM, FTSYSTEMS, USA), respectively. The crude extract was purified by column chromatography on a silica gel (Scharlau GE 0030) (100% hexane -100% EtOAc as eluent). Each fraction was collected and concentrated and then subjected to TLC analysis. All fractions were tested for biosurfactant activity using the oil displacement method.
RESULTS AND DISCUSSION
Isolation of and Screening for Biosurfactant Producing Actinomycetes
A total of 229 strains of actinomycetes were isolated from a total of 25 soil samples. All isolates were initially screened for extracellular biosurfactant by a drop-collapse method for their ability to produce biosurfactant(s) in culture broth containing sesame oil as the sole carbon source. This method is a simple protocol for screening biosurfactant production in a large number of microorganisms (Youssef et al., 2004). Significant drop collapsed activity were observed from three isolates, designated strains S71, S72 and S177. Subsequently, the extracellular biosurfactant production tested positively when grown in assigned broth medium (pH 7) at 30 ±2 °C on a reciprocal shaker set at 120 rpm after 5 days of incubation. Biosurfactant productions of these isolates were confirmed with the oil displacement method (Youssef et al., 2004).
Characterization of Selected Strains
Morphology and chemotaxonomy of the positive isolates were studied and the results indicated that these three isolates belonged to the genus Streptomyces. All three isolates had morphological features and biochemical characteristics that supported their assignment to the genus Streptomyces (Table 1, Fig. 1). Isolates S71 and S72 had identical morphological features, whereas isolate S177
| Table 1: | Morphological and biochemical characteristics of the three biosurfactant-producing actinomycetes (S71, S72 and S177) |
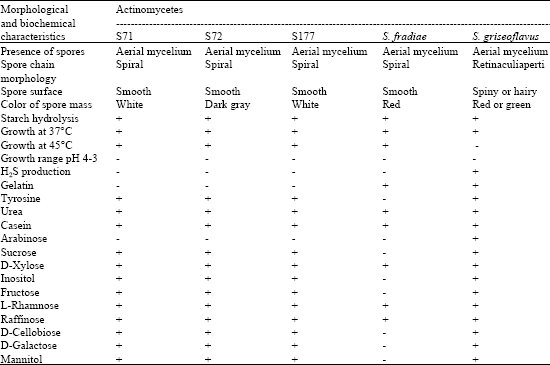 | |
| +: Positive, utilized, -: Negative, Results of reference strain search from Williams et al. (1983) | |
 | |
| Fig. 1: | lectron micrographs of biosurfactant-producing actinomycetes; Streptomyces sp. S71 (A), S72 (B) and S177 (C) |
differed from isolates S71 and S72 in colonial pigment. All three isolates were highly-branched substrate mycelia with production of aerial hyphae with straight or spiral spore chains. Results from chemotaxonomic study showed that all three isolates had LL isomer of diaminopimelic acid (LL-DAP) in their cell wall. These results corresponded with the work done and reported by Kieser et al. (2000). Therefore, isolates S71, S72 and S177 were then assigned to the genus Streptomyces.
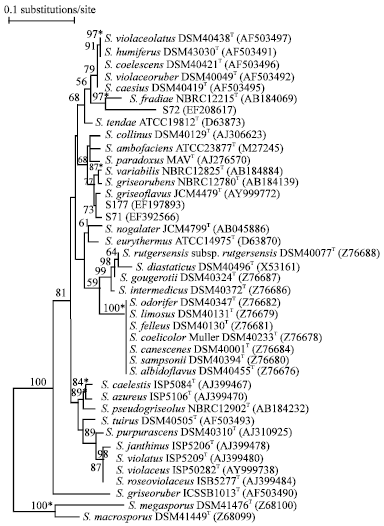 | |
| Fig. 2: | Neighbor-joining tree (Saitou and Nei, 1987) based on almost complete 16S rRNA gene sequences showing relationships between the isolates S71, S72 and S177 and representatives of the genus Streptomyces. The asterisks indicate the branches that were also recovered using the least-squares (Fitch and Margoliash 1967) and maximum-parsimony (Kluge and Farris 1969) tree-making algorithms. The numbers at the nodes indicate the level of bootstrap support (%) based on a neighbor joining analysis of 1000 resampled datasets; only value above 50% are given. The scale bar indicates 0.1 substitutions per nucleotide position |
16S rRNA Gene Sequence Analysis
Further taxonomic characterization of biosurfactant-producing actinomycetes was done by 16S rRNA gene sequence analysis. It is evident from 16S rRNA gene phylogenetic trees that isolates S71, S72 and S177 are members of the genus Streptomyces (Fig. 2). These data were in line with the results obtained from chemotaxonomic studies. Isolates S71 and S177 formed phyletic line that was closely related to S. griseoflavus JCM4479T sharing 16S rRNA gene similarities with the latter of 99.7 and 99.8%, values equivalent to 4 and 2 nt differences at 1402 sites. This taxonomic relationship was supported by a 73% bootstrap value. Similar tree topology was also found in the tree generated with the least-squares (Fitch and Margoliash, 1967) algorithms. It is obvious that these two isolates are closely related to S. griseoflavus.
The isolate S72 was recovered near the S. violaceoruber clade, a taxon underpinned by a 97% bootstrap value. Similar tree topology was also found in the trees generated with the least-squares (Fitch and Margoliash, 1967) and maximum parsimony (Kluge and Farris, 1969) algorithms (data not shown).
| Table 2: | Tests of biosurfactant production in cell-free culture broth by the drop collapse method, oil-displacement method, emulsifying activity and surface tension measurement |
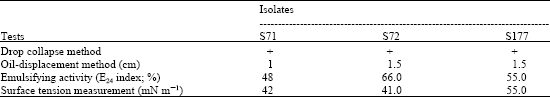 | |
| +: Positive, drop collapsed When larger numbers are observed in the oil displacement and the emulsifying activity, the better the production, When smaller numbers are observed in the surface tension measurement, the results are favorable | |
The isolate showed its closest relationship with the type strain of S. fradiae NBRC12215T, the two organisms shared a 16S rRNA gene similarity of only 98.1%, a value equivalent to 27 nt differences at 1453 sites. These data suggested that the isolate S72 may represent a novel species within the genus Streptomyces.
Extraction of Biosurfactant
The emulsification activity (E24) of cell-free culture broth of the S71, S72 and S177 isolates was 48, 66 and 55%, respectively and the Surface Tension (ST) values of the cell-free culture medium were 42, 41 and 45 mN m-1, respectively (Table 2). These results indicated that actinomycetes remain a vital source of microbial metabolites production. According to the E24 and ST values aforementioned, isolate S72 was further studied for the biosurfactant production in larger scale. After the extraction of the biosurfactant produced by isolate S72, the crude extract was purified by column chromatography and thin layer chromatography. There are two fractions that were positive for biosurfactant activity after tested with the oil displacement method. Further analysis by Nuclear Magnetic Resonance spectroscopy (NMR) and Mass Spectrometry (MS) to determine the structure and molecular weight of the positive fractions will be carried out.
There are very few reports published on actinomycetes that produce extracellular biosuractant(s). This study showed that three Streptomyces isolates, S71, S72 and S177, which were isolated from soils were able to produce extracellular biosurfactant(s) in the modified Kim`s broth medium with sesame seed oil as a sole carbon source. They could grow in Kim`s medium supplemented with crude oil as carbon source . The results from 16S rRNA gene sequence analysis indicated that S71 and S177 were closely related to S. griseoflavus and S72 was closely related to S. fradiae. Richter et al. (1998) reported that S. tendae produced extracellular biosurfactant called streptofactin which was used by the cells to develop aerial mycelium and support the erection of aerial hyphae by lowering the surface tension of water films enclosing the colonies. Additionally, S. griseoflavus, S. parvus and S. plicatus isolated from Kuwait Burgan oil field were found to utilize n-hexadecane, n-octadecane, kerosene and crude oil as carbon and energy sources (Barabas et al., 2001). These results indicated that actinomycetes are still important source of microbial metabolites production. It was found that five different actinomycetes potentially used as bioremediation agents in polycyclic aromatic hydrocarbons degradation produced biosurfactants when grown in various substrates (Pizzul et al., 2006). Gordonia strains (M22, BS25 and BS29) produced two classes of surface-active compounds when using aliphatic hydrocarbons as carbon and energy sources (Franzetti et al., 2008). However, to obtain more diverse potential and novel actinomycetes, it is recommended that a wider rang of soil samples be used for biosurfactant activity.
CONCLUSION
Isolates S71, S72 and S177 produced extracellular biosurfactant which reduced surface tension. They were isolated from soil samples in Chiang Mai, Thailand. These isolates are then characterized by using combined analyses of 16S rRNA gene sequence, morphological and biochemical data and identified to the genus Streptomyces. Phylogenetic analysis using 16S rRNA gene sequence showed that S71 and S177 were closely related to S. griseoflavus (AY999772) sharing 99% 16S rRNA gene similarities, whereas S72 was closely related to the S. fradiae that shared a 16S rRNA gene similarity of only 98% and might be novel species. Further study using type strain for comparison and the structure elucidation of purified biosurfactant will be conducted.
ACKNOWLEDGMENTS
This research was financially supported by the National Research Council of Thailand (NRCT), the Faculty of Science, Chiang Mai University, Chiang Mai Thailand and Rangsit University, Pathumthani, Thailand.
REFERENCES
- Augustine, S.K., S.P. Bhavsar and B.P. Kapadnis, 2005. A non-polyene antifungal antibiotic from Streptomyces albidoflavus PU 23. J. Biosci., 30: 201-211.
Direct Link - Barabas, G., G. Vargha, I.M. Szabo, A. Penyige and S. Damjanovich et al., 2001. n-alkane uptake and utilization by Streptomyces strains. Antonie van Leeuwenhoek, 79: 269-276.
Direct Link - Becker, B., M.P. Lechevalier, R.E. Gordon and H.A. Lechevalier, 1964. Rapid differentiation between Nocardia and Streptomyces by paper chromatography of whole-cell hydrolysates. Applied Microbiol., 12: 421-423.
PubMed - Benson, D.A., I. Karsch-Mizrachi, D.J. Lipman, J. Ostell and D.L. Wheeler, 2004. GenBank: Update. Nucleic Acids Res., 32: D23-26.
Direct Link - Bodour, A.A. and R.M. Miller-Maier, 1998. Application of a modified drop-collapse technique for surfactant quantitation and screening of biosurfactant-producing microorganisms. J. Microbiol. Methods, 32: 273-280.
CrossRefDirect Link - Boone, C.J. and L. Pine, 1968. Rapid method for characterization of actinomycetes by cell wall composition. Applied Microbiol., 16: 279-284.
Direct Link - Cooper, D.G. and B.G. Goldenberg, 1987. Surface-active agents from two Bacillus species. Applied Environ. Microbiol., 53: 224-229.
Direct Link - Desai, J.D. and I.M. Banat, 1997. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev., 61: 47-64.
PubMedDirect Link - Duvnlak, Z., D.G. Cooper and N. Kosaric, 1982. Production of surfactant by Arthrobacter paraffineus ATCC 19558. Biotechnol. Bioeng., 24: 165-175.
CrossRef - Fitch, W.M. and E. Margoliash, 2001. Construction of phylogenetic trees, a method based on mutation distances as estimated from cytochrome c sequences is of general applicability. Science, 155: 279-284.
Direct Link - Franzetti, A., G. Bestetti, P. Caredda, P. La Colla and E. Tamburini, 2008. Surface-active compounds and their role in the access to hydrocarbons in Gordonia strains. FEMS Microbiol. Ecol., 63: 238-248.
Direct Link - Hayakawa, M. and H. Nonomura, 1987. Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J. Ferment. Technol., 65: 501-509.
CrossRefDirect Link - Hickey, R.T. and H.D. Tresner, 1952. A cobalt-containing medium for sporulation of Streptomyces species. J. Bacteriol., 64: 891-892.
PubMed - Imada, C., 2005. Enzyme inhibitors and other bioactive compounds from marine actinomycetes. Antonie Van Leewenhoek, 87: 59-63.
CrossRefDirect Link - Kelly, K.L., 1958. Centroid notations for the revised ISCC-NBS color-name blocks. J. Res. Nat. Bureau Standards USA., 61: 427-427.
Direct Link - Kim, S.H., E.J. Lim, S.O. Lee, J.D. Lee and T.H. Lee, 2000. Purification and characterization of biosurfactants from Nocardia sp. L-417. Biotechnol. Applied Biochem., 31: 249-253.
PubMedDirect Link - Kluge, A.G. and F.G. Farris, 1969. Quantitative phyletics and the evolution of anurans. Syst. Zool., 18: 1-32.
Direct Link - Maidak, B.L., J.L. Cole, T.G. Lilbum, C.T. Parker Jr. and P.R. Saxman et al., 2001. The RDP-II (Ribosomal Database Project). Nucl. Acids Res., 29: 173-174.
CrossRefDirect Link - Maier, R.M., 2003. Biosurfactants: Evolution and diversity in bacteria. Adv. Applied Microbiol., 52: 102-121.
Direct Link - Makkar, R.S. and S.S. Cameotra, 1997. Utilization of molasses for biosurfactant production by two Bacillus strains at thermophilic conditions. J. Am. Oil. Chem. Soc., 74: 887-889.
CrossRefDirect Link - Oskay, A.M., T. Usame and A. Cem, 2004. Antibacterial activity of some actinomycetes isolated from farming soils of Turkey. Afr. J. Biotechnol., 3: 441-446.
Direct Link - Pizzul, L., M.P. Castillo and J. Stenstrom, 2006. Characterization of selected actinomycetes degrading polyaromatic hydrocarbons in liquid culture and spiked soil. World J. Microbiol. Biotechnol., 22: 745-752.
CrossRefDirect Link - Pridham, T.G., C.W. Hesseltine and R.G. Benedict, 1958. A guide for the classification of streptomycetes according to selected groups; placement of strains in morphological sections. Applied Microbiol., 6: 52-79.
PubMedDirect Link - Richter, M., J.M. Willey, R. Suβmuth, G. Jung and H.P. Fiedler, 1998. Streptofactin, a novel biosurfactant with aerial mycelium inducing activity from Streptomyces tendae Tu 901/8c. FEMS Microbiol. Lett., 163: 165-171.
CrossRefDirect Link - Robert, M., M.E. Mercade, M.P. Bosch, J.L. Parra, M.J. Espuny, M.A. Manresa and J. Guinea, 1989. Effect of the carbon source on biosurfactant production by Pseudomonas aeruginosa 44T. Biotechnol. Lett., 11: 871-874.
CrossRef - Saitou, N. and M. Nei, 1987. The neighbour-joining method: A new method for constructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
PubMedDirect Link - Van de Peer, Y. and R. de Wachter, 1994. TREECON for Windows: A software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput. Applied Biol. Sci., 10: 569-570.
CrossRefDirect Link - Wheeler, D.L., D.M. Church, A.E. Lash, D.D. Leipe and T.L. Madden et al., 2001. Database resources of the national center for biotechnology information. Nucleic Acids Res., 29: 11-16.
Direct Link - Williams, S.T., M. Goodfellow, G. Alderson, E.M.H. Wellington, P.H.A. Sneath and M.J. Sackin, 1983. Numerical classification of Streptomyces and related genera. J. Gen. Microbiol., 129: 1743-1813.
CrossRefPubMedDirect Link - Youssef, N.H., K.E. Duncan, D.P. Nagle, K.N. Savage, R.M. Knapp and M.J. McInerney, 2004. Comparison of methods to detect biosurfactant production by diverse microorganisms. J. Microbiol. Methods, 56: 339-347.
CrossRefDirect Link